
Abstract
Two bacterial consortia capable of degrading SLES were isolated from a wastewater treatment plant. The two consortia consisted of three members, Acinetobacter calcoacetiacus and Klebsiella oxytoca in one co-culture (A-K) and Serratia odorifera in the second co-culture (S-A), which contains Acinetobacter calcoacetiacus as well. In all experiments, cells were grown on SLES (1000–7000 ppm) containing the M9 minimal medium as sole carbon source. The co-culture A-K demonstrated a higher growth rate (0.26 h−1) and significant greater viability than that of the co-culture S-A (0.21 h−1). Glucose, sucrose, maltose, mannitol, and succinic acid as carbon sources produced the same degradation rate (∼100 ppm/h) and enhanced the SLES degradation rate by 3-fold upon the control (without an added carbon source). In the case of the co-culture S-A, the situation was different; all the carbon sources being tested except maltose caused a repression in the degradation ability in a range between 25–100%. Maltose causes an enhancement by almost fivefold, compared with the positive control.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Sodium lauryl ether sulfate (SLES) are widely used as surfactants in the formulating of detergent products [1–3]. As they are used largely in household and industrial detergents, SLES is discharged into wastewater collection systems worldwide [4–6]. Sodium lauryl ether sulfate consists of straight carbon chains, usually 12 to 14, and, due to its more simple structure (i.e., no benzene ring), it can be more easily biodegradable than other surfactants such as LAS [6]. Although many studies have dealt with the fate of anionic surfactants in municipal wastewater treatment plants and the fate of SLES discharged to the environment, there have been few reports on the biodegradation of these compounds at high concentrations, such as those present in wastewaters generated from surfactant manufacturing.
The biodegradability of surfactants could be an additional criterion for evaluating such products ( SLES). Incomplete biodegradation of surfactants has led to the development of great masses of foams in streams and rivers in the vicinity of dams or their obstructions [7]. Until now, no information has been available on the complete degradation of high concentrations of these surfactants [8]. The interest is, thus, not whether surfactants are degraded, but under which conditions and by which specific bacteria such degradation takes place.
The aim of this work was to study the ability of two co-cultures as well as their individual isolates for the biodegradability of surfactants (SLES) under different conditions. These co-cultures were: (1) Acinetobacter calcoacetiacus and Klebsiella oxytoca (A-K); and (2) Serratia odorifera and Acinetobacter calcoacetiacus (S-A) isolated from a wastewater treatment plant [9].
Materials and Methods
Materials
Sodium lauryl ether sulfonate (SLES) was supplied by Jordan Sulpho-Chemicals Co. Ltd., Jordan. The SLES was in aqueous sodium salt solutions with minimum purities of 98%, which was used in the preparation of working standards. Benzethoinium chloride (hyamine) and patent blue (disulfide blue) were obtained from Acros Organics (Fisher Scientific, UK). Most of the chemicals used were either from Sigma, USA, or from Fluka Chemika, Switzerland. Other chemicals were analytical grade and were obtained from commercial suppliers.
Bacterial Isolates
Three previous wastewater isolates [9] were tested in single and co-cultures for the biodegradability of SLES. These isolates and their different combinations are shown in Table 1. The co-cultures used for further experiments were: (1) Acinetobacter calcoacetiacus and Klebsiella oxytoca, denoted A-K; and (2) Serratia odorifera and Acinetobacter calcoacetiacus, denoted S-A. Their morphological characteristics were re-verified and their biochemical identity was determined using the REMEL kit (RapIDTM ONE and RapIDTM NF plus systems) procedure.
Growth
In all experiments, acid-washed glassware was used. In all experiments conducted, the surfactants containing uninoculated media were taken as control. The nutrient broth (NB) and agar medium (NA) were used for the isolation of bacteria. The minimal broth media (M9) as described by Miller [10] with slight modifications (consisting of the following: 3.0 g Na2HPO4, 1.5 g KH2PO4, 1.0 g NH4Cl, and 0.5 g NaCl) were dissolved in 500 mL distilled water and the pH adjusted to 7.4 with 6 N NaOH. Then the following compounds were added: 0.24 g MgSO4, 0.05 g CaCl2.6H2O, and 0.05 g FeCl3.6H2O. After autoclaving, the thiamine-HCl (0.001 g) was added by predissolving it in 2 mL distilled water and sterilizing by filtration through a 0.45-μm Millipore filter. SLES was used in concentrations of 3000 ppm (otherwise will be stated), as carbon and energy sources.
The initial cell mass (starting inocula) for each experiment on the M9 medium was grown from single colonies in two consecutive shake flask cultures (overnight followed by 3 h recovery) in nutrient broth. Cells from the 3-hour cultures were harvested by centrifugation and then washed with and resuspended in NB at a concentration of 0.5 OD600 units. One milliliter of these bacterial cells was used to inoculate 100 ml of SLES containing the M9 minimal medium in a 250-ml Erlenmeyer flask for shake flask experiments. In the case of the mixed cultures (A-K and S-A), 1-ml inocula of the two bacterial cells have been always taken to constitute a 50:50 co-culture (the volume adjusted to give a constant OD600, taking into consideration the total number of bacterial cells per OD600) [11].
Analytical Methods
SLES was analyzed in culture media using the two-phase titration method [12], modified by Li and Rosen [13]. The SLES was extracted in an organic solvent (chloroform) and titrated against standardized hyamine (benzethonium chloride) in the presence of a patent (anionic dye). Hyamine solutions were prepared by dissolving 1.86 g l−1 in the case of SLES titration. The endpoint was detected by the pink to blue oily droplets of colour development of the organic layer.
The average degradation rates of SLES were measured by dividing the net amount of degraded SLES for 24 h, since many cells showed no further degradation after 24 h. The reason for calculating the average degradation by this method, as suggested by Loh and Wang [14], was to avoid any errors caused by different lengths of lag phases, and the difficulty in ascertaining the time required to achieve complete degradation or when the degradation had stopped.
Effect of Different Carbon and Nitrogen Sources
Different carbon and nitrogen sources were supplied to the 3000-ppm SLES-containing M9 minimal medium to study their effect on the biodegradation of SLES as a function of time. The carbon sources supplemented independently were glucose, sucrose, maltose, mannitol, and succinic acid, whereas the nitrogen sources being tested included ammonium chloride, trypton, yeast extract, ammonium nitrate, and casein. All of the carbon and nitrogen sources were supplied independently at a concentration of 0.2% to the SLES-containing M9 minimal medium. The succinate was omitted from the original M9 minimal media when studying the effect of carbon sources and the same was made in the case of nitrogen sources omitting the ammonium chloride.
Growth Curves
The SLES-containing minimal medium (M9) was used for growth curves. The medium was supplemented with 3000 mg l−1 SLES as the only carbon source utilized. Growth curves experiments were performed for both co-cultures under the same conditions. The same initial cell mass (starter inocula) for all cases was taken into consideration as mentioned above. Samples were then taken at time intervals and assayed for cell mass OD600 and number of cells per ml by plating on the nutrient agar medium (NA).
Determination of Oxygen Utilization
Measurement of oxygen utilization was performed according to the method described by Khosravi et al. [15]. Respiration of the whole cells was measured polarographically with a Yellow Springs Instruments Model 53 oxygen monitor at room temperature, using an air-saturated 0.1 M potassium phosphate buffer at pH 7. Before measuring the oxygen uptake, the buffer was stirred for 2 h to be air saturated (DO = 250 μM). One milliliter of bacterial culture (cfu was simultaneously determined by plating on nutrient agar) was centrifuged at 10,000g for 5 min and washed twice with 0.1 M potassium phosphate, pH 7, and the resulting pellet added quantitatively to 4 ml air-saturated buffer. The remaining concentration of oxygen in the cell-containing buffer was monitored by an oxygen electrode and recorded for 5 min. Oxygen uptake measurements were made at 10–15-hour intervals and the experiments were repeated 5 times.
Results and Discussions
In this study, two different co-cultures were isolated from a wastewater treatment plant and found to be able to degrade the sodium lauryl ether sulfate (SLES). These co-cultures were: (1) Acinetobacter calcoacetiacus and Klebsiella oxytoca (A-K); and (2) Serratia odorifera and Acinetobacter calcoacetiacus (S-A). The complementary action of strains in breaking down SLES was examined in liquid cultures (Table 1). Reconstitution with all possible combinations of two or three members of the consortium was made. It is clear that the individual bacterial cells produce different effects from their combinations on the level of SLES degradation. Our results suggest, however, that basal expression of SLES degradation activity occurs in the three individual bacterial strains.
Thus, this work includes two different perspectives: firstly, finding these co-cultures as surfactant (SLES) degraders and, secondly, making a comparison in the degrading ability of SLES between the two co-cultures, under different growth conditions.
Effect of Substrate Concentration
Using SLES as sole carbon and energy sources (Fig. 1) also supported our results in that the formation of any biomass is the function of SLES exhaustion. This study may represent the first report of SLES removal by the two co-cultures in such higher concentrations. When both A-K and S-A co-cultures were tested on different SLES concentrations (Fig. 1), the maximum biodegradation rates of SLES for the two co-cultures were 37.3 and 20.52 ppm/h, respectively. The maximal biodegradation rates for A-K and S-A co-cultures were achieved at initial SLES concentration of 4000 and 3000 ppm, respectively. Therefore, a fixed concentration of 3000 ppm SLES was used in the further experiments to compare between the two co-cultures. This SLES degradation was completed (100%) within a concentration range of 1,000–3,000 ppm in 96 hours, whereas higher concentrations of SLES (5,000–7,000 ppm) were completely degraded over a longer incubation time (120–144 hours). This could be interpreted as the higher concentration of SLES possibly leading to a decrease in substrate bioavailability, which slows down its removal. In contrast, the co-culture S-A completely degraded the SLES within 120–144 h, indicating that the surfactant removal rate in the A-K was faster than that in the co-culture S-A. More direct evidence was shown by plotting the initial SLES concentration in the growth media and viable cells for both co-cultures within the first 24 hours upon exposure to different concentrations of surfactant (Fig. 2). The results showed that the viable cell number was increased until the SLES concentration of 4000 and 3000 ppm for A-K and S-A co-cultures, respectively. Due to its simpler structure (i.e., no benzene ring), SLES can undergo rapid complete mineralization compared with other surfactants [16]. Based on the effect of SLES concentration on the cell number, the specific growth rate, μ, can be determined experimentally having the ratio of the difference in the cell concentration to the differences in time according to Nuhoglu and Yalcin [17]

Effect of different substrate concentrations on the biodegradation rate of SLES (ppm/h) by co-cultures A-K and S-A. The culture media used were minimal media (M9) containing SLES as carbon sources only. Culture was incubated at 37°C and under a 150-rpm shaking rate at pH 7.5. The control consisted of uninoculated broth plus 3000 ppm SLES. The data are average of three repeated experiments. Error bars, indicating standard deviations, have been included for all points, but in many cases are smaller than the diameter of the points.

Effect of substrate concentration (SLES) on the viable cell number of the two co-cultures A-K and S-A. The cells were grown on M9 minimal media containing a corresponding initial concentration of SLES. Data are the average of three repeated experiments. Error bars, indicating standard deviations, have been included for all points, but in many cases are smaller than the diameter of the points.
The growth rates achieved for A-K and A-S co-cultures were 0.26 h−1 and 0.21 h−1, respectively. A mixed culture of activated sludge from a municipal wastewater treatment plant grown on higher SDS concentrations (500–2500 mg/l) increased the microbial specific growth rate in comparison to cultures incubated with lower SDS concentrations (in the range of 0.0379 to 0.0567 h−1) [18]. This finding supported our results in which the high concentrations (7000 ppm) of SLES showed no toxic effect on the two co-cultures being tested although they had different degradation abilities.
Oxygen Uptake
The biodegradation rate of SLES by both co-cultures was investigated by measuring the substrate (SLES)-dependent O2 uptake on the SLES-containing minimal medium as the carbon source (Fig. 3). These results were consistent with the pattern observed for SLES removal in which the pattern of oxygen consumption per 108 cell was clearly predominant in the early stages (prior to 72 hours) of incubation time. These data demonstrate that aerobic biodegradation of SLES is a feasible process for SLES removal. The co-culture S-A demonstrated lower respiration than the A-K at almost every point in the growth curve except the period between 36–48 hours of incubation time, the difference being greatest early and, to a lesser extent, late. However, in the co-culture A-K, the oxygen uptake occurred not only in a higher level than in the co-culture S-A, but there was also a shift in the peak of the oxygen uptake level from 36 to 48 hours. The individual bacterial cultures were similar in having lower respiration than that of the consortium. The respiration in both co-cultures was much less sensitive to surfactants than the respiration in other heterotrophic microorganisms [3]. The reason for the low sensitivity of both co-cultures to SLES, compared to the sensitivity of heterotrophic bacteria, is unknown.

O2 uptake rate of co-cultures A-K and S-A as a function of times in M9 containing 3000 ppm of SLES. Oxygen consumption (y axis) is in μmol O2 min−1 (108 cell)−1. The data are the average of five repeated experiments. Error bars, indicating standard deviations, have been included for all points, but in many cases are smaller than the diameter of the points.
Growth Curves
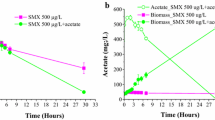
When the growth of the co-cultures (A-K and S-A) was compared using SLES (3000 ppm) as the only carbon sources in the M9 minimal medium (Fig. 4a and b), the maximum growth rates on OD600 measurements of the two consortia were about the same, but co-culture A-K still remained slightly higher (about 10%) in the cell mass of the stationary phase (from 96–144 h), compared with the other co-culture in prolonged culture (Fig. 4a). However, based on viable cell accounts (Fig. 4b), A-K demonstrated significantly greater viability in almost all stages compared with that observed in co-culture S-A.

Optical density at 600 nm (a) and viable cells (b) of co-cultures (A-K and S-A) and their individuals as a function of time in M9 containing 3000 ppm of SLES. Data are the average of four (a) and six (b) repeated experiments. Error bars, indicating standard deviations, have been included for all points, but in some cases they are smaller than the diameters of the points.
Effect of Carbon and Nitrogen Sources
The different carbon and nitrogen sources added in the addition of SLES as carbon sources also caused different effects on the degradation ability (Table 2). When two co-cultures (A-K and S-A) were grown on SLES as the sole carbon source, different degradation rates (30.33 and 20.50, respectively) were achieved. Glucose, sucrose, maltose, mannitol, and succinic acid as carbon sources produced almost the same degradation rate (49.50 ppm/h) and enhanced the SLES degradation rate by 3-fold upon the control (without added carbon source). In the case of the co-culture S-A, the situation is different; all the carbon sources being tested except maltose caused a repression in the degradation ability in the range between 25–100%. Unexpectedly, the maltose caused an enhancement by almost fivefold compared with the positive control. These carbon sources are probably more readily assimilable by the co-culture S-A than the SLES as carbon sources being firstly utilized, compared with the other carbon sources being tested in the A-K co-culture. If we consider the increase in SLES degradation rate as a result of increasing cell biomass, the decrease in SLES degradation rate is thus a result of catabolic repression since no growth inhibition was shown with all carbon and nitrogen sources being tested (data not shown). Therefore, we assume that the simultaneous utilization of conventional nutrients and SLES enables the cells to overcome the inhibition effect of growth. The same explanation could be given for nitrogen sources since they were advantageous in the case of A-K co-culture whereas they were disadvantageous in the S-A co-culture. In conclusion, the data presented here represent the first report about the capability of SLES degradation by these two co-cultures that were isolated from a wastewater plant. This could be a unique bacterial consortia in the degradation of high concentrations of SLES.
Literature Cited
Swisher RD (1987) Surfactant biodegradation, 2nd ed. Marcel Dekker, New York, NY
Jerábková H, Králová B, Náhlik J (1999). Biofilm of Pseudomonas C12B on glass support as catalytic agent for continuous SDS removal. Int Biodeg Biodeter 44:233–241
Brandt KK, Hesselsøe MP, Roslev K, Henriksen, Sørensen J (2001) Toxic Effects of linear alkylbenzene sulfonate on metabolic activity, growth rate, and microcolony formation of Nitrosomonas and Nitrosospira Strains. Appl Environ Microbiol 67:2489–2498
Jiménez L, Breen A, Thomas N, Federle TW, Sayler GS. (1991) Mineralization of linear alkylbenzene sulfonate by a four-member aerobic bacterial consortium. Appl Environ Microbiol 57:1566–1569
Rapaport RA, Eckhoff WS (1990) Monitoring linear alkylbenzene sulfonate in the environment: 1973–1986. Environ Toxicol Chem 9:1245–1257
Karsa DR (1987) Industrial applications of surfactants. The proceedings of a symposium organized by the northwest region of the royal society of chemistry. London, England: The Royal of Chemistry, Burlinton House
Jakobi G, Löhr A (1987) Detergent ingredients. In: Gerhartz W (ed) Ullmannsencyclopedia of industrial chemistry, 5 ed. VCH, Weinheim, pp 338–372
Schleheck D, Lechner M, Sch?nenberger R, Suter MJF, Cook AM (2003) Desulfonation and degradation of sulfodiphenylethercarboxylates from linear alkyldiphenyletherdisulfonate surfactants. Appl Environ Microbiol 69:938–944
Al-Shamailah W (2005) Effect of wastewater treatment on the bacterial quality and quantity at Mutah University Plant. MSc Thesis, Mutah University, Jordan
Miller JH (1972). Experiment in molecular genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York
Khleifat K (2006) Biodegradation of linear alkylbenzene sulfonate by a two-member facultative anaerobic bacterial consortium. Enz Microbial Technol 39(5):1030–1035
Longwell J, Maniece WD (1955). Determination of anionic detergent in sewage effluents and river water. Analyst 80:167–171
Li Z, Rosen M (1981). Two-phase mixed indicator titration method for determination of anionic surfactants. Anal Chem J 53(9):1516–1519
Loh KC, Wang SJ (1998) Enhancement of biodegradation of phenol and a nongrowth substrate 4-chlorophenol by medium augmentation with conventional carbon sources. Biodegradation 8:329–338
Khosravi M, Ryan W, Webster DA, Stork BC (1990a). Variation of oxygen requirement with plasmid size in recombinant Escherichia coli. Plasmid 23:138–143
Fendinger NJ, Versteeg DJ, Weeg E, Dyer SD, Rapaport RA (1994). Environmental behaviour and fate of anionic surfactants. In Baker LA (ed) Environmental Chemistry of Lakes and Reservoirs, Advances in Chemistry Series No. 237. American Chemical Society, Washington, DC. pp 527–557
Nuhoglu A, Yalcin B (2004). Modeling of phenol removal in a batch reactor, Process Biochem 40:1233–1239
Zhang CL, Valsaraj KT, Constant WD, Roy D (1999). Aerobic biodegradation kinetics of four anionic and nonionic surfactants at sub- and supra-critical micelle concentrations. Water Res 33(1):115–124
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Khleifat, K.M. Biodegradation of Sodium Lauryl Ether Sulfate (SLES) by Two Different Bacterial Consortia. Curr Microbiol 53, 444–448 (2006). https://doi.org/10.1007/s00284-006-0266-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00284-006-0266-4