
Abstract
Proteins induced in rice by auxin and zinc were determined by proteome analysis. Cultured suspension cells of rice were treated with 2,4-dichlorophenoxyacetic acid and ZnSO4 and then proteins were separated by two-dimensional polyacrylamide gel electrophoresis; seven proteins were found to be induced by auxin and zinc. Of these seven, methylmalonate-semialdehyde dehydrogenase (MMSDH) was elevated by treatment with auxin alone. MMSDH was detected in cultured suspension cells, root and leaf sheath, but not in leaf blades. MMSDH responded to auxin and gibberellin, but did not respond to brassinolide and cytokinin. Furthermore, the amount of MMSDH in slr1, a constitutive gibberellin response mutant, was 2-fold that of wild type. MMSDH mRNA and protein were stimulated in root formation induced by auxin and/or zinc over a 4-week period. These results suggest that MMSDH may be necessary for root formation in rice induced by auxin and/or zinc.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Auxin plays a critical role in regulating diverse responses both at the whole-plant level, such as apical dominance, lateral root initiation and emergence (Casimiro et al. 2001), and at the cellular level, such as cell elongation, division, and differentiation (Abel and Theologis 1996). In addition, exogenous auxin induces rapid increases in steady-state transcript levels of a small set of genes (Hagen and Guilfoyle 2002), alters ion flux at the plasma membrane, and stimulates changes in the cell-wall (Becker and Hedrich 2002). At relatively low concentrations of indole-3-acetic acid (IAA), tobacco BY-2 cells elongate without dividing, whereas at higher concentrations, they divide with regenerative expansion (Chen 2001). These phenomena in cells are regulated through signal transduction that probably embodies a complex network rather than a linear pathway. However, how auxin affects plant growth and development is not well understood.
In plants, the cotyledons and younger leaves have the highest biosynthetic capacity for auxin. However, auxin levels in these tissues are markedly decreased in a zinc-deficient environment (Cakmak et al. 1989). Therefore, in most studies, functional aspects of zinc nutrition in plants have been investigated in correlative studies coupled with auxin (Skoog 1940). Zinc is an essential mineral nutrient and a cofactor of over 300 enzymes and proteins involved in cell division, nucleic acid metabolism, and protein synthesis (Marschner 1986). Previous research has established the involvement of zinc in the biosynthesis of IAA from tryptophan, since an increase in tryptophan content was detected in zinc-deficient plants (Tsui 1948). However, an increase in the level of tryptophan due to zinc deficiency is not only specific to tryptophan, just as a lower IAA level is not always caused by the impairment of biosynthesis of IAA from tryptophan. A normal level of IAA was found in zinc-deficient radishes (Raphanus sativus cv. Akamaru-Hatsukadaikon) by means of thin layer chromatography, gas-liquid chromatography, and gas chromatography-mass spectrometry (Hossain et al. 1998). These reports suggest the possibility that zinc not only participates in auxin biosynthesis, but also plays multiple roles in plant development induced by auxin. On the other hand, a significantly low level of gibberellin (GA) in zinc-deficient plants of Zea mays L. showing dwarfism similar to the biosynthesis mutant of GA has been reported (Sekimoto et al. 1997; Suge et al. 1986). Thus, zinc is speculated to play important roles in the biosynthesis and metabolism of GA and auxin. Although zinc has been widely studied in plants, with much research into the effects of zinc deficiency, very little is known about the effect of high zinc levels together with phytohormones.
Recently, proteomics has become an essential methodology for large-scale analysis of proteins in various fields of plant biology (Pandey and Mann 2000). Proteomics, in combination with the availability of genome sequence data, has opened up enormous possibilities for identifying the total set of expressed proteins, as well as their expression levels (Rossignol 2001). In addition, proteome analysis using differential display with two-dimensional polyacrylamide gel electrophoresis (2D-PAGE) has several advantages over other approaches commonly used for similar studies, e.g., in efficient separation of complex protein mixtures, analysis of isoforms or secondary modifications of proteins such as glycosylation and phosphorylation of macromolecular complexes such as protein-ligand, protein-protein, protein-DNA interactions, and in proteolysis using low amounts of proteins (Natera et al. 2000).
Some of formation of callus and root in rice is regulated by both auxin and zinc (Saeki et al. 2000); however, the mechanisms of these phenomena remain unclear. In this study, proteins induced by auxin and zinc were analyzed by proteome analysis. The proteins of rice cultured suspension cells induced by 2,4-dichlorophenoxyacetic (2,4-D) and zinc were analyzed using 2D-PAGE, Edman sequencing and matrix-assisted laser desorption-ionization time-of-flight mass spectrometry (MALDI-TOF MS).
Materials and methods
Plant materials
Rice (Oryza sativa L. cv. Nipponbare) was used in this study. Mature seeds were husked, sterilized with 70% ethanol for 5 min and 1% NaClO for 30 min. The seeds were washed with sterilized water and allowed to germinate on an agar plate of MS medium supplemented with 9.0 μM 2,4-D (Wako, Osaka, Japan) and incubated at 25°C in darkness. Cells were allowed to proliferate for about 1 month, and were then subcultured in N6 standard medium supplemented with 4.5 μM 2,4-D for suspension culture. The medium was changed once every 2 weeks during subculturing. A constitutive GA response mutant, slenderrice 1(slr1), was isolated as described in Ikeda et al. (2001). Cultured suspension cells of both wild type and slr1 were subcultured and dedifferentiated for about 1 year, and then used in this study. For hormonal studies with 2,4-D, GA, brassinolide (BL), 6-benzylaminopurine (BA) and/or zinc (ZnSO4) (Wako), rice cultured suspension cells were placed in 2,4-D-free medium for 2 h and incubated for 2 days in the appropriate medium. To examine tissue specificity, rice plants were grown in vessels containing commercial soils blended with chemical fertilizers and cultivated in an air-controlled greenhouse under natural light. To observe the effects of auxin and zinc on root formation, rice seeds were dehusked before being surface sterilized by stirring in 70% ethanol for 2 min and then in 5% NaCLO for 30 min, followed by washes in sterile water. The sterilized seeds were placed on N6 medium, containing 0.45 μM 2,4-D, 30 g l−1 sucrose, 4 g l−1 agar and 520 μM ZnSO4. Cultures were kept at about 30°C.
Protein extraction and 2D-PAGE
A portion (200 mg) of cultured suspension cells was homogenized in 400 μl lysis buffer (O’Farrell 1975) containing 8 M urea, 2% Nonidet P-40, 2% ampholine (pH 3.5–10 and 5.0–8.0), 5% (w/v) polyvinylpyrrolidone. The homogenate was centrifuged at 15,000 g for 10 min and the supernatant (50 μl) was subjected to 2D-PAGE. Isoelectric focusing (IEF) and immobilized pH gradient (IPG) (Daiichi Kagaku, Tokyo, Japan) were carried out in a glass tube for the first dimension, and SDS-PAGE for the second dimension was performed in a 15% separation gel and detected by Coomassie Brilliant Blue (CBB) staining. CBB-stained gels were scanned using a flatbed scanner, and the positions of individual proteins were analyzed and evaluated automatically using Image Master 2D Elite software (Amersham Biosciences, Buckinghamshire, UK). The isoelectric point and molecular mass of each protein were calibrated using 2D-PAGE standards (Bio-Rad, Richmond, Calif.).
Amino acid sequence analysis
To determine the N-terminal amino acid sequence of the protein, proteins separated by 2D-PAGE were electroblotted onto polyvinylidene difluoride (PVDF) membranes (Fluorotrans; Pall, Port Washington, N.Y.) using a semi-dry transfer blotter (Nippon Eido, Tokyo, Japan), and visualized by CBB staining. For internal amino acid sequencing, protein spots were removed from 2D-PAGE gels stained with CBB, and were then electro-eluted from the gel pieces using an electrophoretic concentrator (ISCO, Lincoln, Calif.) at 200 V for 2 h. Each protein solution was dialyzed for 2 days against deionized water. Staphylococcus aureus V8 protease (Pierce, Rockford, Ill.) (10 μl; 0.1 μg μl−1) in deionized water and 10 μl SDS sample buffer (pH 6.8) were overlaid with 20 μl protein dissolved in 20 μl SDS sample buffer (pH 6.8) (Laemmli 1970). Electrophoresis was performed until the sample and protease were stacked in the stacking gel, and the power supply was interrupted for 30 min for protein digestion (Cleveland et al. 1977). Electrophoresis was then continued, and the separated digests were electroblotted onto a PVDF membrane.
The proteins were excised from the PVDF membrane and analyzed with a gas-phase protein sequencer (Model 494; PE Applied Biosystems, Foster City, Calif.). The amino acid sequences obtained were compared with protein sequences in the SwissProt database using the FASTA sequence alignment program (Pearson and Lipman 1998).
MALDI-TOF MS
CBB-stained protein spots were excised from gels, washed with 25% (v/v) methanol and 7% (v/v) acetic acid for 12 h at room temperature, and then destained with 50 mM NH4HCO3 in 50% (v/v) methanol for 1 h at 40°C. Proteins were reduced with 10 mM DTT in 100 mM NH4HCO3 for 1 h at 60°C and then incubated with 40 mM iodoacetamide in 100 mM NH4HCO3 for 30 min at room temperature. The gel pieces were minced and allowed to dry, and then rehydrated in 100 mM NH4HCO3 with 1 pmol trypsin at 37°C overnight. The digested peptides were extracted from the gel slices with 0.1% trifluoroacetic acid (TFA) in 50% (v/v) acetonitrile/water three times. The peptide solution, thus obtained was dried, reconstituted with 30 μM 0.1% TFA in 5% acetonitrile/water, and then desalted with ZipTip C18 pipette tips (Millipore, Bedford, Mass.). MALDI-TOF MS was performed using a Voyager Elite XL time-of-flight mass spectrometer (Applied Biosystems, Framingham, Mass.). The peptide solution was mixed with the matrix solution—the supernatant of a 50% acetonitrile solution saturated with α-cyano-4-hydroxycinnamic acid—and then air dried on the flat surface of a stainless steel plate. Calibrations were carried out using a standard peptide mixture. The mass spectra were subjected to a sequence database search with Mascot software (Matrix Science, London, UK).
Preparation of DNA probe
The methylmalonate-semialdehyde dehydrogenase (MMSDH) gene-specific probe was amplified from an expressed sequence tag (EST) clone of rice MMSDH subcloned in pBluescript SK + vector. The EST clone was obtained from the DNA bank of National Institute of Agrobiological Sciences (Japan). The full-length insert of MMSDH was amplified by PCR using T7 (5′-AATACGACTCACTATAG-3′) and T3 (5′-ATTAACCCTCACTAAAG-3′) primers. PCR reactions were performed as follows: 94°C for 3 min, then 30 cycles of 94°C for 1 min, 60°C for 1 min, 72°C for 2 min, followed by an extension step at 72°C for 10 min. The PCR products were purified using a QIAquick PCR purification kit (Qiagen, Hilden, Germany), and then labeled with [α-32P]dCTP (Amersham) using a random prime labeling system (Rediprime II, Amersham), and used as probe in Northern blot analysis.
RNA isolation and Northern blot analysis
Total RNA was isolated from rice cultured suspension cells, roots, leaf sheaths, and leaf blades by phenol/chloroform extraction as described by Chomczynski and Sacchi (1987). For Northern analysis total RNA (20 μg) was separated by electrophoresis in a 1.2% agarose-formaldehyde gel and blotted onto a Hybond-N membrane (Amersham). Blots were hybridized with the 32P-labeled full-length insert of the MMSDH EST clone using hybridization buffer (UltraHyb; Ambion, Austin, Tex.). Blots were washed twice, first in 2×SSC and 0.1% SDS at 65°C for 15 min, then in 0.1×SSC and 0.1% SDS at 65°C for 20 min, then analyzed with a phosphor image program on a Typhoon 8610 variable imager (Amersham).
Protein extraction and immunoblotting
Proteins were extracted from 200 mg rice cultured suspension cells by grinding in 400 μl SDS sample buffer (Laemmli 1970). For rice plants, proteins were extracted from 1,000 mg rice roots by grading in 1,000 μl SDS sample buffer. The lysates were centrifuged at 15,000 g at 4°C for 10 min. The supernatant was precipitated with 50% trichloroacetic acid (final concentration 10%) on ice for 30 min, centrifuged at 15,000 g for 10 min, and the pellet suspended in 100 μl SDS sample buffer. Aliquots of 5 μl were subjected to SDS-PAGE. For immunoblot analysis, protein samples were separated on a 15% SDS-PAGE, and transferred onto PVDF membrane. Anti-MMSDH antibody was obtained against the following sequence: STAAAASWLSFSASS. Anti-rabbit IgG horseradish peroxidase-linked antibodies (Amersham) were used as secondary antibodies. Binding of antibodies was detected using an ECL Plus Western blotting detection kit (Amersham).
Results and discussion
Seven proteins are induced by treatment with auxin and zinc in rice cultured suspension cells
Auxin and zinc are involved in callus and root formation in rice plants (Saeki et al. 2000). To investigate proteins regulated by auxin and zinc, 2D-PAGE was performed using cultured suspension cells. Cultured suspension cells require auxin for multiplication. Cultured suspension cells of rice were treated with various levels of ZnSO4 in the presence of 2,4-D. The concentration of 2,4-D in the culture medium was kept stable at 4.5 μM, which is the amount usually added to the culture medium in order to maintain multiplication of cultured suspension cells of rice. When incubated with 260 μM ZnSO4, which is 50 times more than the zinc content in N6 standard medium, seven proteins were induced (Fig. 1A). The proteins all declined at higher zinc concentrations (520 and 1,040 μM ZnSO4, Fig. 1B). The proteins determined here as auxin-induced and zinc-induced were named CS326, CS531, CS658, CS670, CS743, CS783, and CS822.

Comparison of two-dimensional polyacrylamide gel electrophoresis (2D-PAGE) patterns of proteins in rice cultured suspension cells treated with auxin and zinc. The auxin concentration in the culture medium was kept constant [4.5 μM 2,4-dichlorophenoxyacetic acid (2,4-D)]. ZnSO4 concentrations (μM): a 5.2 (control); b 260; c 520; d 1,040. A Proteins were extracted from 2-day-old cultured suspension cells, separated by 2D-PAGE and detected by Coomassie Brilliant Blue (CBB) staining. Isoelectric focusing (IEF) for the first dimension is from right to left, and SDS-PAGE for the second dimension is from top to bottom. Arrows Positions of auxin-induced and zinc-induced proteins. B Protein spots were quantified densitometrically relative to the control (a). Each value represents the mean of three replicates
The seven proteins were identified by electroblotting onto a PVDF membrane and determining the N-terminal sequence by Edman degradation in a gas-phase sequencer. N-Terminal sequences were obtained for six proteins, with only one protein (CS822) being N-terminally blocked (Table 1). In addition, internal amino acid sequencing was performed, identifying these proteins as MMSDH (CS326), NADPH-dependent oxidoreductase (NADPH-DO, CS658), elongation factor (CS531), elongation factor 1-β′ (CS670), nascent polypeptide associated complex α chain (CS743), l-ascorbate peroxidase (APO, CS783) and late embryogenesis abundant protein (LEA, CS822).
The amino acid sequence tags of CS743 using the N-terminal domain showed complete alignment to the nascent polypeptide-associated complex α chain from O. sativa. The sequence of this protein is given as distributed in class V of rice chromosome 1 (Sasaki et al. 2002). Class V matches an EST nucleotide sequence, though the proteins remain unknown. LEA proteins (CS822) accumulate during periods of water deficiency, functioning to maintain ion and water homeostasis. A tomato LEA protein confers salt- and freezing-tolerance when expressed in Saccharomyces cerevisiae (Imai et al. 1996), whereas the wheat LEA protein Em functions as an osmoprotective molecule in yeast (Swire-Clark and Marcotte 1999). In protoplasts isolated from the laminar pulvinus of Phaseolus vulgaris, auxin enhanced the cellular content of osmotic solutes (Iino et al. 2001). Romo et al. (2001) suggested that the cellular function of CapLEA-1 and CapLEA-2, belonging to group 3 LEAs, is to prevent high ion concentration damage. The induction of LEA in this experiment may be a response to the high zinc conditions.
Treatment with auxin and zinc induced two elongation factors (CS531 and CS670) in rice cultured suspension cells. Translation elongation factor-1 (EF-1) is composed of four subunits: α, β, β′ and γ. EF-1α plays a major role in protein synthesis by catalyzing the binding of aminoacyl tRNA to the acceptor site on the ribosome, and the genes encoding it are constitutively expressed in plant cells (Curie et al. 1993; Morelli et al. 1994). EF-1β and EF-1β′ stimulate the exchange of GDP bound to EF-1α with GTP. EF-1α is highly expressed in tissues such as meristems, somatic embryos, and auxin-treated tissues that are undergoing high levels of protein synthesis and cell division (Morelli et al. 1994). Roots are a particularly attractive system for studying auxin action because of their morphological simplicity and well-characterized auxin responses. Indirect evidence from several other studies suggests that auxin has a central role in cell division and elongation in the growing root. In the case of callus, a high level of 2,4-D is required for both induction and maintenance. The EF-1α promoter directed strong gusA expression primarily in root tips and regenerating callus, rather than in the leaves, of Gladiolus, consistent with its expression in actively dividing cells (Kamo and Blowers 1999). This study raises the possibility that auxin confers a regulating effect in active sites such as root and callus, even though the direct effector may be biosynthesis of the elongation factor.
Kato and Esaka (1999) proposed that APO (CS783) functions as a scavenger of the H2O2 produced by basal metabolism that accompanies both cell division and cell elongation. The growth responses induced by auxin appear to be generally correlated with a degradation of cell-wall polysaccharides, and auxin promoted the production of superoxide radicals (O2 −), a hydroxyl radical (•OH) precursor (Schopfer 2001). This •OH could act as a site-specific oxidant, targeted to play a useful physiological role in the cell-wall loosening processes underlying cell expansion (Fry 1998). Moreover, •OH can be generated from O2 − and H2O2 by peroxidases bound to the cell-wall matrix and, in this way, •OH is directed to sites of polymer cleavage induced cell-wall loosening (Schopfer 2001). This may account for the preferential induction of APO upon treatment with auxin and zinc (Fig. 1 and Table 1), and also for the positive effect of the latter two compounds on cell growth. The induction of such unique proteins suggests that, when present together with zinc, auxin brings about various activities in cell growth and development that may as yet be unknown, or which have been unconsidered to date.
MMSDH is markedly induced by auxin alone
Callus induction can be induced on practically all plant explants. Differences in the rate of cell conversion towards dedifferentiation are conditioned by the chemical structure of the exogenous growth regulators used at the ontogenetic stage of explants, and the concentration gradient between exogenous and endogenous growth regulators (Hlinkva and Ruzickova 2000). This shows that the development of the whole plant, not just the individual cell, is conditioned not only by the genetic background of the plant cell but also by the concentration gradient of exogenous growth regulators. We examined how the auxin-induced and zinc-induced proteins behaved in response to varying the auxin level. Two proteins (MMSDH and NADPH-DO) were induced by 2,4-D, especially MMSDH, which was markedly elevated with increasing 2,4-D concentration (Fig. 2), whereas there was no noticeable change in the levels of the other five proteins in response to 2,4-D.

Comparison of 2D-PAGE pattern of proteins in rice cultured suspension cells treated with increasing concentration of auxin (2,4-D; μM): a 0, b 0.45, c 4.5, d 22.6. A Proteins were extracted from cultured suspension cells incubated for 2 days, separated by 2D-PAGE and detected by CBB staining. IEF for the first dimension is from right to left and SDS-PAGE for the second dimension is from top to bottom. Arrows Positions of auxin-induced proteins. B Protein spot of MMSDH was quantified densitometrically relative to the control (a). Each value represents the mean of three replicates
NADPH-DOs include NADPH oxidase, nitric oxide synthase and other flavoproteins. These enzymes are essential for various cell functions involving NADPH, and are capable of reducing carbonyl groups on a wide variety of compounds, including sugars, aldehyde metabolites and steroid hormones. NADPH oxidases have been found in human fetal membranes and plasma membranes of plants, and play important roles in defense of the fetus against bacterial infiltration from the maternal side (Matsubara et al. 2001), and in the regulation of defense strategies upon infection with pathogens or stimulation by elicitors (Mehdy et al. 1996), respectively. Some redox enzymes, including NADPH-oxidoreductase, are capable of generating O2 −, which plays many roles in plants (Van Gestelen et al. 1997). Schopfer (2001) demonstrated that •OH, originating from O2 − and H2O2 in the cell-wall, might be an essential component of the biochemical mechanism engaged in wall loosening during auxin-induced extension growth. Our findings, together with these other reports, indicate that auxin induces NADPH-DO, with subsequent cell-wall loosening resulting in cell growth.
MMSDH has been isolated from rat and human liver, and is also found in other tissues such as kidney, heart, and muscle (Kedishvili et al. 1992). Recently, a number of pistil-abundant cDNA clones that are related to flowering events in rice were isolated using simplified differential display; these clones included some predictive to encode MMSDH (Yoshida and Kuboyama 1997). MMSDH catalyzes the irreversible oxidative decarboxylation of malonate-semialdehydes and methylmalonate-semialdehydes to acetyl-CoA and propionyl-CoA, respectively, in the distal portions of the valine and pyrimidine catabolic pathways. Using H2O2 as a model stress, Sweetlove et al. (2002) demonstrated that MMSDH decreased dramatically under oxidative stress. Oxidative stress imposes significant flux restrictions on the TCA cycle and electron transport, thereby limiting the synthesis of ATP. In this study, MMSDH, representing a range of metabolic activities, was increased upon treatment with auxin. This indicates that auxin-mediated plant growth is a result of accelerated cell metabolism due to MMSDH induced by auxin.
The high MMSDH expression levels induced upon addition of auxin are maintained over time
To characterize auxin-induced proteins in more detail, a time-course of changes in levels of MMSDH and NADPH-DO was examined, comparing untreated cultures with those treated with 2,4-D every day for 6 days (Fig. 3). In the absence of 2,4-D, the level of neither protein changed appreciably during the experimental period. Upon addition of 4.5 μM 2,4-D, the level of MMSDH was elevated, and remained high with the passage of time. As found in these results, a constant level of MMSDH was maintained, although there were differences in the amount accumulated according to culture conditions.

Changes in levels of two auxin-induced proteins over time (circles MMSDH, diamonds NADPH-DO). Cultured suspension cells were treated without (open circles, open diamonds) or with 4.5 μM 2,4-D (filled circles, filled diamonds) for 6 days. The graph depicts relative protein amounts over time
To further confirm the identification of MMSDH in more detail, internal amino acid sequences were determined using the Cleveland peptide mapping method (Cleveland et al. 1977) and MALDI-TOF MS. Briefly, internal amino acid sequencing was performed on a gas-phase protein sequencer. Based on the sequence tags obtained, a homology search was carried out in the DNA data bank of Japan (DDBJ) with FASTA. Protein spots resolved by 2D-PAGE were then excised from the gel and digested with trypsin while still in the gel slice. The resultant tryptic peptides were measured by MALDI-TOF MS. Based on the set of observed molecular masses, a sequence database search using Mascot was carried out for protein identification. The auxin-induced protein CS326 matched the reported amino acid sequence of a rice protein, i.e., MMSDH from O. sativa (Fig. 4).

Amino acid sequence comparison of MMSDH (accession number AF045770) and CS326. Amino acids are numbered on the left. Boxed amino acids N-Terminal amino acid sequence determined using protein sequencer, double underlined amino acids internal amino acid sequence determined using mass spectrometry
MMSDH is unique among the known aldehyde dehydrogenase (ALDHs) because it is CoA-dependent and capable of conserving energy in the form of a CoA ester during the oxidation of an aldehyde. The sequence (SAG) XFXXXGQXCX (AGN) present in ALDHs is considered to be the consensus active site motif (Kedishvili et al. 1992). Perfect conservation of this consensus motif occurs in the MMSDH identified here, between amino acid residues 308 and 319 (AXFXXXGQXCXA) (Fig. 4). Like other ALDHs, MMSDH has esterase activity, hydrolyzing p-nitrophenyl acetate with concomitant formation of an S-acyl enzyme and, in the presence of CoA, generating acetyl-CoA (Kedishvili et al. 1992). Decarboxylation of methylmalonate-semialdehyde is an important feature of the reaction. Acetyl-CoA is an important metabolic intermediate involved in the glyoxylate cycle, TCA cycle, and so on, and is also involved in generating fatty acids. Paul et al. (1998) have reported that rapid activation of a phospholipase A2, a typical signal transduction reaction, was shown by auxin in cultured suspension parsley (Petrosilenum crispum L.) and soybean (Glycine max L.) cells, and furthermore that fatty acid(s) could be second messengers in several auxin functions, especially in cell elongation. Other workers have also indicated that phospholipase A activity generates fatty acids as metabolites; fatty acids and their derivatives are recognized more and more as signaling intermediates (Kirsch et al. 1997). The results presented here suggest that MMSDH is enhanced by application of auxin, because it seems that MMSDH is involved in generating fatty acids, which is necessary for auxin functions.
Tissue specificity and hormonal effects
To determine where MMSDH is expressed in rice, we performed Northern and Western blot analyses using cultured suspension cells, roots, leaf sheaths, and leaf blades (Fig. 5A). In Northern blot analysis, the MMSDH probe hybridized to a 1.6 kb message. Differences in mRNA abundance between these tissues was observed, with the greatest accumulation of mRNA being found in cultured suspension cells. mRNA also accumulated in roots and leaf sheaths, but only slightly in leaf blades. A similar result was seen in Western blot analysis, with highest protein accumulation in cultured suspension cells. The protein expression level in roots was slightly higher than in leaf blades, but was scarcely detectable in leaf sheaths, where mRNA also showed negligible levels. Stem elongation and root development are regulated by GA and auxin, respectively (Reid et al. 1983; Laskowski et al. 1995). The accumulation of MMSDH mRNA in these tissues may be the result of localization to tissues that are targets of auxin-induced and GA-induced growth.

Expression of MMSDH mRNA and protein in different tissues. Total RNA and protein were extracted from cultured suspension cells incubated for 2 days, or from different tissues of 2-week-old seedlings. A Northern blot of total RNA hybridized to an MMSDH probe. Ethidium bromide-stained rRNA bands are shown to verify equal loading of the total RNA. B Upper panel CBB-stained proteins are shown to verify equal loading of the protein, lower panel immunodetection using an anti-MMSDH antibody on a Western blot of protein separated by SDS-PAGE. Lanes: 1 Cultured suspension cells, 2 root, 3 leaf sheath, 4 leaf blade
Auxin, GA, cytokinin and brassinosteroid particularly promote fundamental processes such as cell elongation and division (Kende and Zeevaart 1997). The induction of MMSDH could result either from an involvement in cell development or as a response to excess hormones induced by increasing auxin levels in the culture medium. To distinguish between these possibilities, we examined the response of MMSDH to GA, BL, and BA as well as to auxin. As shown in Fig. 6, MMSDH increased when auxin and GA were added to the culture medium during incubation of cultured suspension cells for 2 days. However, BL and BA did not influence expression of MMSDH, indicating that expression of MMSDH is triggered by developmental processes regulated by auxin and GA.

Changes in MMSDH levels in rice cultured suspension cells treated with various hormones. Rice cultured suspension cells were treated with A 2,4-D, B gibberellic acid (GA), C brassinolide (BL), and D 6-benzylaminopurine (BA) in varying concentrations as indicated. Circles Positions of MMSDH spot. Histograms Relative amounts of MMSDH under the various hormone treatments
It is generally accepted that these phytohormones are associated with cell expansion in whole plants or excised organs as described above. Lee et al. (2000), working with mesophyll suspension cultures of Zinnia, reported that cytokinin had a greater effect on radial cell expansion, while auxin and BL promoted only cell elongation, and GA had nearly equal effects on expansion in both directions. The selective response of MMSDH to various hormones indicates its involvement in certain specific effects of auxin and GA activity in plants, e.g., in the direction of cell expansion caused by phytohormones.
MMSDH in slr1, a constitutive GA response mutant
The above results show that MMSDH is implicated in GA activity as well as that of auxin (Fig. 7). To investigate whether MMSDH is involved in GA signal transduction or not, we used slr1, a line with a single recessive mutation resulting in a constitutive GA response phenotype. The mutant contains a loss-of function mutation in the SLR1 gene, which is a negative regulator of the GA signaling process, and consequently elongates as if saturated with GA (Ikeda et al. 2001). In slr1, there was an enhanced level of MMSDH compared with the wild type (Fig. 7A); the relative amount was approximately 2-fold (Fig. 7B). Similarly, the longitudinal length of epidermal cells of leaf sheaths in slr1 is approximately 2-fold that of wild type (Ikeda et al. 2001). This finding supports the suggestion above, i.e., that the MMSDH response, at least regarding cell growth, can be attributed to GA activation.

2D-PAGE pattern of proteins in cultured suspension cells from wild type rice and the GA response mutant of rice, slr1. A Proteins were extracted from wild type (a) and slr1 (b) cultured suspension cells, separated by 2D-PAGE and detected by CBB staining. Arrowhead Position of MMSDH. B Relative amounts of MMSDH in cultured suspension cells separated by 2D-PAGE. a Wild type, b slr1
Effect of auxin and zinc on root formation and the expression of MMSDH
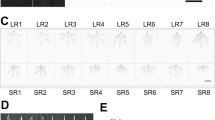
Numerous earlier studies have shown that IAA, the major endogenous auxin in plant, is necessary for both the initiation and the later development of lateral roots and adventitious roots. Four-week-old rice seedlings were grown on N6 medium in vitro, and treated without phytohormones (as a control) and with 0.45 μM 2,4-D and 0.45 μM 2,4-D/520 μM zinc; the differences in treatment were apparently reflected in root formation (Fig. 8A). In this experiment, 0.45 μM 2,4-D supplied to the culture medium stimulated rooting compared with the control, and induced emergence of lateral and adventitious roots (Fig. 8A). Root morphology in the control, with no added phytohormones, generally showed root elongation (Fig. 8A). The addition of a high level of zinc (520 μM) to the culture medium with 0.45 μM 2,4-D, further enhanced lateral and adventitious root formation (Fig. 8A). Standard N6 medium contains 5.2 μM zinc, whereas our results show that the optimal concentration (520 μM) is 100-fold higher, and this without showing any inhibition due to excess zinc. Similar results were observed by Hossain et al. (1997) and Saeki et al. (2000), who demonstrated that a high level of zinc stimulates callus formation, one of the physiological functions of auxin. Taken together, these findings indicate that zinc at high concentration is an important factor in auxin action, especially in callus and root formation.

Effects of auxin and zinc on root formation and MMSDH expression. A Rice seedlings were grown in N6 medium treated without 2,4-D as control (1), or with 0.45 μM 2,4-D (2) or 0.45 μM 2,4-D/520 μM zinc (3). B Northern blot of total RNA hybridized to the MMSDH probe. Ethidium bromide-stained rRNA bands are shown to verify equal loading of total RNA. C CBB-stained gel (upper panel) and Western blot reacted with anti-MMSDH antibody (lower panel) of proteins separated by SDS-PAGE. Lanes in B and C: 1 no treatment, 2 0.45 μM 2,4-D, 3 0.45 μM 2,4-D/520 μM zinc
MMSDH mRNA and protein expression were enhanced, and increased rooting was observed, through addition of auxin alone or auxin+zinc (Fig. 8B,C). Addition of 0.45 μM 2,4-D (without or with zinc) dramatically increased the amount of MMSDH mRNA compared with the control that had no added phytohormones in the culture medium. This indicates that MMSDH is an auxin-regulated gene and might be required for auxin action in rice root formation. This result also suggests that MMSDH is related to cell development, corroborating the results observed in slr1 (Fig. 7).
Concluding remarks
The relationship of zinc to auxin has been well documented and considered in auxin synthesis. Although at present it has become possible to study auxin effects within the framework of what we know about signal transduction in general, the relationship of auxin and zinc, even in auxin synthesis, is not yet clear. We demonstrated that high levels of zinc stimulated root formation in the presence of auxin (Fig. 8A). The results shown here lead to the possibility that zinc plays an important role with respect to auxin-regulated plant growth and development. This concept is given support by the induction of various proteins involved in cell growth in response to zinc and auxin (Fig. 1 and Table 1). MMSDH was identified as a phytohormone-regulated protein in this study, particularly in response to auxin, and MMSDH was also found to be elevated in slr1 (Fig. 7). In addition, MMSDH mRNA and protein expression were observed in cultured suspension cells, roots, and leaf sheaths (Fig. 5). These results suggest that MMSDH plays a role in the developmental stages of organs and cells, and also indicate that MMSDH may be involved in an interactive effect with auxin and GA, or in individual responses between MMSDH and auxin or MMSDH and GA on plant growth. Further study is needed to clarify how MMSDH is involved in response to plant growth as induced by phytohormones with the aim of understanding the mechanisms of the physiological function of auxin.
The findings that seven proteins were induced by auxin and zinc might have important implications for understanding the physiological effect of auxin on plant development. In this study, we characterized MMSDH as one of these proteins. MMSDH mRNA levels in roots increased concomitant with auxin-induced and/or zinc-induced root formation. The results presented here are the first indication that MMSDH is related to the physiological functions of phytohormones, especially auxin and GA, and may contribute to understanding of the molecular basis of rice response to auxin action, and serve as a basis for additional studies. Further examination and characterization of other auxin-induced and zinc-induced proteins may reveal at least one of the development pathways involved in auxin action.
Abbreviations
- BA :
-
6-Benzylaminopurine
- BL :
-
Brassinolide
- CBB :
-
Coomassie Brilliant Blue
- 2,4-D :
-
2,4-Dichlorophenoxyacetic acid
- GA :
-
Gibberellic acid
- IAA :
-
Indole-3-acetic acid
- MALDI-TOF MS :
-
Matrix-assisted laser desorption-ionization time-of-flight mass spectrometry
- MMSDH :
-
Methylmalonate-semialdehyde dehydrogenase
- 2D-PAGE :
-
Two-dimensional polyacrylamide gel electrophoresis
- PVDF :
-
Polyvinylidene difluoride
References
Abel S, Theologis A (1996) Early genes and auxin action. Plant Physiol 111:9–17
Becker D, Hedrich R (2002) Channelling auxin action: modulation of ion transport by indole-3-acetic acid. Plant Mol Biol 49:349–356
Cakmak I, Marschner H, Bangerth F (1989) Effect of zinc nutritional status on growth, protein metabolism and leaves of indole-3-acetic acid and other phytohormones in bean (Phaseolus vulgaris L.). J Exp Bot 40:405–412
Casimiro I, Marchant A, Bhalerao RP, Beeckman T, Dhooge S, Swarup R, Graham N, Inzé D, Sandberg G, Casero PJ, Bennett M (2001) Auxin transport promotes Arabidopsis lateral root initiation. Plant Cell 13:843–852
Chen JG (2001) Dual auxin signaling pathways control cell elongation and division. J Plant Growth Regul 20:255–264
Chomczynski P, Sacchi N (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162:156–159
Cleveland DW, Fischer SG, Kirschner MW, Laemmli UK (1977) Peptide mapping by limited proteolysis in sodium dodecyl sulfate and analysis by gel. J Biol Chem 252:1102–1106
Curie C, Axelos M, Bardet C, Atanassova R, Chaubet N, Lescure B (1993) Modular organization and developmental activity of an Arabidopsis thaliana EF-1α gene promoter. Mol Gen Genet 238:428–436
Fry SC (1998) Oxidative scission of plant cell wall polysaccharides by ascorbate-induced hydroxyl radicals. Biochem J 332:507–515
Hagen G, Guilfoyle T (2002) Auxin-responsive gene expression: genes, promoters and regulatory factors. Plant Mol Biol 49:373–385
Hlinkva E, Ruzickova C (2000) Gene expression of soybean calli culture affected by auxinoids with various chemical structure. Soybean Genet Newslett 27
Hossain B, Hirata N, Nagatomo Y, Akashi R, Takaki H (1997) Internal zinc accumulation is correlated with increased growth in rice suspension culture. J Plant Growth Regul 16:239–243
Hossain B, Hirata N, Nagatomo Y, Suiko M, Takaki H (1998) Zinc nutrition and level of endogenous indole-3-acetic acid in radish shoots. J Plant Nutr 21:1113–1128
Iino M, Long C, Wang X (2001) Auxin- and abscisic acid-dependent osmoregulation in protoplasts of Phaseolus vulgaris pulvini. Plant Cell Physiol 42:1219–1227
Ikeda A, Ueguchi-Tanaka M, Sonoda Y, Kitano H, Koshioka M, Futsuhara Y, Matsuoka M, Yamaguchi J (2001) Slender rice, a constitutive gibberellin response mutant, is caused by null mutation of the SLR1 gene, an ortholog of the height-regulating gene GAI/RGA/RHT/D8. Plant Cell 13:999–1010
Imai R, Chang L, Ohta A, Bray EA, Takagi M (1996) A lea-class gene of tomato confers salt and freezing tolerance when expressed in Saccharomyces cerevisiae. Gene 170:243–248
Kamo K, Blowers A (1999) Tissue specificity and expression levels of gusA under rolD, mannopine synthase and translation elongation factor 1 subunit β promoters in transgenic Gladiolus plants. Plant Cell Rep 18:809–815
Kato N, Esaka M (1999) Changes in ascorbate oxidase gene expression and ascorbate levels in cell division and cell elongation in tobacco cells. Physiol Plant 105:321–329
Kedishvili NY, Popov KM, Rougraff PM, Zhao Y, Crabb DW, Harris RA (1992) CoA-dependent methylmalonate-semialdehyde dehydrogenase, a unique member of the aldehyde dehydrogenase superfamily. J Biol Chem 267:19724–19729
Kende H, Zeevaart JAD (1997) The five “classical” plant hormones. Plant Cell 9:1197–1210
Kirsch C, Takamiya-Wik M, Reinold S, Hahlbrock K, Somssich IE (1997) Rapid, transient, and highly localized induction of plastidial omega-3-fatty acid desaturase mRNA at fungal infection sites in Petrosilenum crispum. Proc Natl Acad Sci USA 94:2079–2084
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680–685
Laskowski MJ, Williams ME, Nusbaum HC, Sussex IM (1995) Formation of lateral root meristems is a two-stage process. Development 121:3303–3310
Lee S, Woffenden BJ, Beers EP, Roberts AW (2000) Expansion of cultured Zinnia mesophyll cells in response to hormones and light. Physiol Plant 108:216–222
Marschner H (1986) Mineral nutrition of higher plants. Academic, New York
Matsubara S, Takizawa T, Takayama T, Iwasaki R, Minakami H, Sato I (2001) Cytochemically detectable glucose-6-phosphate dehydrogenase is present in human fetal membrane chorion leave trophoblasts. Placenta 22:613–616
Mehdy MC, Sharma YK, Sathasivan K, Bays NW (1996) The role of activated oxygen species in plant disease resistance. Physiol Plant 98:365–374
Morelli JK, Shewmaker CK, Vayda ME (1994) Biphasic stimulation of translational activity correlates with induction of translation elongation factor 1 subunit β upon wounding in potato tubers. Plant Physiol 106:897–903
Natera SH, Guerreiro N, Djordjevic MA (2000) Proteome analysis of differentially displayed proteins as a tool for the investigation of symbiosis. Mol Plant Microbe Interact 13:995–1009
O’Farrell PH (1975) High resolution two-dimensional electrophoresis of proteins. J Biol Chem 250:4007–4021
Pandey A, Mann M (2000) Proteomics to study genes and genomes. Nature 405:837–846
Paul RU, Holk A, Scherer GFE (1998) Fatty acids and lysophospholipids as potential second messengers in auxin action. Rapid activation of phospholipase A2 activity by auxin in suspension-cultured parsley and soybean cells. Plant J 16:601–611
Pearson WR, Lipman DJ (1998) Improved tools for biological sequence comparison. Proc Natl Acad Sci USA 85:2444–2448
Reid JB, Murfet IC, Potts WC (1983) Internode length in Pisum. II. Additional information on the relationship and action of loci Le, La, Cry, Na and Lm. J Exp Bot 34:349–364
Romo S, Labrador E, Dopico B (2001) Water stress-regulated gene expression in Cicer arietinum seedling and plants. Plant Physiol Biochem 39:1017–1026
Rossignol M (2001) Analysis of the plant proteome. Curr Opin Biotechnol 12:131–134
Saeki Y, Yasukouchi A, Nagatomo Y, Takaki H (2000) Distinctive expression of a zinc-binding protein in rice callus growth with high zinc concentration. Soil Sci Nutr 46:209–216
Sasaki T, Matsumoto T, Yamamoto K, Sakata K, Baba T, Katayose Y, Wu J, Niimura Y, Cheng Z, Nagamura Y, Antonio BA, Kanamori H, Hosokawa S, Masukawa M, Arikawa K, Chiden Y, Hayashi M, Okamoto M, Ando K, Aoki H, Arita K, Hamada M, Harada C, Hijishita S, Honda M, Ichikawa Y, Idonuma A, Iijima M, Ikeda M, Ikeno M, Ito S, Ito T, Ito Y, Iwabuchi A, Kamiya K, Karasawa W, Katagiri S, Kikuta A, Kobayashi N, Kono I, Machita K, Maehara T, Mizuno H, Mizubayashi T, Mukai Y, Nagasaki H, Nakashima M, Nakama Y, Nakamichi Y, Nakamura M, Namiki N, Negishi M, Ohta I, Ono N, Saji S, Sakai K, Shibata M, Shimokawa T, Shomura A, Song J, Takazaki Y, Terasawa K, Tsuji K, Waki K, Yamagata H, Yamane H, Yoshiki S, Yoshihara R, Yukawa K, Zhong H, Iwama H, Endo T, Ito H, Hahn JH, Kim H, Eun M, Yano M, Jiang J, Gojobori T (2002) The genome sequence and structure of rice chromosome 1. Nature 420:312–316
Schopfer P (2001) Hydroxyl radical-induced cell-wall loosening in vitro and in vivo: implications for the control of elongation growth. Plant J 28:679–688
Sekimoto H, Hoshi M, Nomura T, Yokota T (1997) Zinc deficiency affects the levels of endogenous gibberellins in Zea mays L. Plant Cell Physiol 38:1087–1090
Skoog F (1940) Relationships between zinc and auxin in the growth of higher plant. Am J Bot 27:939–951
Suge H, Takahashi S, Arita S, Takaki H (1986) Gibberellin relationships in zinc-deficient plants. Plant Cell Physiol 27:1005–1012
Sweetlove LJ, Heazlewood JL, Herald V, Holtzapffel R, Day DA, Millar AH (2002) The impact of oxidative stress on Arabidopsis mitochondria. Plant J 32:891–904
Swire-Clark GA, Marcotte WR Jr (1999) The wheat LEA protein functions as an osmoprotective molecule in Saccharomyces cerevisiae. Plant Mol Biol 39:117–128
Tsui C (1948) The role of zinc in auxin synthesis in tomato plants. Am J Bot 35:172–179
Van Gestelen P, Asard H, Caubergs RJ (1997) Solubilization and separation of a plant plasma membrane NADPH-O2 − synthase from other NAD(P)H oxidoreductases. Plant Physiol 115:543–550
Yoshida KT, Kuboyama T (1997) Isolation of sexual reproduction-related genes in rice by simplified differential display. Rice Genet Newslett 14:126–129
Acknowledgements
We thank Dr. M. Matsuoka for the gift of rice plant materials. This work was supported by a grant from a MAFF Rice Genome Project PR-1215.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H. Uchimiya
Rights and permissions
About this article
Cite this article
Oguchi, K., Tanaka, N., Komatsu, S. et al. Methylmalonate-semialdehyde dehydrogenase is induced in auxin-stimulated and zinc-stimulated root formation in rice. Plant Cell Rep 22, 848–858 (2004). https://doi.org/10.1007/s00299-004-0778-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00299-004-0778-y