
Abstract
Multiprotein bridging factor 1 (MBF1) is a transcriptional co-activator that mediates transcriptional activation by bridging sequence-specific activator like proteins and the TATA-box binding protein (TBP). MBF1 has been well-studied in Arabidopsis thaliana, Saccharomyces cerevisiae, Drosophila melanogaster, and Homo sapiens, but it is not well understood in filamentous fungi. In this study, we report the identification and characterization of a MBF1 ortholog (MoMBF1) in the rice blast fungus Magnaporthe oryzae), which causes the devastating rice blast disease and is an ideal model for studying the growth, development and pathogenic mechanisms of filamentous fungi. MoMBF1 encodes a 161 amino acid protein with a typical MBF1 domain and HTH domain. Bioinformatics were used to analyze the structural domains in MoMBF1 and its phylogenetic relationship to other homologs from different organisms. We have generated MoMBF1 deletion mutants (ΔMoMBF1) and functional complementation transformants, and found that the deletion mutants showed significant defects in vegetative growth and tolerance to exogenous stresses, such as 1 M sorbitol, 0.5 M NaCl, and 5 mM H2O2. Moreover, ΔMoMBF1 showed reduced pathogenicity with smaller infection lesions than wild type and the complementation strain, and decreased response to the accumulation of ROS (reactive oxygen species) in planta at the initial infection stage. Taken together, our data indicate that MoMBF1 is required for vegetative growth, pathogenicity and stress response in M. oryzae.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Multiprotein bridging factor 1 (MBF1), also known as endothelial differentiation-related factor 1 (EDF1), is a common transcriptional co-activator in eukaryotic organisms (De Benedictis et al. 2001; Leidi et al. 2010), linking specific leucine zipper-containing transcription factors and TATA-box binding proteins (TBP) in humans, yeast and Drosophila and enhancing gene transcription (Takemaru et al. 1998). As such, MBF1 plays a significant role in the regulation of eukaryotic gene transcription. The biological function of MBF1 has been studied extensively in eukaryotes such as Arabidopsis thaliana, Saccharomyces cerevisiae, Drosophila melanogaster, and Homo sapiens, but few studies have addressed its function in filamentous fungi (Jindra et al. 2004; Suzuki et al. 2011; Takemaru et al. 1998; Ying et al. 2014). In this regard, Magnaporthe oryzae is an ideal model filamentous fungus that is the pathogen of a major rice disease (rice blast disease) (Li et al. 2014). Thus, the study of the biological function of MBF1 gene in M. oryzae (MoMBF1) is significant in understanding the mechanism of development and pathogenicity of filamentous fungi (Dean et al. 2012; Li et al. 2015). MBF1 is widespread in eukaryotes and archaea and is relatively conserved, but it is absent from Eubacteria (Jindra et al. 2004). It consists of two structural domains: a flexible N terminal region, which binds to various activators, and a C terminal Cro-like Helix Turn Helix (HTH) domain. The major difference in MBF1 between eukaryotes and archaea is the zinc-ribbon (Blombach et al. 2014; de Koning et al. 2009). MBF1 was initially identified and purified from the posterior silk gland of Bombyx mori (Li et al. 1994), and was later found in mammals, Arabidopsis, and yeast. In insects such as silkworms and Drosophila, MBF1 activated transcription of the fushi tarazu (ftz) gene by bridge-linking the transcription activator FTZ-F1 (nuclear hormone receptor) to TBP (Ueda et al. 1992). The ftz gene of Drosophila is specifically expressed during embryogenesis and metamorphosis and involved in body segmentation (Liu et al. 2003).
In plants, MBF1 is involved in plant development and tolerance to environmental stress, which may serve as the primary target gene for signal transduction pathways. There is only one MBF1 gene in most plant, but there are three MBF1 genes in Arabidopsis (At-MBF1a, At-MBF1b, and At-MBF1c) all of which affect growth and phenotype including germination, plant shape, and leaf size. Overexpression of AtMBF1c leads to more seeds and higher tolerance to high temperature, hypertonic stress, and fungal infection (Suzuki et al. 2005, 2008; Tsuda and Yamazaki 2004). Transient expression of MBF1 protein can be induced after treatment of potato fruit with ethylene in early maturity, and in tobacco in response to high temperature and drought stress (Rizhsky et al. 2002). MBF1 protein expression can also be up-regulated after injury, fungal infection, and salicylic acid treatment (Ho and Gasch 2015). In summary, MBF1 is a special co-activator and can enhance transcription by bridge-linking other transcription factors, TBP and specific activators of gene transcription, to regulate various physiological processes within the organism, especially under stress conditions.
In Saccharomyces cerevisiae, Mbf1 and Gcn4 (transcription factor of bZIP) interacted directly with TBP and regulated histidine synthesis by mediating Gcn4 dependent transcription and activation of HIS3 gene (Tian et al. 2007). Deletion of either MBF1 or Gcn4 can reduce the transcription to basal levels imidazoleglycerol-phosphate dehydratase (IGPD, HIS3p), leading to enhanced sensitivity to 3-aminotriazole, an inhibitor of IGPD activity. Recently, Ying et al. identified the MBF1 from the fungal insect pathogen Beauveria bassiana and showed that BbMBF1 acts as a key regulatory cofactor controlling hyphal morphogenesis, stress responses and virulence in B. bassiana (Ying et al. 2014). Aside from S. cerevisiae and Beauveria bassiana, few other fungal MBFs have been identified. Rice is the world’s most widely planted food crop, and its annual yield is severely affected by M. oryzae infection, which highlights economic significance in the study of growth and pathogenicity of M. oryzae, a model organism for study of filamentous fungi (Howard and Valent 1996; Ribot et al. 2008).
Here, we report the characterization of an MBF1 orthologue (MoMBF1) in M. oryzae. Seven MoMBF1-knockout mutant strains have been generated by homologous recombination. Our data indicate that ΔMoMBF1 mutants show reduced mycelial growth and pathogenicity as well as decreased tolerance to exogenous stresses, indicating that MoMBF1 play an important role in the regulation of vegetative growth, pathogenicity, and tolerance to environmental stresses.
Materials and methods
Tested strains and culture conditions
The M. oryzae strain KU80 preserved in the current lab was used as wild type for transformation. MoMBF1-Com (MoMBF1 complementary strains) and ΔMoMBF1 (MoMBF1 knockout mutant of M. oryzae) were generated in this test. All strains were cultured on SYM agar medium (10 g containing soluble starch, 3 g sucrose, 2 g yeast extract, and 18 g agar powder in 1 L medium) for vegetative growth. Liquid complete medium (CM: 6 g yeast extract, 6 g casein hydrolysate, and 10 g sucrose in 1 L medium) was used to prepare the vegetative mycelia for the extraction of DNA and RNA. TB3 medium (containing 3 g yeast extract, 3 g acid hydrolysis casein, 20 g sucrose, and 15 g agar powder in 1 L medium) for regeneration of protoplast. For conidiation, strain blocks were inoculated on Oatmeal agar medium (OMA: oatmeal 20 g, agar powder 20 g in 1 L medium, pH 6.0–6.5) at 28 °C for 7 days in a dark room followed by 3 days of continuous illumination under fluorescent light.
Fungal transformation and disruption of MoMBF1 gene
The gene deletion mutant were generated by the split-marker approach (Yu et al. 2004; Zheng et al. 2012). Sequences of a 716 and 779 bp for the upstream and downstream fragments were amplified with specific primer Mbf1-AF/Mbf1-AR and Mbf1-BF/Mbf1-BR. Primers HYG/F, HY/R, YG/F, and HYG/R were used for amplification of the partial fragments of the hygromycin phosphotransferase (hph) gene as described (Mehrabi et al. 2008). For the transformation, protoplast preparation and transformation were described by Chen et al. (2008). Hygromycin- or neomycin-resistant transformants were selected on media supplemented with 250 g/mL hygromycin B (Roche Applied Science) or 200 g/mL G418 (Invitrogen). Fungal mycelium was collected from SYM medium and lysed at 28 °C for 3 h using lywallzyme solution, after which the suspension was filtered with 2-layer gauze. The filtrate was collected and centrifuged. The supernatant was discarded and protoplast was re-suspended using STC buffer (containing 10 mL, 0.1 M Tris–HCl, 1.1 g anhydrous calcium chloride, and 40 g sucrose in every 200 mL buffer solution). The concentration of protoplast was adjusted to 1 × 107–1 × 108 cell/mL. Then 2 μg target DNA fragment was mixed with 200 μL protoplast for transformation. The mixture was kept on ice for 20 min followed by addition of 5 mL PTC (containing 40 % PEG-modified STC). The mixture was kept at room temperature for 20 min. Then 5 mL TB3 liquid medium was added and it was allowed to regenerate overnight. The next day, this mixture was added to 40 ml dissolved (<50 °C) protoplast regeneration medium (containing 200 µg/mL hygromycin B) and gently stirred. The mixture was inoculated on 15 cm culture medium. After coagulation, upper-level TB3 culture medium containing hygromycin B (200 µg/ml) was added to screen the transformant. The generated transformant was picked and transferred to solid SYM medium to extract DNA for PCR testing. The gene deletion mutants were generated by the split-marker approach (Yu et al. 2004; Zheng et al. 2012).
Screening and molecular validation of the transformants
DNA of the transformants were extracted for PCR validation according to a method described by Talbot et al. (1993). Pairs of primers located within ORF of MoMBF1 (Mbf1-OF/Mbf1-OR), interior primer of hygromycin phosphotransferase (HPt), and upstream primer MoMBF1 (H853/Mbf1-UA) were used in PCR amplification to validate the mutant. In addition, the RT-PCR method (using primer Mbf1-OF/Mbf1-OR) was used to confirm the transformant from the transcriptional level. Primer sequences used in this article are listed in Table 1.
Complementation assay
A 3155 bp sequence covering the ORF of MoMBF1, promoter sequences (1600 bp ahead of ORF) and its 3′ UTR region (noncoding region) (around 500 bp) was amplified by the primers Mbf1-CF/Mbf1-CR. The sequence was connected to pKNTG vector (harboring geneticin-resistance marker) using endonuclease Xho I and Hind III to construct vector with complementary function. The constructs was co-transformed into protoplasts of the target mutant strain. Transformants were selected by growing on plates with geneticin and then screened by PCR (using primer Mbf1-OF/Mbf1-OR) (Primer listed in Table 1).
Yeast complementation assay
The yeast wild-type strain AEY 3087 and mutant strain mbf1Δ were obtained from Dr. Siebers’s laboratory (Coto et al. 2011). The whole open reading frame (ORF) of MoMBF1 was amplified by PCR from the cDNA of the wild-type strain KU80. Then the ORF fragment was cloned into pYES2 vector (Invitrogen). pYES2-MoMbf1 or empty vector pYES2 were transformed into yeast wild-type strain AEY 3087 or mutant strain Δmbf1 as described by the previews report (Coto et al. 2011). The transformants were transferred onto supplemented minimal medium plates with or without 3 mM aminotriazole. The plates were grown in the incubator for 3 days at 30 °C.
Vegetative growth and osmosensitivity test
Wild-type and mutant mycelial pellets of the same diameter were collected with a hole puncher and planted at the center of SYM medium for inverted culture at 28 °C. For the osmosenitivity test, wild-type and mutant mycelial pellets of the same diameter were collected with a hole puncher and planted on SYM plates containing 1 M sorbitol, 0.5 M NaCl, 3 mM, 5 mM H2O2 cultured in the dark at 28 °C to determine their effects on the fungal growth. Colony diameter was measured on day 9 after planting and images were photographed on day 9. The tests were performed in triplicate with three replicates each time.
Conidial morphology, conidium germination, appressorium formation and penetration test
The fungus was cultured on oatmeal agar plates for 7 days then carefully scraped out the aerial hyphae on the surface with sterile slides and light cultured at 28 °C for 2 days. Then the conidia were collected by rinsing with sterile water and the volume was titrated to 1 mL, followed by calculation of the counts of conidia in each dish using a hemocytometer. Counting was performed three times per dish with three biological repeats. Conidia suspension with concentrated of 5 × 104/mL was used for spore germination and appressorium formation test on hydrophobic surfaces of Gelbond film (BMA Company) (Zhang et al. 2009a). Drops 15 µL in volume were generated with three drops on each surface and triplicate were set for each treatment. Spore drops were cultured within humid environments at 28 °C and the spore germination and appressorium formation were observed under microscope at 0.5, 1, 2, 4, 8, and 24 h. Three fields were selected for each observation with three biological repeats. Appressorium penetration on onion epidermal cells was performed as described (Odenbach et al. 2007). Each well of a 24-well culture plate was added to 2 mL distilled water. Three layers of onion inner epidermis of 5 × 5 mm in size were scratched and placed in distilled water. Then 10 µL of conidia suspension at a concentration of 5 × 104/mL suspension was added dropwise to the surface of the onion epidermal cells with 0.4 μM or without diphenyleneiodonium (DPI) dissolved in DMSO, and 10 duplicates were set. Spores were cultured in a humid environment and the infection condition was observed after 24 and 48 h. The experiment was performed three times with three biological repeats.
Pathogenicity assays
The pathogenicity of rice was tested as in a previous report (Talbot et al. 1993). The conidial suspensions were sprayed with solution at a concentration of 2 × 105/mL (containing 0.02 % Tween) on a susceptible rice cultivar Nihonbare at the three-leaf stage. The infected plants were incubated in a humid box for 12 h at 25 °C, then moved to a humid room for 7 days until the symptoms become apparent. The number of lesions on the leaves were counted as described previously (Dong et al. 2015). The pathogenicity test was conducted at least three times. The infected leaves were photographed at day 7.
ROS accumulation assay
Conidia suspensions of a concentration of 1 × 105/mL were inoculated with 1 month old rice leaf sheath of rice cultivar Nihonbare and 6 day old barley plants. The samples were collected at 24 h post inoculation (hpi) and 48 hpi. Diamino-benzidine (DAB) staining was performed following the protocol of Huang et al. (2011). For the rice leaf sheath, the leaf sheath were cut at the base by a razor blade and placed in a 1 mg/ml solution of DAB for 8 h under darkness at room temperature. For the barley leaves, first the leaves were placed in the DAB for 8 h under the same condition with rice leaf sheath, decolorized by boiling the leave with 96 % ethanol for 10 min. Remove the 96 % ethanol and add fresh 100 % ethanol and incubate the samples in room temperature overnight.
Quantitative real-time RT-PCR of pathogenesis-related genes (PR genes)
The infected Rice (cultivar Nipponbare) tissues used for Quantitative real-time reverse transcription PCR (real-time qRT-PCR) analysis were collected at 0, 24, 48, 72 h post inoculation (hpi). qRT-PCR was carried out using primer pairs of the following genes: OsPR1α, OsPBZ1 and OsWRKY53. The actin gene OsActin (Os03g50885) was used as an endogenous control. Total RNA isolated from the frozen plant tissues with Eastep™ Total RNA extraction Kit (Promega (Beijing) Biotech Co., Ltd, LS1030) according to the manufacturer’s instruction. 5 μg of total RNA was reverse transcribed into first-strand cDNA with oligo (dT) primer using GoScript™ Reverse Transcription System (Promega (Beijing) Biotech Co., Ltd Promega, A5001). PCR were performed using a 20 μl reaction of SYBR® Premix Ex Taq™ II (Tli RNaseH Plus) system: 2 × SYBR Premix 10 μl, dH2O 6 μl, Primer(10 μM) 0.8 μl, cDNA 3 μl (25 ng of input RNA). Real-time quantitative RT-PCR was run on the CFX96 Real-Time PCR Detection System (Bio-Rad). Then PCT condition was as follows: 95 °C for 1 min (1 cycle); 95 °C for 25 s, 58 °C for 25 s (40 Cycles); 95 °C for 25 s, 60 °C for 15 s (melting curve). Relative abundance of transcripts was calculated by the 2−ΔΔCt method (Livak and Schmittgen 2001). Quantitative RT-PCR was conducted at least twice with three replicates from independent biological experiments.
Yeast two-hybrid assay
Yeast two-hybrid assay was performed following the manufacturer’s instructions (Matchmaker™ Gold Yeast Two-Hybrid System, Clontech Laboratories, Inc. Cat. No 630489). The whole ORF cDNAs for MoMBF1 and MoGcn4 were cloned into the pGBKT7 and pGADT7 vector, respectively. The constructs were transformed into yeast AH109 strain. Single colonies of the transformants were selected by growing on SD/-Trp-Leu-His plates and SD/-Trp-Leu-His-Ade/X-α-GAL plates.
Results
Sequence alignment and phylogenetic analysis of Mbf1 Proteins from different organisms
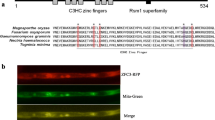
The MoMBF1 gene (MGG_08203) of M. oryzae contains 567 bp, including an 81-bp intron and coding a region for 161-amino acid protein. Based on M. oryzae genome database, there is only one MBF1 gene in M. oryzae. Online Pfam software (http://pfam.xfam.org/) analysis indicated that MoMBF1 contains an N-terminal MBF1 domain and a C terminal HTX domain like all other MBF1 proteins, suggesting the conservation of MBF1 gene in evolution (Fig. 1a). Sequence alignment and phylogenetic analysis were performed using MEGA5.0 software for M. oryzae MoMBF1 (XP_003715182.1), At-MBF1a (NP_565981.1), At-MBF1b (NP_191427.1), and At-MBF1c (NP_189093.1) in A. thaliana, hMBF1α (NP_694880.1) and hMBF1β (NP_003783.1) in humans, MBF1 (NP_014942.4) of Saccharomyces cerevisiae, FgMBF1 (XP_011319121.1) in Fusarium graminearum, NcMBF1 (XP_960690.1) in Neurospora crassa, DmMBF1 (NP_524110.1) in D. melanogaster, ZmMBF1 (NP_001151413.1) in maize, TbMBF1 (ADT84285.1) in Thermococcus barophilus, and BmMBF1 (NP_001036824.1) in silkworms. The results showed two branches of MBF1 genes evolved from ancient bacteria: one in fungi and another in plants and animals (Fig. 1b). Sequence alignment based on the NCBI database indicated homology between MoMBF1 protein and other fungal MBF1 proteins ranging from 40 to 43 % (F. graminearum 42 %; Neurospora crassa 43 %; S. cerevisiae 40 %). The phylogenetic relationship shed light on the evolution and function of MBF1 proteins.

Sequence alignments and analysis of the phylogenetic relationships among MoMBF1 and its homologs. a Sequence alignment of MoMBF1 and its homologs. The black underlining shows the two typical structures: HTH_XRE superfamily and MBF1 superfamily. b A neighbor-joining tree was constructed using the amino acid sequence of MBF1 proteins from different organisms. The tree was constructed using pairwise distance and MEGA5 software (Tamura et al. 2011). (Hs Homo sapiens; At Arabidopsis thaliana; Dm Drosophila melanogaster; Bm Bombyx mori; Tb Thermococcus barophilus; Zm Zea mays; Fg Fusarium graminearum; Nc Neurospora crassa; Sc Saccharomyces cerevisiae; Mg Magnaporthe oryzea)
Complementation of yeast Δmbf1 by MoMBF1
Previous studies revealed that MBF1 is an evolutionarily conserved transcriptional co-activator in other species. However, the function of co-activators from fungus has not been studied so for and the biological role of MBF1 as multiprotein bridging factor has not been demonstrated. To address this issue, complementation study in yeast was performed to test whether MoMBF1 is functional for complementation in yeast mbf1Δ mutant. The yeast mutant lacking MBF1 is viable to grow on galactose or sucrose media, but sensitive to 3-AT, an inhibitor of the His3 gene product. Here, we transformed the pYES2-MoMbf1 or the empty pYES2 vector into the yeast wild-type strain and the yeast mbf1Δ mutant. As expected, the mbf1Δ strain transformed with pYES2-MoMbf1 strain was able to grow in 3 mM 3-AT plate. The mbf1Δ strain with a empty pYES2 vector was sensitive to 3-AT (Fig. 2). Therefore, the results indicated that MoMBF1 like the eukaryotic MBF1 from insects, human, or plants, was able to complement and restore the MBF1 function in yeast.

MoMBF1 complements the S. cerevisiae mbf1Δ mutant. 3 AT sensitivity of yeast wild type (WT), the mbf1Δ mutant strain carrying pYES2-MBF1 (+),the mbf1Δ mutant strain carrying pYES2 empty vector (−). Serial dilutions of the respective strains were placed on minimal medium with or without 3 mM aminotriazole (AT). The plates were grown at 30 °C for 3 days
MoMBF1 deletion and complementation
To investigate the biological function of MoMBF1 in M. oryzae, MoMBF1 knockout mutants were constructed by homologous recombination (Fig. 3a) (Zheng et al. 2012). Targeted deletion mutants were identified by PCR of genomic DNA of the transformants, with two pairs of primers, Mbf-OF/Mbf-OR and Mbf-UA/H853. The candidate knockout mutants were specifically amplified by the Mbf-UA/H853 primers but not by the Mbf-OF/Mbf-OR primers (Fig. 3b). The mutants were further confirmed by RT-RCR to amplify the transcripts of MoMBF1 gene. A total of seven knockout mutants were generated and two mutants, ΔMoMBF1-1 and ΔMoMBF2-2 were randomly selected for further analysis. As indicated in Fig. 3c, the RT-PCR tests did not detect any MoMBF1 transcription in the mutants. Functional complementation strain MoMBF1-Com for one of the mutants was constructed and RT-PCR experiment showed that transcription of MoMBF1 was recovered in the complementation strain (Fig. 3b, c).

Construction and confirmation of the MoMBF1 deletion mutants. a Knockout strategy for MoMBF1. Thick arrows indicate orientation of the MoMBF1 (white) and hygromycin phosphotransferase (hph) gene (gray). Short arrows indicate the primers used in this assay. A long fragment amplified by the SOE-PCR contained the homologous sequences flanking the hph gene will replace the ORF of MoMBF1. b DNA verification of the deletion mutant ΔMoMBF1 and the complementary transformant MoMBF1-Com with the specific primers Mbf1-OF/Mbf1-OR and Mbf1-UA/H853. c Total RNA samples (approximately 1 mg per reaction) isolated from mycelia of WT, ΔMoMBF1 and MoMBF1-Com were subjected to RT-PCR using ΔMoMBF1 gene-specific primers Mbf1-OF and Mbf1-OR (Table 1). The RT-PCR product was a 194 bp fragment in WT and ΔMoMBF1-Com as predicted, but it is absent from the deletion mutant ΔMoMBF1
MoMBF1 is involved in vegetative growth
The ΔMoMBF1 mutants showed normal mycelial morphology but a slower growth rate than wild-type or complementary strains (Fig. 4a). After cultivation under the same conditions for 9 days, the average diameters of the wild-type and complementation strains were 5.36 and 5.28 cm, respectively. In contrast, the average diameter of the two mutants was only 4.81 and 4.86 cm (Fig. 4b), indicating that MoMBF1 is necessary for normal vegetative growth.

Colony morphology and growth rate of MoMBF1 deletion and complementation mutants. a The colonies were cultured on SYM medium for 9 days. b The diameter of the colonies were measured on days 3, 5, 7, and 9 after culture on SYM medium under 28 °C. Error bars represent the standard deviation and asterisks represent highly significant differences (P < 0.01)
MoMBF1 is involved in osmotic adaptation and oxidative stress response
MBF1 proteins have been implicated in plant growth and a number of stress reactions. To determine whether MoMBF1 is also involved in stress response, the growth rate of mutants was assessed in different types of stress environments. The wild-type KU80, ΔMoMBF1, and complemented strains were exposed to different conditions including osmotic stress (1 M sorbitol and 0.5 M NaCl) and oxidative stress (3 mM and 5 mM H2O2). Our results showed more sensitivity of the mutants to both osmotic and oxidative stresses. As indicated in Fig. 5a, b, the osmotic stresses inhibited the growth rate of the mutants ΔMoMBF1-1 and ΔMoMBF1-2 by 30–33 % while the inhibition for the wild-type was less than 20 %. Oxidative stresses also inhibited the growth of the ΔMoMBF1 mutants more strongly than the wild-type strain, which was more obvious with 5 mM H2O2 than 3 mM H2O2. These results indicated that ΔMoMBF1 was highly sensitive to exogenous stress and responsible for the H2O2 tolerance phenotype.

Sensitivities of MoMBF1 deletion and complementation mutants to different exogenous stresses. a The colonies were cultured on SYM medium containing 1 M sorbitol, 0.5 M NaCl, 3 Mm H2O2 and 5 Mm H2O2, respectively. b Growth inhibition rates under different exogenous stresses. Error bars represent the standard deviation and asterisks represent high significant differences (P < 0.01)
Since the MoMBF1 mutant was hypersensitive to osmotic and oxidative stresses, we speculated that it may result from defects in the cell wall composition and function. To investigate this possibility, we tested the integrity of cell walls and membranes of ΔMoMBF1. Mycelial plugs were cultured on SYM agar supplement with 200 μg/ml calcofluor white (CFW) and 200 μg/ml Congo red (CR), which affect the fungal cell wall assembly by binding chitin and β-1, 4-glucans, respectively (Zhang et al. 2010). The mycelial growth rates of ΔMoMBF1 were not severely changed compared with that of wild-type KU80 and complemented strains (Data not showed). These results indicate that MoMBF1 was not involved in cell wall integrity.
MoMBF1 is dispensable in conidial germination and appressorium development
Next, we analyzed conidiation of ΔMoMBF1 and found normal conidiospores and conidia production (Table 2). The spore germination and appressorium formation rate on hydrophobic surface showed no differences between the wild type and mutants (Table 2). These results indicated that MoMBF1 is dispensable for spore germination and appressorium development.
MoMBF1 is required for virulence
To determine whether the virulence of ΔMoMBF1 was altered in comparison to wild-type and complementation strains, susceptible rice cultivar CO39 was inoculated at the three and half leaf stage. After 7 days, wide type and complementary strains produced typical lesions on rice leaves while the ΔMoMBF1 mutant produced much smaller lesions (Fig. 6a), indicating reduced virulence. Statistical analysis showed average 29.67 lesions per 2.4 cm2 of rice leaves by ΔMoMBF1 infection, but the wild-type and complementation strains produced 70.33 and 74.00 lesions, respectively, on average (Fig. 6b). In addition, the wide type and complementation strains produced more pathologically severe lesions at levels 4 and 5 while the ΔMoMBF1 infection produced only lesions at levels 2 and 3 (Valent et al. 1991). The results indicated that the virulence of M. oryzae was significantly reduced upon MoMBF1 disruption.

The pathogenicity of MoMBF1 deletion and complementation mutants on rice cultivar CO39. a Lesions caused by MoMBF1 deletion and complementation mutants at 7 dpi (days post inoculation). b Bar chart of mean lesion density on rice leaves infected with MoMBF1 deletion and complementation mutants per unit area. Error bars represent the standard deviation and asterisks represent highly significant differences (P < 0.01). c Invasive growth of MoMBF1 and complementation mutants in onion epidermal cells at 24 hpi (hours post inoculation). Bar 20 μm. d The percentage of appressorium-mediated penetration and infectious hyphae development of Wild type, MoMBF1 mutant and complementation strains with DPI and without DPI treated onion epidermis. The level of IH development were scored at 24 hpi
To determine the mechanism of reduced virulence in ΔMoMBF1, we investigated the appressorial penetration and invasive growth of the ΔMoMBF1 mutant on onion epidermal cells upon conidial suspension-mediated infection. As showen in Fig. 6c, the appressoria formed by the ΔMoMBF1 mutant appeared normal. However, the penetration rate on onion epidermal cell by ΔMoMBF1 was significantly reduced at 24 h. Only 47.3 % of appressorium of ΔMoMBF1 infected onion skins after 24 h of inoculation, but 72.3 % of wild-type and 70.6 % of complementary strain appressoria were infectious (Fig. 6d). The infection rates of all the strains were reached to 100 % after prolonged incubation time at 48 h post inoculation (Date not show). These results indicated that the appressorial penetration of ΔMoMBF1 was delayed at early infectious stage, which may contribute the reduction of the virulence in rice.
MoMBF1 is essential for overcoming plant defense responses
Stronger host defense response may also contribute to the reduced pathogenicity by the ΔMoMBF1 mutant. Autofluorescence was a hallmark of plant defense response in rice blast disease (Tanabe et al. 2009). We compared the emission of autofluorescence at the primary rice cells infected by the wild-type and the ΔMoMBF1 mutant. A strong autofluorescence was detected in rice cells infected the ΔMoMBF1 mutant (Fig. 7a). In contrast, autofluorescence only appeared in the cell wall of wild-type-infected plants. Autofluorescence at the site of infection reflects the accumulation of phenolic compounds and cell wall strengthening (Shalaby and Horwitz 2015). These results indicated that the ΔMoMBF1 mutant triggers stronger host responses than the wild type at the initial infection stage.

a Fluorescence and DIC microscopy of infected rice sheaths 48 hpi for the autofluorescence at the site of infection. Scale bars 20 μm. b Proportion of DAB stained epidermal cells of the MoMBF1 deletion mutant at 48 hpi. Asterisks indicate significant differences. c The MoMBF1 deletion mutant induced strong plant defense responses in planta. DAB staining was performed on wild type and ΔMoMBF1 conidia 24 and 48 hpi. Compared with wild type, ΔMoMBF1 showed a strong ROS accumulation on plant cell. The experiment were repeated for three times. Images were taken with a microscope. Scale bars 20 μm. d The relative expression of rice pathogenesis-related (PR) genes at early infectious stage by wild type strain and ΔMoMBF1. Samples were collected at 0, 24, 48 and 72 hpi
Another major and early plant defense response is the accumulation of reactive oxygen species (ROS). The conidia of wild type and ΔMoMBF1 mutant were used to inoculate susceptible rice and barley cultivars. The accumulation of hydrogen peroxide (H2O2) at the infection sites was detected by straining with 3,3′-diamino-benzidine (DAB) at 24 and 48 h after inoculation. The ΔMoMBF1-infected samples showed ROS signal at 24 h post inoculation (hpi) and reached maximal ROS signal at 48 hpi compared with the wild type-infected samples (Fig. 7a). The spot-like DAB signals in epidermal cells infected by ΔMoMBF1 were higher than the wild type-infected samples by measuring at 48 hpi (Fig. 7b). As a control, ROS signal was undetectable in non-inoculated leaves under the same conditions (data not shown). These results indicate that the loss of the MoMBF1 gene may abrogate ROS reduction in the host plant cell during early stage of infecion.
To determine whether the inhibition of plant ROS generation can restore infectious hyphae (IH) development of the MoMBF1 mutant, we used 0.4 μM of diphenyleneiodonium (DPI), an inhibitor of NADPH oxidases, to prevent the generation of ROS in the rice cell (Chi et al. 2009). We tested the appressorial penetration and invasive growth of the ΔMoMBF1 mutant on onion epidermal cells with or without DPI (0.04 μM). The results showed that DPI treatment recovered the IH development of ΔMoMBF1 up to 69 % at 24 hpi (Fig. 6d), suggesting that MoMBF1 is involved in overcoming the defense response by degrading oxidases in the plant cell.
Given the fact that the increased ROS accumulation happened in the early stage of infection stage of the ΔMoMBF1, we next determined whether the pathogenesis-related (PR) gene were simulated by the infection of ΔMoMBF1 infection. The expression patterns of three PR genes (PR1, WRKY53 and PBZ1) were examined by quantitative RT-PCR. The results showed the expression of PR1, WRKY53 and PBZ1 was highly induced by ΔMoMBF1 at 24 hpi compared to wild type (Fig. 7b) indicating that the infection of ΔMoMBF1 induced the plant defense responses in planta.
Interaction studies of MoMbf1 and MoGcn4
In S. cerevisiae, a bZIP transcription factor GCN4 cooperate with MBF1 to recruit the TATA-box protein to bind to the promoter region of its target genes. To investigate if MoMBF1 can interact with GCN4 to mediate GCN4-dependent transcriptional activation in M. oryzea, we conducted yeast two-hybrid assay. In M. oryzea, the GCN4 homologue MoGcn4 (MGG_00602) is annotated as a cross-pathway control protein 1 with a ORF of 720-bp interrupted by a 81-bp intron, encoding a 240-amino acid polypeptide that shares 62, 57, 53, 51, 44 % amino acid identity to the homologues in Fusarium fujikuroi (CAP20091.1), Fusarium graminearum PH-1 (XP_011328479.1), Neurospora crassa (XP_957665.1), Saccharomyces cerevisiae (CAE52217.1) and Aspergillus nidulans (AAL09315.1), respectively.
The plasmids pBD-Mbf1 and pAD-Gcn4 were constructed and transformed into the yeast strain AH109, and their interaction was tested by the transcriptional activation of the reporter genes and LacZ. The results showed none of the AH109 Strains carrying both the pBD-Mbf1 and pAD-Gcn4 could grow on a SD/-Trp-Leu-His plate or exhibit blue color on SD/-Trp-Leu-His/X-α-GAL plate (Fig. 8), indicated that no interaction between MoMBF1 and MoGcn4. In addition, the AH109 strain carrying pBD-MoMBF1 did not grow on the SD/-Trp-Leu medium, confirming that MoMBF1 is not a general transcription factor and cannot activate transcription by itself.

MoMBF1 do not interact with MoGcn4 in yeast two-hybrid experiments. The co-transformation of pBD-MBF1 and pAD-Gcn4 can grow on SD/-Trp-Leu plates, but can not grow on SD/-Trp-Leu-His plates. Positive control: Co-transformation of pGBKT7-53 and pGADT7-T. Negative control: Co-transformation of pGBKT7-lam and pGADT7-T. X-Gal: 5-bromo-4-chloro-3-indolyl-β-d-galacto-pyranoside. Blue staining of the yeast colonies indicates the expression of interacting hybrid proteins
Discussion
MBF1 is a highly conserved transcriptional co-activator that is implicated in various physiological and biochemical processes in eukaryotes, including endothelial cell differentiation, lipid metabolism, hormone regulation, development of the central nervous system and histidine metabolism by bridge linking the DNA binding region of transcription factor and TATA-box binding protein (Leidi et al. 2009; Liu et al. 2007). MBF1 proteins in plants control multiple development processes and environmental stress tolerance, and MBF1 deficiency can cause defects in growth and stress tolerance (Mariotti et al. 2000). In Drosophila, MBF1 moves from the cytoplasm to nucleus after binding to the transcription factor D-Jun and the complex stabilizes D-Jun (Jindra et al. 2004). Human MBF1 plays significant role in the differentiation of endothelial cells. However, few studies have addressed MBF1 in fungi, especially filamentous fungi. One recent study showed that colony morphology can be affected by lack of BbMBF1 in the fungal pathogen B. bassiana, with increased sensitivity to osmotic pressure and reduced pathogenicity (Ying et al. 2014). In this study, we functionally characterized the MBF1 ortholog in M. oryzae (MoMBF1). Based on bioinformatic analyses, Mbf1 is widely distributed and highly conserved in eukaryotes and archaea. Previous studies reported that the MBF1 homologs from Arabidopsis, potato, silk worm and human can complement the MBF1 function in the yeast deletion strains (mbf1Δ) (Coto et al. 2011; Suzuki et al. 2008; Zanetti et al. 2003). In this study, we found that MBF1 from M. oryzae was also able to complement the MBF1 function in yeast. Like the function of MBF1 in B. bassiana, MoMBF1 is required for vegetative growth, pathogenicity as well as response to exogenous stresses in M. oryzae.
MBF1 genes have been implicated in exogenous stress and oxidative stress responses in different organisms (Arce et al. 2010; Suzuki et al. 2005). Consistently, our study has demonstrated the significance of MoMBF1 in response to the exogenous stresses. The MoMBF1 deletion mutants are defective in mycelial growth and more sensitive to inhibition by osmotic and oxidative stresses than the wild-type. Indeed, this defect is due to MoMBF1 deficiency because the MoMBF1 complementation transformant can completely restore the wild-type phonotype. Thus, MoMBF1 plays an important role in mycelia growth and in the response to exogenous stresses. In eukaryotic cells, the mitogen-activated protein kinases (MAPK) are involved in transducing a variety of extracellular signals and regulating growth and differentiation processes (Xu 2000). Previous report showed the HOG pathway related genes, Hog1, Pbs2, Ste11, Ssk2 and Ssk22, are essential for maintaining the osmotic gradient across the plasma membrane by accumulating glycerol in M. oryzae and other fungi (Thines et al. 2000; Zhang et al. 2009b; Zhao et al. 2005). The two-component histidine kinases genes and the ABC transporter genes of blast fungus are involved in the osmotic stress response and pathogenicity (Kim et al. 2013; Motoyama et al. 2008; Zhang et al. 2010). It was reported, the ABC and major facilitator superfamily (MFS) transporters, the histidine kinases genes, and MAPK pathway genes in B. bassiana were regulated by BbMBF1. Therefor, these genes may also be regulated by MoMBF1 in M. oryzae to respond osmotic and oxidative stresses.
We have also found that MoMBF1 contributes to the virulence of M. oryzae in rice by regulation of appressorial penetration and invasive growth and by blocking ROS accumulation in planta. We speculate that MoMBF1 may control the expression of genes involved in degradation of ROS-producing enzymes. In support of this contention, we have detected robust induction of the PR genes (WRKY53, PBZ1, and PR1α) in ΔMoMBF1-challenged rice tissues. Taken together, our data indicate that MoMBF1 plays a crucial role in the virulence of the rice blast fungus.
In S. cerevisiae, the bZIP transcription factor Gcn4p is a key regulator in response to the amino acid starvation (Hinnebusch and Natarajan 2002; Natarajan et al. 2001). In yeast, a GST pull-down assays showed Mbf1 interacts with Gcn4 and a TATA-box binding protein (TBP) directly, mediating the Gcn4-dependent transcriptional activation of the HIS3 gene, which encodes imidazole-3-phosphate dehydratase. However, in our research, a yeast two-hybrid experiment showed Mbf1 and Gcn4 proteins do not interact with each other in M. oryzea. Similar results was reported in F. fujikuroi, no interaction was detected between Cpc1(GCN4 ortholog) and Mbf1 using two different yeast two-hybrid system (Schonig et al. 2009). The pull-down or co-immunoprecipitation assays may need to be performed to determine the interaction between MBF1 and GCN4 in M. oryzae.
Transcription factor are key regulators of gene expression and play an important role in diverse processes, including developmental control and initiation of stress and defense responses (Liu et al. 2014). Our study indicates that the transcription co-activator MBF1 regulates vegetative growth, pathogenicity, and stress response in the rice blast fungus M. oryzae.
References
Arce DP, Godoy AV, Tsuda K, Yamazaki K, Valle EM, Iglesias MJ, Di Mauro MF, Casalongue CA (2010) The analysis of an Arabidopsis triple knock-down mutant reveals functions for MBF1 genes under oxidative stress conditions. J Plant Physiol 167:194–200. doi:10.1016/j.jplph.2009.09.003
Blombach F, Launay H, Snijders AP, Zorraquino V, Wu H, de Koning B, Brouns SJ, Ettema TJ, Camilloni C, Cavalli A, Vendruscolo M, Dickman MJ, Cabrita LD, La Teana A, Benelli D, Londei P, Christodoulou J, van der Oost J (2014) Archaeal MBF1 binds to 30S and 70S ribosomes via its helix-turn-helix domain. Biochem J 462:373–384. doi:10.1042/BJ20131474
Chen J, Zheng W, Zheng S, Zhang D, Sang W, Chen X, Li G, Lu G, Wang Z (2008) Rac1 is required for pathogenicity and Chm1-dependent conidiogenesis in rice fungal pathogen Magnaporthe grisea. PLoS Pathog 4:e1000202. doi:10.1371/journal.ppat.1000202
Chi MH, Park SY, Kim S, Lee YH (2009) A novel pathogenicity gene is required in the rice blast fungus to suppress the basal defenses of the host. PLoS Pathog 5:e1000401. doi:10.1371/journal.ppat.1000401
Coto JM, Ehrenhofer-Murray AE, Pons T, Siebers B (2011) Functional analysis of archaeal MBF1 by complementation studies in yeast. Biol Direct 6:18. doi:10.1186/1745-6150-6-18
De Benedictis L, Mariotti M, Dragoni I, Maier JA (2001) Cloning and characterization of murine EDF-1. Gene 275:299–304
de Koning B, Blombach F, Wu H, Brouns SJ, van der Oost J (2009) Role of multiprotein bridging factor 1 in archaea: bridging the domains? Biochem Soc Trans 37:52–57. doi:10.1042/BST0370052
Dean R, Van Kan JA, Pretorius ZA, Hammond-Kosack KE, Di Pietro A, Spanu PD, Rudd JJ, Dickman M, Kahmann R, Ellis J, Foster GD (2012) The top 10 fungal pathogens in molecular plant pathology. Mol plant Pathol 13:414–430. doi:10.1111/j.1364-3703.2011.00783.x
Dong Y, Li Y, Zhao M, Jing M, Liu X, Liu M, Guo X, Zhang X, Chen Y, Liu Y, Liu Y, Ye W, Zhang H, Wang Y, Zheng X, Wang P, Zhang Z (2015) Global genome and transcriptome analyses of Magnaporthe oryzae epidemic isolate 98-06 uncover novel effectors and pathogenicity-related genes, revealing gene gain and lose dynamics in genome evolution. PLoS Pathog 11:e1004801. doi:10.1371/journal.ppat.1004801
Hinnebusch AG, Natarajan K (2002) Gcn4p, a master regulator of gene expression, is controlled at multiple levels by diverse signals of starvation and stress. Eukaryot Cell 1:22–32
Ho YH, Gasch AP (2015) Exploiting the yeast stress-activated signaling network to inform on stress biology and disease signaling. Curr Genet 61:503–511. doi:10.1007/s00294-015-0491-0
Howard RJ, Valent B (1996) Breaking and entering: host penetration by the fungal rice blast pathogen Magnaporthe grisea. Annu Rev Microbiol 50:491–512. doi:10.1146/annurev.micro.50.1.491
Huang K, Czymmek KJ, Caplan JL, Sweigard JA, Donofrio NM (2011) HYR1-mediated detoxification of reactive oxygen species is required for full virulence in the rice blast fungus. PLoS Pathog 7:e1001335. doi:10.1371/journal.ppat.1001335
Jindra M, Gaziova I, Uhlirova M, Okabe M, Hiromi Y, Hirose S (2004) Coactivator MBF1 preserves the redox-dependent AP-1 activity during oxidative stress in Drosophila. EMBO J 23:3538–3547. doi:10.1038/sj.emboj.7600356
Kim Y, Park SY, Kim D, Choi J, Lee YH, Lee JH, Choi W (2013) Genome-scale analysis of ABC transporter genes and characterization of the ABCC type transporter genes in Magnaporthe oryzae. Genomics 101:354–361. doi:10.1016/j.ygeno.2013.04.003
Leidi M, Mariotti M, Maier JA (2009) Transcriptional coactivator EDF-1 is required for PPARgamma-stimulated adipogenesis. Cell Mol Life Sci CMLS 66:2733–2742. doi:10.1007/s00018-009-0069-4
Leidi M, Mariotti M, Maier JA (2010) The effects of silencing EDF-1 in human endothelial cells. Atherosclerosis 211:55–60. doi:10.1016/j.atherosclerosis.2010.01.022
Li FQ, Ueda H, Hirose S (1994) Mediators of activation of fushi tarazu gene transcription by BmFTZ-F1. Mol Cell Biol 14:3013–3021
Li C, Yang J, Zhou W, Chen XL, Huang JG, Cheng ZH, Zhao WS, Zhang Y, Peng YL (2014) A spindle pole antigen gene MoSPA2 is important for polar cell growth of vegetative hyphae and conidia, but is dispensable for pathogenicity in Magnaporthe oryzae. Curr Genet 60:255–263. doi:10.1007/s00294-014-0431-4
Li Y, Que Y, Liu Y, Yue X, Meng X, Zhang Z, Wang Z (2015) The putative Ggamma subunit gene MGG1 is required for conidiation, appressorium formation, mating and pathogenicity in Magnaporthe oryzae. Curr Genet 61:641–651. doi:10.1007/s00294-015-0490-1
Liu QX, Jindra M, Ueda H, Hiromi Y, Hirose S (2003) Drosophila MBF1 is a co-activator for Tracheae defective and contributes to the formation of tracheal and nervous systems. Development 130:719–728
Liu QX, Nakashima-Kamimura N, Ikeo K, Hirose S, Gojobori T (2007) Compensatory change of interacting amino acids in the coevolution of transcriptional coactivator MBF1 and TATA-box-binding protein. Mol Biol Evol 24:1458–1463. doi:10.1093/molbev/msm073
Liu W, Liu J, Triplett L, Leach JE, Wang GL (2014) Novel insights into rice innate immunity against bacterial and fungal pathogens. Annu Rev Phytopathol 52:213–241. doi:10.1146/annurev-phyto-102313-045926
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-ΔΔC(T)) method. Methods 25:402–408. doi:10.1006/meth.2001.1262
Mariotti M, De Benedictis L, Avon E, Maier JA (2000) Interaction between endothelial differentiation-related factor-1 and calmodulin in vitro and in vivo. J Biol Chem 275:24047–24051. doi:10.1074/jbc.M001928200
Mehrabi R, Ding S, Xu JR (2008) MADS-box transcription factor mig1 is required for infectious growth in Magnaporthe grisea. Eukaryot Cell 7:791–799. doi:10.1128/EC.00009-08
Motoyama T, Ochiai N, Morita M, Iida Y, Usami R, Kudo T (2008) Involvement of putative response regulator genes of the rice blast fungus Magnaporthe oryzae in osmotic stress response, fungicide action, and pathogenicity. Curr Genet 54:185–195. doi:10.1007/s00294-008-0211-0
Natarajan K, Meyer MR, Jackson BM, Slade D, Roberts C, Hinnebusch AG, Marton MJ (2001) Transcriptional profiling shows that Gcn4p is a master regulator of gene expression during amino acid starvation in yeast. Mol Cell Biol 21:4347–4368. doi:10.1128/MCB.21.13.4347-4368.2001
Odenbach D, Breth B, Thines E, Weber RW, Anke H, Foster AJ (2007) The transcription factor Con7p is a central regulator of infection-related morphogenesis in the rice blast fungus Magnaporthe grisea. Mol Microbiol 64:293–307. doi:10.1111/j.1365-2958.2007.05643.x
Ribot C, Hirsch J, Balzergue S, Tharreau D, Notteghem JL, Lebrun MH, Morel JB (2008) Susceptibility of rice to the blast fungus, Magnaporthe grisea. J Plant Physiol 165:114–124. doi:10.1016/j.jplph.2007.06.013
Rizhsky L, Liang H, Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco. Plant Physiol 130:1143–1151. doi:10.1104/pp.006858
Schonig B, Vogel S, Tudzynski B (2009) Cpc1 mediates cross-pathway control independently of Mbf1 in Fusarium fujikuroi. Fungal Genet Biol FG & B 46:898–908. doi:10.1016/j.fgb.2009.08.003
Shalaby S, Horwitz BA (2015) Plant phenolic compounds and oxidative stress: integrated signals in fungal–plant interactions. Curr Genet 61:347–357. doi:10.1007/s00294-014-0458-6
Suzuki N, Rizhsky L, Liang H, Shuman J, Shulaev V, Mittler R (2005) Enhanced tolerance to environmental stress in transgenic plants expressing the transcriptional coactivator multiprotein bridging factor 1c. Plant Physiol 139:1313–1322. doi:10.1104/pp.105.070110
Suzuki N, Bajad S, Shuman J, Shulaev V, Mittler R (2008) The transcriptional co-activator MBF1c is a key regulator of thermotolerance in Arabidopsis thaliana. J Biol Chem 283:9269–9275. doi:10.1074/jbc.M709187200
Suzuki N, Sejima H, Tam R, Schlauch K, Mittler R (2011) Identification of the MBF1 heat-response regulon of Arabidopsis thaliana. Plant J Cell Mol Biol 66:844–851. doi:10.1111/j.1365-313X.2011.04550.x
Takemaru K, Harashima S, Ueda H, Hirose S (1998) Yeast coactivator MBF1 mediates GCN4-dependent transcriptional activation. Mol Cell Biol 18:4971–4976
Talbot NJ, Ebbole DJ, Hamer JE (1993) Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast fungus Magnaporthe grisea. Plant Cell 5:1575–1590. doi:10.1105/tpc.5.11.1575
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28:2731–2739. doi:10.1093/molbev/msr121
Tanabe S, Nishizawa Y, Minami E (2009) Effects of catalase on the accumulation of H(2)O(2) in rice cells inoculated with rice blast fungus, Magnaporthe oryzae. Physiol Plant 137:148–154. doi:10.1111/j.1399-3054.2009.01272.x
Thines E, Weber RW, Talbot NJ (2000) MAP kinase and protein kinase A-dependent mobilization of triacylglycerol and glycogen during appressorium turgor generation by Magnaporthe grisea. Plant Cell 12:1703–1718
Tian C, Kasuga T, Sachs MS, Glass NL (2007) Transcriptional profiling of cross pathway control in Neurospora crassa and comparative analysis of the Gcn4 and CPC1 regulons. Eukaryot Cell 6:1018–1029. doi:10.1128/EC.00078-07
Tsuda K, Yamazaki K (2004) Structure and expression analysis of three subtypes of Arabidopsis MBF1 genes. Biochim Biophys Acta 1680:1–10. doi:10.1016/j.bbaexp.2004.08.004
Ueda H, Sun GC, Murata T, Hirose S (1992) A novel DNA-binding motif abuts the zinc finger domain of insect nuclear hormone receptor FTZ-F1 and mouse embryonal long terminal repeat-binding protein. Mol Cell Biol 12:5667–5672
Valent B, Farrall L, Chumley FG (1991) Magnaporthe grisea genes for pathogenicity and virulence identified through a series of backcrosses. Genetics 127:87–101
Xu JR (2000) Map kinases in fungal pathogens. Fungal Genet Biol FG & B 31:137–152. doi:10.1006/fgbi.2000.1237
Ying SH, Ji XP, Wang XX, Feng MG, Keyhani NO (2014) The transcriptional co-activator multiprotein bridging factor 1 from the fungal insect pathogen, Beauveria bassiana, mediates regulation of hyphal morphogenesis, stress tolerance and virulence. Environ Microbiol 16:1879–1897. doi:10.1111/1462-2920.12450
Yu JH, Hamari Z, Han KH, Seo JA, Reyes-Dominguez Y, Scazzocchio C (2004) Double-joint PCR: a PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genet Biol FG & B 41:973–981. doi:10.1016/j.fgb.2004.08.001
Zanetti ME, Blanco FA, Daleo GR, Casalongue CA (2003) Phosphorylation of a member of the MBF1 transcriptional co-activator family, StMBF1, is stimulated in potato cell suspensions upon fungal elicitor challenge. J Exp Bot 54:623–632
Zhang H, Zhao Q, Liu K, Zhang Z, Wang Y, Zheng X (2009a) MgCRZ1, a transcription factor of Magnaporthe grisea, controls growth, development and is involved in full virulence. FEMS Microbiol Lett 293:160–169. doi:10.1111/j.1574-6968.2009.01524.x
Zhang Y, Zhao J, Fang W, Zhang J, Luo Z, Zhang M, Fan Y, Pei Y (2009b) Mitogen-activated protein kinase hog1 in the entomopathogenic fungus Beauveria bassiana regulates environmental stress responses and virulence to insects. Appl Environ Microbiol 75:3787–3795. doi:10.1128/AEM.01913-08
Zhang H, Liu K, Zhang X, Song W, Zhao Q, Dong Y, Guo M, Zheng X, Zhang Z (2010) A two-component histidine kinase, MoSLN1, is required for cell wall integrity and pathogenicity of the rice blast fungus, Magnaporthe oryzae. Curr Genet 56:517–528. doi:10.1007/s00294-010-0319-x
Zhao X, Kim Y, Park G, Xu JR (2005) A mitogen-activated protein kinase cascade regulating infection-related morphogenesis in Magnaporthe grisea. Plant Cell 17:1317–1329. doi:10.1105/tpc.104.029116
Zheng W, Zhao X, Xie Q, Huang Q, Zhang C, Zhai H, Xu L, Lu G, Shim WB, Wang Z (2012) A conserved homeobox transcription factor Htf1 is required for phialide development and conidiogenesis in Fusarium species. PLoS One 7:e45432. doi:10.1371/journal.pone.0045432
Acknowledgments
This work was funded by a National Science Foundation of China (31571943). We thank Dr. Bettina Siebers from University of Duisburg Essen for kindly providing the yeast strains.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M. Kupiec.
Rights and permissions
About this article

Cite this article
Fan, G., Zhang, K., Huang, H. et al. Multiprotein-bridging factor 1 regulates vegetative growth, osmotic stress, and virulence in Magnaporthe oryzae . Curr Genet 63, 293–309 (2017). https://doi.org/10.1007/s00294-016-0636-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00294-016-0636-9