
Abstract
Purpose
Carbonic anhydrase IX (CAIX) is a hypoxia-inducible enzyme with extracellular catalytic domain that is overexpressed in a variety of cancers including breast cancer and plays a crucial role in maintaining favourable intracellular pH and reducing extracellular pH. The purpose of the current study was to elucidate the prognostic significance of CAIX in the intrinsic subtypes of breast cancer and to characterise CAIX as a drug target in breast cancer.
Methods
The prognostic significance of CAIX mRNA expression was interrogated in a cohort of 3,455 breast tumours by using an online tool, Kaplan–Meier plotter. The functional effects of stable CAIX depletion by shRNA in three breast cancer cell lines—MDA-MB-231, MCF7 and SKBR-3, representing basal-like, luminal A and HER2+ subtypes, respectively—were studied by proliferation, invasion, clonal spheroid formation and chemosensitivity assays under normoxia and hypoxia. Finally, the effect of pharmacological CA inhibition alone or in the combination with doxorubicin on self-renewal was assessed by spheroid-forming assay.
Results
High CAIX mRNA expression was significantly associated with poor survival in patients with basal-like, luminal B and triple-negative breast cancer, but not luminal A and HER+ subtypes. Silencing of CAIX expression had no significant effect on the cell proliferation or viability upon treatment with doxorubicin in any of the cell lines studied, while it inhibited spheroid formation in hypoxic conditions. Furthermore, pharmacological inhibition of CAs using acetazolamide had a synergistic effect with doxorubicin on decreasing the spheroid-forming efficiency in MDA-MB-231 cells.
Conclusions
Inhibition of CAIX reduces the self-renewal capacity of breast cancer cells, and the combination of doxorubicin and CAIX inhibition is an attractive therapeutic strategy in basal-like and triple-negative breast cancer, which warrants further investigations.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Hypoxia and extracellular acidosis are common features of the vast majority of solid cancers that not only provide selective advantages for cancer cells, which are better adapted for survival in such conditions, but also promote cancer progression. Hypoxia signalling has been found to enhance cancer cell survival, chemoresistance, motility, tumour angiogenesis as well as self-renewal capacity and proliferation of putative cancer stem cells [1–3]. Extracellular acidosis arises by switching metabolism to glycolysis resulting in increased production and excretion of acidic metabolites, such as lactic and carbonic acids. Extracellular pH (pHe) in tumour tissues can be as low as 5.9 with a mean around 6.8, whereas in normal tissues under physiological conditions, it is around 7.2–7.4 [4, 5]. Acidic pHe of peritumoural tissues has been found to drive local invasion [6] and to promote the expression of stem cell markers, self-renewal and growth of glioma [7].
Carbonic anhydrase IX (CAIX) is a hypoxia-inducible enzyme with extracellular catalytic domain that plays a crucial role in the maintaining favourable (neutral to alkaline) intracellular pH (pHi) and reducing pHe [8]. CAIX catalyses are the reversible hydration of carbon dioxide to bicarbonate and protons in the extracellular space. Bicarbonate ions are transported into the cytoplasm by Cl−/HCO3 − exchangers and Na+/HCO3 − co-transporters resulting in the alkalinisation of pHi, while the proton remains in the extracellular space [8, 9].
According to Oncomine data, CAIX expression in normal tissues is limited to the intestinal and stomach mucosa, gallbladder and testis (www.oncomine.org; Shyamsundar and Roth Normal tissue datasets), while it is overexpressed in a wide variety of cancers, including breast cancer (BC) [8, 9]. Essentially, transcription of the CA9 gene is induced by binding of HIF-1α to the hypoxia-responsive element in the core promoter of CA9 [10]. However, at least in breast cancer, the expression of CAIX was found to be dependent on the interaction between HIF-1α and Notch3 intracellular domain, thus suggesting that Notch3 signalling may serve as a link between the regulation of hypoxia response and the maintenance of stem cell phenotype [11]. Furthermore, CAIX expression in cancers can be induced by the hypomethylation of the CA9 promoter or by inactivating mutations in the VHL gene that may drive the overexpression of CAIX independently of hypoxic conditions [12–14]. Most of the studies show that its overexpression in BC is a poor prognostic marker for distant metastasis and survival [15–17]; however, several other studies did not find a significant association with relapse-free or overall survival [18, 19]. It seems likely that these contradictory findings at least partially may be explained by its differential expression in various subtypes of BC [16, 20]; however, the prognostic significance of CAIX expression within the subtypes has not been systematically studied so far.
CAIX has recently emerged as a very attractive therapeutic target due to the following features: (1) CAIX shows tissue-restricted pattern and low expression level in normal tissues while it is induced in a broad range of cancers [8, 9]; (2) it is located on the surface of cells [21, 22]; (3) CAIX-knockout mice show only a mild phenotype related to gastric pit cell hyperplasia and glandular atrophy [23]; and (4) its overexpression in cancers confers a survival advantage in hypoxia and facilitates cell migration and metastasis [16, 24–26]. Depletion of CAIX in cancer cell lines has been shown to result in attenuated growth of tumour xenografts and inhibition of spontaneous metastasis formation [16, 26]. Interestingly, CAIX was found to contribute to cell migration and invasion by facilitating formation of focal adhesions, ion transport and pH control at the leading edge of lamellipodia of moving cells [27, 28]. Currently, substantial effort is devoted to the development of potent, cancer-selective CAIX inhibitors, including monoclonal antibodies [29] and small molecule inhibitors [30–32].
In the current study, we assessed the prognostic and predictive relevance of CAIX mRNA expression in intrinsic subtypes of BC by a meta-analysis of gene expression profiles from 3,455 breast tumours using Kaplan–Meier plotter [33] and investigated the biological effects of genetic silencing and pharmacological inhibition of CAIX in three breast cancer cell lines representing basal-like, luminal A and HER2+ subtypes.
Materials and methods
Reagents
Doxorubicin was purchased from MP Biomedicals (Carlsbad, CA, USA), and acetazolamide was purchased from Sigma-Aldrich (St. Louis, MO, USA). Both reagents were dissolved in dimethyl sulfoxide (DMSO) and the final concentration of DMSO in the medium never exceeded 0.1 %.
Meta-analysis by Kaplan–Meier Plotter
The prognostic significance of CAIX mRNA expression levels in breast cancer subtypes was evaluated using an online tool—Kaplan–Meier Plotter (http://kmplot.com/analysis/) that integrates gene expression and clinical data on 3,455 breast cancer patients. Relapse-free survival (RFS) and overall survival (OS) were selected as survival endpoints. For CAIX expression, Affymetrix ID 205199_at probe set was used. The probe set quality was estimated as intermediate—i.e. average expression between 100 and 500 expression units. Patient samples were split into two groups according to upper quartile of CAIX expression level. This cut-off was selected as it more closely matches to the overexpression rates observed in immunohistochemical studies [15, 16, 19]. The patient groups were compared by Kaplan–Meier survival plots, and the hazard ratio (HR; 95 % confidence intervals) and log-rank P values were calculated. Biased arrays (i.e. those with two or more parameters out of the 95 % range of all arrays) were excluded from the analysis. No cut-offs were applied for the duration of the patient follow-up.
Cell culture, stable transfection and exposure to hypoxia
The human breast cancer cell lines MDA-MD-231, MCF7 and SKBR-3 were purchased from European Collection of Cell Cultures (ECACC, UK) and maintained as recommended by the manufacturer. To generate stable CAIX-silenced cells, all three cell lines were transfected with a pool of three plasmids each encoding CAIX-specific shRNA (1 µg) and a negative control (NC) plasmids encoding scrambled shRNAs (1 µg) (Santa Cruz Biotechnology, TX, USA) using shRNA transfection medium and shRNA transfection reagent (Santa Cruz Biotechnology, USA) according to the manufacturers’ protocol. For selection of stably transfected cells, puromycin dihydrochloride (Santa Cruz Biotechnology, USA) was added to medium 48 h post-transfection (6 µg/ml for MCF7, 2 µg/ml for MDA-MB-231 and 1 µg/ml for SKBR-3 cells). The efficiency of CAIX silencing was assessed by qRT-PCR and immunocytochemistry. The obtained stable transfectants with depleted CAIX expression were designated as MDA-MD-231-shCAIX, MCF7-shCAIX and SKBR-3-shCAIX and the respective negative controls as MDA-MD-231-shNC, MCF7-shNC and SKBR-3-shNC. The cells were either incubated at 37 °C with 5 % CO2 in a humidified incubator (normoxia) or exposed to hypoxia at 1 % oxygen, 94 % N2 and 5 % CO2 at 37 °C in a tightly sealed chamber (Billupus-Rothemberg Inc., USA).
For the growth curve analysis, the cells were seeded at a density 0.5 × 104 cells per well in 24-well plates and cultured for 7 days under hypoxia or normoxia. Triplicate wells were trypsinised and the cells were counted using hemocytometer every 24 h.
RNA isolation and quantitative RT-PCR
Total RNA was isolated using TRI reagent (Sigma-Aldrich, USA) according to the manufacturers’ protocol. RNA was treated with DNAse I (Ambion, Life Technologies, USA) prior to cDNA synthesis and quantified by NanoDrop ND-100 spectrophotometer. cDNA was synthesised by random hexamer priming from 2 µg of total RNA by using RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, MA, USA) according to the manufacturers’ instructions. Quantitative RT-PCRs (qRT-PCR) were performed using 2 µl of 1:10 diluted cDNA reaction mixtures, ABSolute Blue™ SYBR green Low ROX (Thermo Fisher Scientific, USA), and ViiA 7 real-time PCR system (Applied Biosystems, Life Technologies, USA). The primer sequences used for CAIX amplification were as follows: 5′-GAAGAAATCGCTGAGGAAGG-3′ (forward) and 5′-CTGTCTGGTTAAACACAGTCC-3′ (reverse). To normalise the expression data, normalisation factor was calculated for each cDNA from the expression values of three most stable reference genes (ACTB, PGK1, and YWHAZ; the primer sequences are available upon request) selected among seven frequently used housekeeping genes by using geNorm software.
Immunofluorescence analysis
Spheroids were deposited on poly-l-lysine-coated slides by centrifuging at 800 rpm for 5 min using a Shandon cytospin, fixed in ice cold methanol/acetone (1:1) for 20 min, washed in PBS and blocked with 3 % BSA in PBS for 30 min. Slides were incubated with rabbit anti-CAIX primary antibody (clone H120, Santa Cruz Biotechnology, USA) (1:50) in PBS, 1 % BSA 14 h at +4 °C, washed in PBS 3 × 5 min, and incubated with goat anti-rabbit secondary antibody conjugated with FITC (Jacksons Immunoresearch, USA) at a dilution 1:100 in PBS, 1 % BSA for 1 h at room temperature. Slides were mounted with Prolong Gold antifade reagent with DAPI (Invitrogen, Life Technologies, USA). Images were captured using Leica TCP SP2 SE confocal microscope (Leica Microsystems, Germany) using the same exposure time to capture images.
Cytotoxicity assay
The effect of CAIX silencing on sensitivity to doxorubicin was assessed by MTT (3-[4,5–dimethylthiazol–2–yl]-2,5-diphenyltetrazolium bromide) assay (Cayman Chemicals, Ann Arbor, MI, USA). MDA-MD-231-shCAIX, MCF7-shCAIX and SKBR-3-shCAIX and the respective negative control cells were seeded at density 5 × 103 cells per well in 96-well plates with doxorubicin at six concentrations ranging from 0.05 to 50 µM or DMSO as the solvent control in triplicates. After 72 h of incubation under hypoxia or normoxia, MTT reagent was added to a final concentration of 1 mM and incubated for 4 h. The formazan crystals were solubilised using DMSO, and the concentration was determined by measuring optical density at 540 nm using Anthos HTII plate reader. Half-maximal inhibitory concentrations (IC50 values) were calculated using Graph Pad Prism 5 software (GraphPad Software, Inc, USA).
Spheroid-forming assay
For spheroid cultures, single-cell suspensions of MDA-MD-231-shCAIX, MCF7-shCAIX and SKBR-3-shCAIX and the respective negative control cells were plated at density 2 × 103 cells per ml in 96-well ultra-low attachment plates (Corning, NY, USA) in serum-free DMEM/F12 medium containing 1X B-27 supplement Minus AO (Life Technologies, Carlsbad, CA, USA), EGF (20 ng/ml, R&D Systems, MN, USA), bFGF (10 ng/ml, R&D Systems, MN, USA) and 1 % methylcellulose (Sigma-Aldrich, St. Louis, MO, USA). After 7 days in culture, the tumour spheroids containing at least 16 cells were counted under an inverted microscope. For subsequent passages, the spheroids were dissociated enzymatically with 1× trypsin and mechanically by passing through a 21G needle and re-plated at density 2 × 103 cells per ml.
To evaluate the effects of drugs, the cells were treated with doxorubicin and/or acetazolamide at various concentrations or with DMSO as the solvent control in six replicate wells and the spheroids were counted as described above. IC50 values for spheroid-forming ability were calculated using Graph Pad Prism 5 software (GraphPad Software, Inc, USA). The drug interaction was evaluated by determining the combination index (CI) using the CompuSyn software (ComboSyn, Inc. Paramus, NJ, USA).
Invasion assay
The invasive potential of MDA-MD-231-shCAIX, MCF7-shCAIX and SKBR-3-shCAIX and the respective negative control cells was assessed using fluorimetric QCM ECMatrix Cell Invasion Assay with ECM-coated inserts, 8 μm pore size (Merck Millipore, USA) according to the manufacturers’ instructions. The lower chamber was filled with DMEM/F12 with 10 % FBS. Cells were seeded in the upper chamber at density of 0.5 × 106 cells/ml in serum-free medium and maintained in hypoxia for 48 h. The cells that migrated through the ECM layer were dissociated from the membrane, subsequently lysed and detected by CyQuant GR dye using Victor 3 V 1420 Multilabel Counter (Perkin Elmer, CA, USA).
Statistical analysis
Sphere-forming efficiency was calculated as the number of spheroids formed per 1,000 single cells seeded. All experiments in sphere-forming assays were performed twice with six replicate wells. The concentration of drugs that inhibited the sphere-forming efficiency by 50 % (IC50) was calculated using GraphPad Prism software version 5 (GraphPad Software, Inc, USA). All qRT-PCR experiments were performed in duplicates, and the data are represented in graphs as means ± SD. All experiments in growth curve, cytotoxicity and invasion assays were performed in triplicates. The data are represented as means ± SD. Statistical significance was determined by the nonparametric Mann–Whitney U test, and differences were considered to be significant at P < 0.05.
Results
High CAIX mRNA expression is associated with poor survival in basal-like, luminal B and triple-negative but not luminal A and HER2+ breast cancer
In order to evaluate the prognostic relevance of CAIX mRNA expression in patients with different breast cancer subtypes, we exploited an online tool, Kaplan–Meier plotter, interrogating microarray-based gene expression and survival data in a large cohort of breast cancer patients (n = 3,455). Tumour samples were classified into the following subtypes according to their gene expression signature: basal-like, luminal A, luminal B, HER2+ and triple-negative breast cancer (TNBC) that was defined as a subset of basal-like cancers negative for the progesterone receptor expression (Table 1) [33]. Kaplan–Meier analyses showed that high CAIX mRNA expression was significantly associated with shorter RFS in patients with the basal subtype (HR = 1.65, P = 3.5 × 10−4) and TNBC (HR = 2.54, P = 1.7 × 10−4), while no association with RFS was found in luminal A and B subtypes, and HER2+ subtype even showed an inverse correlation, yet it did not reach a statistical significance (Fig. 1; Table 1). However, when OS was used as the endpoint, only luminal B subtype showed an association between high CAIX expression and worse OS (HR = 1.85, P = 0.0058). The same tendency was present in basal-like cancers; however, it did not achieve statistical significance, whereas CAIX expression was not associated with OS in luminal A and HER2+ subtypes.

Kaplan–Meier plots showing the association between CAIX mRNA expression and relapse-free survival or overall survival in various subtypes of breast cancer. a–e Kaplan–Meier relapse-free survival (RFS) curves stratifying patients with different breast cancer subtypes by CAIX expression; f Kaplan–Meier overall survival (OS) curve stratifying luminal B breast cancer patients by CAIX expression. Black lines patients with low CAIX mRNA levels, red patients with high CAIX mRNA levels (cut-off: upper quartile of CAIX expression level); vertical lines censored events
When the cases were further subdivided according to the treatment they received (chemotherapy vs endocrine therapy vs systemically untreated patients), high CAIX expression was significantly associated with shorter RFS in patients with basal-like BC who were systemically treated with chemotherapy (HR = 2.2, P = 0.014), but not in patients without systemic treatment, luminal A or B subtypes receiving endocrine therapy or HER2+ subtype receiving chemotherapy. On the contrary, in TNBC cases, stronger association between CAIX expression and RFS was observed in systemically untreated patients than in chemotherapy-treated patients (Table 1).
Genetic silencing of CAIX in breast cancer cells reduces spheroid-forming ability and invasiveness
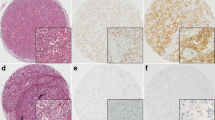
To interrogate the functional role of CAIX in breast cancer, three cell lines—MDA-MD-231, MCF7 and SKBR-3 cells representing basal-like, luminal A and HER2+ subtypes, respectively— were selected for the study. All cell lines expressed low levels of CAIX mRNA under normoxia, while it was strongly upregulated under hypoxia (Fig. 2a). The cell lines were stably transfected with a pool of three plasmids encoding CAIX-specific shRNAs or with a plasmid encoding scrambled shRNAs. qRT-PCR analysis demonstrated that CAIX mRNA expression was decreased by 98 and 81 % in MDA-MD-231 cells, by 93 and 86 in MCF7 cells and 90 and 84 % in SKBR-3 cells under normoxia and hypoxia, respectively (Fig. 2b). Figure 2c shows that hypoxia-induced CAIX protein expression was efficiently decreased in CAIX-shRNA-transfected cells compared with NC-shRNA-transfected cells grown as spheroids.

Effects of CAIX silencing by shRNA in breast cancer cells. a qRT-PCR analysis of CAIX mRNA expression in MDA-MD-231, MCF7 and SKBR-3 cells under normoxia and hypoxia. b qRT-PCR analysis of CAIX mRNA silencing by shRNA in MDA-MD-231, MCF7 and SKBR-3 cells. c Immunofluorescence analysis of CAIX silencing in MDA-MD-231, MCF7 and SKBR-3 cells cultured as spheroids in serum-free medium under hypoxia (scale bar 37.6 µm). d Growth curves for MDA-MD-231-shCAIX, MCF7-shCAIX and SKBR-3-shCAIX and the respective negative control cells under hypoxia. e Invasion assay: the cells that migrated through the ECM-coated membrane with 8 μm pore size in the transwell invasion assay were dissociated, lysed and quantified by staining with CyQuant GR dye. Each bar represents the mean from three replicates ± SD. f Spheroid formation assay: CAIX-shRNA- and NC-shRNA-transfected cells were grown as spheroids under normoxia or hypoxia for 7 days, and multicellular spheroids containing at least 16 cells were counted and expressed as a number of spheroids per 1,000 single cells plated. Each bar represents the mean from six replicates ± SD. g Passaging of spheroids: primary spheroids were dissociated and re-plated at density 2 × 103 cells/ml; the secondary and tertiary spheroids were counted after 7 days
The growth curve analysis showed that CAIX silencing did not affect the proliferation rate and/or cell death rate in all three CAIX-shRNA-transfected cell lines compared with the control cells grown in a monolayer under hypoxia (Fig. 2d) or normoxia (not shown).
The effect of CAIX silencing on the invasive potential under hypoxia was assessed by the transwell invasion assay through ECM-coated membranes. The number of invasive cells was decreased by 21.5 % in the MDA-MD-231-shCAIX cells compared with MDA-MD-231-shNC cells (P = 0.03), while no significant differences were observed between MCF7-shCAIX, SKBR-3-shCAIX and the respective control cells (Fig. 2e).
To assess the spheroid-forming ability, the cells were plated at low density in methylcellulose-containing serum-free medium in ultra-low attachment plates. In these conditions, cells grew as non-adherent, 3D multicellular spheroids. Clonal origin of the spheroids was confirmed by co-culture of DiD-labelled and DiO-labelled cells (data not shown). This assay showed that MDA-MD-231-shCAIX and SKBR-3-shCAIX cells had significantly decreased number of spheroid-forming cells compared with the control cells both under normoxia and hypoxia, while the spheroid-forming ability of MCF7-shCAIX cells was decreased only in hypoxia (Fig. 2f). Moreover, secondary and tertiary spheroid formation was significantly decreased in MDA-MD-231-shCAIX cells, and they were not able to survive beyond third passage, indicating their reduced self-renewal capacity in vitro, while it was only slightly lower in MCF7-shCAIX cells (Fig. 2g).
Genetic silencing and pharmacological inhibition of CAIX synergises with doxorubicin to reduce the spheroid-forming ability
To examine the effect of CAIX silencing on the survival of cells upon treatment with a conventional chemotherapeutic drug—doxorubicin—the viability of MDA-MD-231-shCAIX, MCF7-shCAIX and the respective control cells was tested by MTT assay in the presence of 0.05–50 µM doxorubicin. The dose–response curves were generated under normoxia and hypoxia, and the doxorubicin concentration that inhibited their growth by 50 % (IC50) was calculated (Table 2). This demonstrated that CAIX depletion did not have a significant effect on the growth inhibition by doxorubicin in both cell lines under hypoxic and normoxic conditions.
Given that CAIX depletion did not affect the cell proliferation, while it reduced the spheroid-forming ability, thus suggesting that it specifically affects a cell population with the stem cell properties, we next examined how the combination of CAIX depletion and treatment with doxorubicin affects the sphere-forming efficiency under hypoxia (Fig. 3a). In MDA-MD-231 cells, silencing of CAIX remarkably reduced the number of sphere-forming cells in the presence of doxorubicin under hypoxia (by fivefold at 10nM doxorubicin and sixfold at 100nM doxorubicin, P < 0.005). The same tendency was observed in MCF7-shCAIX and SKBR-3-shCAIX cells; however, the effect was less prominent (<2-fold).

Effects of genetic silencing and pharmacological inhibition of CAIX on sensitivity of breast cancer cells to doxorubicin. a Spheroid-forming ability of MDA-MD-231-shCAIX, MCF7-shCAIX and SKBR-3-shCAIX and the respective control cells treated with doxorubicin (DOX) under hypoxia for 7 days. The spheroid-forming efficacy is calculated relatively to DMSO control for each cell line. b Spheroid-forming ability of MDA-MD-231 cells under co-treatment with doxorubicin and acetazolamide (AZM) under hypoxia for 7 days. The spheroid-forming efficacy is calculated relatively to DMSO-treated cells. c Calculation of CI values for the DOX and AZM combinations demonstrates synergistic interaction for reducing spheroid-forming ability. CI < 1—synergism, CI = 1—additive effect and CI > 1—antagonism
Finally, we examined whether the same effect can be achieved via pharmacological inhibition of carbonic anhydrases. The MDA-MD-231 cells were treated with doxorubicin and a non-selective CA inhibitor acetazolamide either alone or in the combination. Co-treatment with doxorubicin (10, 50, 100 nM) and acetazolamide (100, 500, 1,000 nM) inhibited spheroid formation more efficiently than either drug alone (Fig. 3b). In the presence of 100 nM acetazolamide, IC50 of doxorubicin for the spheroid formation decreased from 19.82 to 6.57, and in the presence of 500 nM acetazolamide—to 3.78. To evaluate the mode of drug interaction, the combination index (CI) was calculated for each of the drug combination using the CompuSyn software (Fig. 3c). A CI ranged from 0.21 to 0.87 for various dose combinations indicating strong to moderate synergism [34, 35].
Discussion
Several previous immunohistochemical studies have demonstrated that high CAIX expression level is significantly associated with poor survival in breast cancer patients [15–17, 20, 36]. In the current study, we used an online tool, Kaplan–Meier Plotter [33], to evaluate the prognostic and predictive significance of CAIX mRNA expression in various intrinsic subtypes of BC and showed that high CAIX mRNA expression is significantly associated with worse RFS in the basal-like and triple-negative BC and with worse OS in luminal B subtype, while no association was found in the luminal A and HER2+ subtypes. In the basal-like BC cases, the association was stronger in chemotherapy-treated patients than in a cohort of systemically untreated patients suggesting that CAIX is a predictive rather than prognostic biomarker in the basal-like BC and the measurement of CAIX mRNA level could be used to predict effectiveness of chemotherapy. This is in agreement with previous IHC studies showing that high CAIX expression correlates with chemoresistance in basal-like BC [20] and serves as a predictive marker of doxorubicin resistance [37]. Whereas in TNBC, CAIX mRNA expression appeared to be a truly prognostic marker as it shows stronger correlation with RFS in systemically untreated patients than in chemotherapy-treated patients. However, the sample size in these cohorts is relatively small that precludes drawing conclusions about prognostic versus predictive relevance of CAIX in TNBC and potentially different roles of CAIX in basal-like BC and TNBC.
The lack of association with the survival in luminal A and HER2+ subtypes at least partially may be due to lower rate of CAIX overexpression, distinct activation mechanisms and biological roles of CAIX in these subtypes. Although in this study, we could not carry out a statistical analysis of the CAIX expression levels in various subtypes, we observed that the median expression values tended to be lower in luminal A and HER2+ subtypes compared with basal-like BC and TNBC. This is in line with the studies by Lou et al. [16] reporting that 51 % of basal breast cancers are CAIX positive by IHC, while only 8 % of luminal A, 11 % luminal B and 33 % of HER2+ cancers are CAIX positive, and by Tan et al. [20] showing that basal-like tumours were nine times more likely to be associated with CAIX expression and that patients with CAIX-positive tumours had significantly worse prognosis. On the other hand, increasing evidence suggest that the response to hypoxia and activation of HIF-dependent gene networks is particularly robust in TNBC and may represent a distinctive molecular feature of TNBC [38]. In TNBC, HIF-1α has been shown to be activated not only by the conventional pathway —local intratumoural hypoxia— but also by downregulation of SHARP1 [39] and by unfolded protein response mediated by XBP1 [40]. Interestingly, silencing of SHARP1 in TNBC cells has been shown to result in the increase in CAIX expression, and SHARP1 expression level in TNBC tissues was shown to inversely correlate with the number of CAIX-positive cells [39]. This shows that the mechanisms leading to the CAIX overexpression can be distinct in TNBC compared with other subtypes of breast cancer; however, whether or not CAIX by itself plays distinct functional roles in various subtypes remains to be determined.
Furthermore, the differences in the prognostic and predictive significance of CAIX in various BC subtypes may also arise from different patterns of cell types that produce CAIX. Although in tumours, CAIX is expressed predominantly by cancer cells, it has been found in adjacent stroma too [41]. Moreover, the proportion of CAIX-positive stroma cells has been found to differ among BC subtypes [42]. Interestingly, CAIX expression by cancer-associated fibroblasts has been found to be a better predictor of outcome than its expression by cancer cells in patients with lung cancer [43], while in prostate cancer, it has been found to drive epithelial-mesenchymal transition in cancer cells [44].
CAIX has been previously proposed to serve as a therapeutic target in breast [16, 45] and renal cancer [46–48]. Depletion of CAIX expression in mouse and human breast cancer cells has been found to attenuate tumour growth and inhibited formation of spontaneous lung metastasis in mouse models [16]. However, at least in renal cancer, clinical trials with a monoclonal antibody G250 that binds to CAIX so far have shown only moderate success [47]. Here, we show that depletion of CAIX expression did not affect the proliferation rate and survival of breast cancer cells, while it significantly reduced the spheroid-forming efficiency under hypoxia. Formation of clonal spheroids is a characteristic of breast cancer stem cells (CSCs) and correlates with the tumourigenicity and therefore has been used as a measure of self-renewal capacity [49, 50]. Hence, these results suggest that inhibition of CAIX is unlikely to result in a rapid regression of a tumour mass, whereas it may play a role in regulating cancer stem cell population and prevent the emergence of cells with CSC phenotype in the hypoxic niche.
Furthermore, the genetic silencing of CAIX expression significantly decreased the spheroid-forming capacity of BC cells upon treatment with doxorubicin. This effect was more prominent in the MDA-MD-231 cells compared with MCF7 and SKBR3 cells. At least partially, this difference could be attributed to the distinct expression pattern of the CA family members in these cells. CAIX has been shown to be the only membrane-associated CA in the MDA-MD-231 cells, while MCF cells also express CAXII that may compensate the effects of CAIX depletion [51]. This suggests that in some cancers, inhibiting CAIX alone would not be sufficient to achieve a therapeutic benefit and that this would require the development of dual CAIX and CAXII inhibitors. Finally, we showed that the pharmacological inhibition of CAs by a non-selective CA inhibitor acetazolamide has a synergistic effect with doxorubicin on reducing the number of spheroid-forming cells. The molecular mechanism underlying the synergistic drug interaction is not known. However, a recent study by Lock et al. [24] showed that CAIX is required for mTOR expression, thereby modulating mTORC1 signalling, which has been suggested to play a crucial role in the maintenance of cancer stem cell phenotype in colorectal cancer [52, 53]. Hence, it seems likely that the synergistic effect on inhibiting the spheroid-forming ability is achieved by simultaneous targeting of mTORC1 pathway by CAIX inhibition and eliciting DNA damage by doxorubicin.
Interestingly, high CAIX expression has been found to predict doxorubicin resistance in breast cancer patients [37]. Doxorubicin is a highly active anthracycline drug that is frequently used for the treatment of advanced BC, in particular TNBC; however, a major limitation for its use is severe toxicity, including cardiotoxicity and hepatotoxicity [54]. Taken together, our study suggests that combining CAIX inhibition with doxorubicin could be a promising therapeutic strategy for the treatment of TNBC patients with high CAIX expression. We speculate that this would increase the therapeutic efficacy and would allow decreasing the dosage of doxorubicin, thus allowing to avoid the toxic side effects.
References
Soeda A, Park M, Lee D, Mintz A, Androutsellis-Theotokis A, McKay RD, Engh J, Iwama T, Kunisada T, Kassam AB, Pollack IF, Park DM (2009) Hypoxia promotes expansion of the CD133-positive glioma stem cells through activation of HIF-1alpha. Oncogene 28:3949–3959
Sullivan R, Pare GC, Frederiksen LJ, Semenza GL, Graham CH (2008) Hypoxia-induced resistance to anticancer drugs is associated with decreased senescence and requires hypoxia-inducible factor-1 activity. Mol Cancer Ther 7:1961–1973
Heddleston JM, Li Z, Lathia JD, Bao S, Hjelmeland AB, Rich JN (2010) Hypoxia inducible factors in cancer stem cells. Br J Cancer 102:789–795
Stubbs M, McSheehy PM, Griffiths JR, Bashford CL (2000) Causes and consequences of tumour acidity and implications for treatment. Mol Med Today 6:15–19
Vaupel P, Kallinowski F, Okunieff P (1990) Blood flow, oxygen consumption and tissue oxygenation of human tumors. Adv Exp Med Biol 277:895–905
Estrella V, Chen T, Lloyd M, Wojtkowiak J, Cornnell HH, Ibrahim-Hashim A, Bailey K, Balagurunathan Y, Rothberg JM, Sloane BF, Johnson J, Gatenby RA, Gillies RJ (2013) Acidity generated by the tumor microenvironment drives local invasion. Cancer Res 73:1524–1535
Hjelmeland AB, Wu Q, Heddleston JM, Choudhary GS, MacSwords J, Lathia JD, McLendon R, Lindner D, Sloan A, Rich JN (2011) Acidic stress promotes a glioma stem cell phenotype. Cell Death Differ 18:829–840
McDonald PC, Winum JY, Supuran CT, Dedhar S (2012) Recent developments in targeting carbonic anhydrase IX for cancer therapeutics. Oncotarget 3:84–97
Chiche J, Ilc K, Brahimi-Horn MC, Pouyssegur J (2010) Membrane-bound carbonic anhydrases are key pH regulators controlling tumor growth and cell migration. Adv Enzyme Regul 50:20–33
Wykoff CC, Beasley NJ, Watson PH, Turner KJ, Pastorek J, Sibtain A, Wilson GD, Turley H, Talks KL, Maxwell PH, Pugh CW, Ratcliffe PJ, Harris AL (2000) Hypoxia-inducible expression of tumor-associated carbonic anhydrases. Cancer Res 60:7075–7083
Shareef MM, Udayakumar TS, Sinha VK, Saleem SM, Griggs WW (2013) Interaction of HIF-1alpha and Notch3 is required for the expression of carbonic anhydrase 9 in breast carcinoma cells. Genes Cancer 4:513–523
Sung HY, Ju W, Ahn JH (2014) DNA hypomethylation-mediated overexpression of carbonic anhydrase 9 induces an aggressive phenotype in ovarian cancer cells. Yonsei Med J 55:1656–1663
Nakamura J, Kitajima Y, Kai K, Hashiguchi K, Hiraki M, Noshiro H, Miyazaki K (2011) Expression of hypoxic marker CA IX is regulated by site-specific DNA methylation and is associated with the histology of gastric cancer. Am J Pathol 178:515–524
Wiesener MS, Munchenhagen PM, Berger I, Morgan NV, Roigas J, Schwiertz A, Jurgensen JS, Gruber G, Maxwell PH, Loning SA, Frei U, Maher ER, Grone HJ, Eckardt KU (2001) Constitutive activation of hypoxia-inducible genes related to overexpression of hypoxia-inducible factor-1alpha in clear cell renal carcinomas. Cancer Res 61:5215–5222
Hussain SA, Ganesan R, Reynolds G, Gross L, Stevens A, Pastorek J, Murray PG, Perunovic B, Anwar MS, Billingham L, James ND, Spooner D, Poole CJ, Rea DW, Palmer DH (2007) Hypoxia-regulated carbonic anhydrase IX expression is associated with poor survival in patients with invasive breast cancer. Br J Cancer 96:104–109
Lou Y, McDonald PC, Oloumi A, Chia S, Ostlund C, Ahmadi A, Kyle A, dem Auf KU, Leung S, Huntsman D, Clarke B, Sutherland BW, Waterhouse D, Bally M, Roskelley C, Overall CM, Minchinton A, Pacchiano F, Carta F, Scozzafava A, Touisni N, Winum JY, Supuran CT, Dedhar S (2011) Targeting tumor hypoxia: suppression of breast tumor growth and metastasis by novel carbonic anhydrase IX inhibitors. Cancer Res 71:3364–3376
Beketic-Oreskovic L, Ozretic P, Rabbani ZN, Jackson IL, Sarcevic B, Levanat S, Maric P, Babic I, Vujaskovic Z (2011) Prognostic significance of carbonic anhydrase IX (CA-IX), endoglin (CD105) and 8-hydroxy-2′-deoxyguanosine (8-OHdG) in breast cancer patients. Pathol Oncol Res 17:593–603
Span PN, Bussink J, Manders P, Beex LV, Sweep CG (2003) Carbonic anhydrase-9 expression levels and prognosis in human breast cancer: association with treatment outcome. Br J Cancer 89:271–276
Kyndi M, Sorensen FB, Knudsen H, Alsner J, Overgaard M, Nielsen HM, Overgaard J (2008) Carbonic anhydrase IX and response to postmastectomy radiotherapy in high-risk breast cancer: a subgroup analysis of the DBCG82 b and c trials. Breast Cancer Res 10:R24
Tan EY, Yan M, Campo L, Han C, Takano E, Turley H, Candiloro I, Pezzella F, Gatter KC, Millar EK, O’Toole SA, McNeil CM, Crea P, Segara D, Sutherland RL, Harris AL, Fox SB (2009) The key hypoxia regulated gene CAIX is upregulated in basal-like breast tumours and is associated with resistance to chemotherapy. Br J Cancer 100:405–411
Li Y, Tu C, Wang H, Silverman DN, Frost SC (2011) Catalysis and pH control by membrane-associated carbonic anhydrase IX in MDA-MB-231 breast cancer cells. J Biol Chem 286:15789–15796
Swietach P, Patiar S, Supuran CT, Harris AL, Vaughan-Jones RD (2009) The role of carbonic anhydrase 9 in regulating extracellular and intracellular ph in three-dimensional tumor cell growths. J Biol Chem 284:20299–20310
Leppilampi M, Karttunen TJ, Kivela J, Gut MO, Pastorekova S, Pastorek J, Parkkila S (2005) Gastric pit cell hyperplasia and glandular atrophy in carbonic anhydrase IX knockout mice: studies on two strains C57/BL6 and BALB/C. Transgenic Res 14:655–663
Lock FE, McDonald PC, Lou Y, Serrano I, Chafe SC, Ostlund C, Aparicio S, Winum JY, Supuran CT, Dedhar S (2013) Targeting carbonic anhydrase IX depletes breast cancer stem cells within the hypoxic niche. Oncogene 32:5210–5219
Pastorek J (2014) Pastorekova S. Hypoxia-induced carbonic anhydrase IX as a target for cancer therapy. From biology to clinical use, Semin Cancer Biol
McIntyre A, Patiar S, Wigfield S, Li JL, Ledaki I, Turley H, Leek R, Snell C, Gatter K, Sly WS, Vaughan-Jones RD, Swietach P, Harris AL (2012) Carbonic anhydrase IX promotes tumor growth and necrosis in vivo and inhibition enhances anti-VEGF therapy. Clin Cancer Res 18:3100–3111
Csaderova L, Debreova M, Radvak P, Stano M, Vrestiakova M, Kopacek J, Pastorekova S, Svastova E (2013) The effect of carbonic anhydrase IX on focal contacts during cell spreading and migration. Front Physiol 4:271
Svastova E, Witarski W, Csaderova L, Kosik I, Skvarkova L, Hulikova A, Zatovicova M, Barathova M, Kopacek J, Pastorek J, Pastorekova S (2012) Carbonic anhydrase IX interacts with bicarbonate transporters in lamellipodia and increases cell migration via its catalytic domain. J Biol Chem 287:3392–3402
Stillebroer AB, Boerman OC, Desar IM, Boers-Sonderen MJ, van Herpen CM, Langenhuijsen JF, Smith-Jones PM, Oosterwijk E, Oyen WJ, Mulders PF (2013) Phase 1 radioimmunotherapy study with lutetium 177-labeled anti-carbonic anhydrase IX monoclonal antibody girentuximab in patients with advanced renal cell carcinoma. Eur Urol 64:478–485
Monti SM, Supuran CT, De SG (2013) Anticancer carbonic anhydrase inhibitors: a patent review (2008–2013). Expert Opin Ther Pat 23:737–749
Rogez-Florent T, Meignan S, Foulon C, Six P, Gros A, Bal-Mahieu C, Supuran CT, Scozzafava A, Frederick R, Masereel B, Depreux P, Lansiaux A, Goossens JF, Gluszok S, Goossens L (2013) New selective carbonic anhydrase IX inhibitors: synthesis and pharmacological evaluation of diarylpyrazole-benzenesulfonamides. Bioorg Med Chem 21:1451–1464
Dudutiene V, Matuliene J, Smirnov A, Timm DD, Zubriene A, Baranauskiene L, Morkunaite V, Smirnoviene J, Michailoviene V, Juozapaitiene V, Mickeviciute A, Kazokaite J, Baksyte S, Kasiliauskaite A, Jachno J, Revuckiene J, Kisonaite M, Pilipuityte V, Ivanauskaite E, Milinaviciute G, Smirnovas V, Petrikaite V, Kairys V, Petrauskas V, Norvaisas P, Linge D, Gibieza P, Capkauskaite E, Zaksauskas A, Kazlauskas E, Manakova E, Grazulis S, Ladbury JE, Matulis D (2014) Discovery and characterization of novel selective inhibitors of carbonic anhydrase IX. J Med Chem
Gyorffy B, Lanczky A, Eklund AC, Denkert C, Budczies J, Li Q, Szallasi Z (2010) An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1,809 patients. Breast Cancer Res Treat 123:725–731
Chou TC (2010) Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res 70:440–446
Chou TC (2006) Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol Rev 58:621–681
Kaya AO, Gunel N, Benekli M, Akyurek N, Buyukberber S, Tatli H, Coskun U, Yildiz R, Yaman E, Ozturk B (2012) Hypoxia inducible factor-1 alpha and carbonic anhydrase IX overexpression are associated with poor survival in breast cancer patients. J BUON 17:663–668
Betof AS, Rabbani ZN, Hardee ME, Kim SJ, Broadwater G, Bentley RC, Snyder SA, Vujaskovic Z, Oosterwijk E, Harris LN, Horton JK, Dewhirst MW, Blackwell KL (2012) Carbonic anhydrase IX is a predictive marker of doxorubicin resistance in early-stage breast cancer independent of HER2 and TOP2A amplification. Br J Cancer 106:916–922
Bernardi R, Gianni L (2014) Hallmarks of triple negative breast cancer are emerging at last? Cell Res
Montagner M, Enzo E, Forcato M, Zanconato F, Parenti A, Rampazzo E, Basso G, Leo G, Rosato A, Bicciato S, Cordenonsi M, Piccolo S (2012) SHARP1 suppresses breast cancer metastasis by promoting degradation of hypoxia-inducible factors. Nature 487:380–384
Chen X, Iliopoulos D, Zhang Q, Tang Q, Greenblatt MB, Hatziapostolou M, Lim E, Tam WL, Ni M, Chen Y, Mai J, Shen H, Hu DZ, Adoro S, Hu B, Song M, Tan C, Landis MD, Ferrari M, Shin SJ, Brown M, Chang JC, Liu XS, Glimcher LH (2014) XBP1 promotes triple-negative breast cancer by controlling the HIF1alpha pathway. Nature 508:103–107
Bartosova M, Parkkila S, Pohlodek K, Karttunen TJ, Galbavy S, Mucha V, Harris AL, Pastorek J, Pastorekova S (2002) Expression of carbonic anhydrase IX in breast is associated with malignant tissues and is related to overexpression of c-erbB2. J Pathol 197:314–321
Choi J, Kim DH, Jung WH, Koo JS (2013) Metabolic interaction between cancer cells and stromal cells according to breast cancer molecular subtype. Breast Cancer Res 15:R78
Nakao M, Ishii G, Nagai K, Kawase A, Kenmotsu H, Kon-No H, Hishida T, Nishimura M, Yoshida J, Ochiai A (2009) Prognostic significance of carbonic anhydrase IX expression by cancer-associated fibroblasts in lung adenocarcinoma. Cancer 115:2732–2743
Fiaschi T, Giannoni E, Taddei ML, Cirri P, Marini A, Pintus G, Nativi C, Richichi B, Scozzafava A, Carta F, Torre E, Supuran CT, Chiarugi P (2013) Carbonic anhydrase IX from cancer-associated fibroblasts drives epithelial-mesenchymal transition in prostate carcinoma cells. Cell Cycle 12:1791–1801
Pacchiano F, Carta F, McDonald PC, Lou Y, Vullo D, Scozzafava A, Dedhar S, Supuran CT (2011) Ureido-substituted benzenesulfonamides potently inhibit carbonic anhydrase IX and show antimetastatic activity in a model of breast cancer metastasis. J Med Chem 54:1896–1902
Buller F, Steiner M, Frey K, Mircsof D, Scheuermann J, Kalisch M, Buhlmann P, Supuran CT, Neri D (2011) Selection of carbonic anhydrase IX inhibitors from one million DNA-encoded compounds. ACS Chem Biol 6:336–344
Bleumer I, Knuth A, Oosterwijk E, Hofmann R, Varga Z, Lamers C, Kruit W, Melchior S, Mala C, Ullrich S, De MP, Mulders PF, Beck J (2004) A phase II trial of chimeric monoclonal antibody G250 for advanced renal cell carcinoma patients. Br J Cancer 90:985–990
Bauer S, Oosterwijk-Wakka JC, Adrian N, Oosterwijk E, Fischer E, Wuest T, Stenner F, Perani A, Cohen L, Knuth A, Divgi C, Jager D, Scott AM, Ritter G, Old LJ, Renner C (2009) Targeted therapy of renal cell carcinoma: synergistic activity of cG250-TNF and IFNg. Int J Cancer 125:115–123
Fillmore CM, Kuperwasser C (2008) Human breast cancer cell lines contain stem-like cells that self-renew, give rise to phenotypically diverse progeny and survive chemotherapy. Breast Cancer Res 10:R25
Zhang M, Behbod F, Atkinson RL, Landis MD, Kittrell F, Edwards D, Medina D, Tsimelzon A, Hilsenbeck S, Green JE, Michalowska AM, Rosen JM (2008) Identification of tumor-initiating cells in a p53-null mouse model of breast cancer. Cancer Res 68:4674–4682
Li Y, Wang H, Oosterwijk E, Tu C, Shiverick KT, Silverman DN, Frost SC (2009) Expression and activity of carbonic anhydrase IX is associated with metabolic dysfunction in MDA-MB-231 breast cancer cells. Cancer Invest 27:613–623
Cai Z, Ke J, He X, Yuan R, Chen Y, Wu X, Wang L, Wang J, Lan P, Wu X (2014) Significance of mTOR signaling and its inhibitor against cancer stem-like cells in colorectal cancer. Ann Surg Oncol 21:179–188
Francipane MG, Lagasse E (2013) Selective targeting of human colon cancer stem-like cells by the mTOR inhibitor Torin-1. Oncotarget 4:1948–1962
Carvalho C, Santos RX, Cardoso S, Correia S, Oliveira PJ, Santos MS, Moreira PI (2009) Doxorubicin: the good, the bad and the ugly effect. Curr Med Chem 16:3267–3285
Acknowledgments
This study was supported by the Latvian National Research Programme BIOMEDICINE.
Conflict of interest
None.
Author information
Authors and Affiliations
Corresponding author
Additional information
Lāsma Ivanova and Elīna Zandberga have contributed equally to this work.
Rights and permissions
About this article

Cite this article
Ivanova, L., Zandberga, E., Siliņa, K. et al. Prognostic relevance of carbonic anhydrase IX expression is distinct in various subtypes of breast cancer and its silencing suppresses self-renewal capacity of breast cancer cells. Cancer Chemother Pharmacol 75, 235–246 (2015). https://doi.org/10.1007/s00280-014-2635-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00280-014-2635-1