
Abstract
The nonhematopoietic bone marrow (BM) microenvironment provides a functional niche for hematopoietic cell maintenance, recruitment, and differentiation. It consists of multiple cell types including vasculature, bone, adipose tissue, and fibroblast-like bone marrow stromal cells (BMSC), which can be summarized under the generic term niche cells. BMSC express Toll-like receptors (TLRs) and are capable to respond to TLR-agonists by changing their cytokine expression pattern in order to more efficiently support hematopoiesis. Here, we show that in addition to enhanced myeloid colony formation from human CD34+ cells, lipopolysaccharide (LPS) stimulation retains overall higher numbers of CD34+ cells in co-culture assays using BMSC, with eightfold more CD34+ cells that underwent up to three divisions as compared to non-stimulated assays. When subjected to cytokine-supplemented myeloid colony-forming unit (CFU) assays or transplanted into newborn RAG2−/− γc −/− mice, CD34+ cells from LPS-stimulated BMSC cultures give rise to the full spectrum of myeloid colonies and T and B cells, respectively, thus supporting maintenance of myeloid and lymphoid primed hematopoietic progenitor cells (HPCs) under inflammatory conditions. Collectively, we suggest that BMSC enhance hematopoiesis during inflammatory conditions to support the replenishment of innate immune effector cells and to prevent the exhaustion of the hematopoietic stem and progenitor cell (HSPC) pool.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Bone marrow mesenchymal stem or stromal cells (BMSC) are part of the bone marrow environment and recapitulate bone and marrow formation upon transplantation of in vitro cultured cells [1]. In-depth genetic studies have provided evidence that BMSC constitute a heterogenous group of cells that are in intimate contact with hematopoietic cells and express cytokines that promote HSC maintenance and differentiation [2–6]. In vitro, BMSC have been shown to increase both the expansion of myeloid as well as hematopoietic stem and progenitor cells (HSPCs) [7–11]. Furthermore, it has been shown that BMSC express functional Toll-like receptors which increase their extent to support proliferation and myeloid differentiation of HSPCs in response to lipopolysaccharide (LPS) [12]. We here hypothesized that in addition to the support of myeloid cell development, LPS stimulation of BMSC increases their capacity to maintain early progenitors with myeloid as well as T and B cell proliferation potential.
Material and methods
Myeloid colony-forming assays
Assays were performed as described [13]. Specifically, to asses colony-forming unit (CFU) supporting capacity of human BMSCs supernatants either from LPS-stimulated or unstimulated BMSCs as well as IMDM supplemented with 20 % FCS, 2 mM L-glutamine, and 50 μM 2-mercaptoethanol for negative and positive controls were mixed with methylcellulose (Methocult H4100, 2.6 %, StemCell Technologies, Vancouver, Canada) to yield a final concentration of 0.9 % methylcellulose. Factors were added in the following concentrations as indicated within the figure: huSCF (10 ng/ml), huFLT3L (10 ng/ml), huTPO (50 ng/ml), huIL-3 (20 ng/ml), huIL-6 (10 ng/ml), huIL-11 (10 ng/ml), huGM-CSF (50 ng/ml), human erythropoietin huEPO (4 U/ml) (complete cytokine mix), and LPS (10 μg/ml). To asses CFU capacity from freshly isolated human CD34+ cells in the presence or absence of LPS and human hematopoietic cells recovered from either LPS-stimulated or unstimulated CB CD34+ and BMSC co-cultures or from cytokine-supported cultures, cells were added at a density of 1500 and 2000 cells per milliliter, respectively, to a methylcellulose/IMDM (20 % FCS 2 mM L-glutamine, 50 μM 2-mercaptoethanol) premix including complete human cytokine supplementation as described above and LPS as indicated. For evaluation of CFU activity from lin−c-Kit+ WT mouse, bone marrow (BM) in the presence or absence of LPS 4 × 102 cells were plated in methylcellulose (Methocult M3231, StemCell Technologies) mixed with IMDM (30 % FCS, 2 mM L-glutamine, 50 μM 2-mercaptoethanol) with the following factors added: mIL-3 (10 ng/ml), hIL-6 (10 ng/ml), mSCF (10 ng/ml), mGM-CSF (10 ng/ml), mTPO (50 ng/ml), huEPO (2 U/ml) (all R&D systems), and LPS (10 μg/ml; InvivoGen) as indicated.
Newborn transplantation assay
BALB/c Rag2−/−γc −/− mice were maintained at the Institute for Research in Biomedicine animal facility and treated in accordance with guidelines of the Swiss Federal Veterinary Office. All institutional and national guidelines for the care and use of laboratory animals were followed. Experiments were approved by the Dipartimento della Sanità e Socialità, Ticino, Switzerland. Newborn Rag2−/−γc −/− mice were irradiated within a 4-h interval with 2 × 2 Gy from a Cesium 137 source (Biobeam 8000, STS GmbH, Braunschweig, Germany) at 3.75 Gy/min as previously described [14]. At 2 h post-irradiation, mice were transplanted with hematopoietic cells recovered from co-cultures in 25 μl PBS into the liver (i.h.) using a 30-gauge needle (Hamilton Bonaduz AG, Bonaduz, Switzerland). Mice were weaned at 3 weeks and sacrificed at 4 weeks of age. For FACS analysis of human cell transplanted Rag2−/−γc −/− mice, monoclonal antibodies biotinylated or conjugated against the following human antigens were used: CD3 (UCHT1), CD4 (13B8.2), CD8 (B9.11) (Immunotech/Beckman Coulter, Marseille, France), CD19 (HIB19), CD34 (581), IL-3Rα/CD123 (9F5) (BD Biosciences, San Diego, USA), and CD45 (HI30) (Caltag, Carlsbad, USA).
Results and discussion
Cultured human BMSCs respond to TLR4 agonist stimulation with the secretion of myelopoiesis supporting cytokines
We isolated primary human stromal cells from the BM by plastic adherence. Stromal cultures developed from proliferating mesenchymal precursors (CFU-Fs) fulfilled MSC defining properties according to current consensus [15, 16]. As previously published, human BMSC were found to express a spectrum of Toll-like receptors (TLRs) [12, 17–19] with most predominant expression of TLR4 (Fig. 1a). Stimulation of human BMSC with the TLR4 agonist LPS induced production of G-CSF and GM-CSF protein and significantly enhanced the secretion of M-CSF, IL-6, and IL-11 (Fig. 1b). The biological activity of secreted cytokines was demonstrated in myeloid colony-forming unit (CFU) assays using human cord blood (CB) CD34+ HSPCs [13]. Human BMSC constitutively expressed cytokines that supported the formation of myeloid colonies, while in contrast to a fully cytokine-supplemented CFU control, mixed or pure erythroid colonies were not detected (Fig. 1c). Myeloid CFU activity was about 2.5-fold higher when supernatants of LPS-stimulated BMSCs were used, while direct LPS addition to unstimulated BMSC supernatant or media alone did not change CFU activity, arguing against a direct effect of LPS on TLR4 expressing CD34+ HSPCs in this setting (Fig. 1c). It has previously been shown that TLR1/2 as well as TLR7/8 agonists instruct commitment of human hematopoietic stem cells to a myeloid cell fate [20, 21]. To further rule out a predominant direct effect of LPS on hematopoietic progenitor cells, we applied CFU assays using human CD34+ CB hematopoietic progenitor cells (Fig 1d). TLR4 stimulation in the absence of stroma did not grossly influence the frequency of colony formation from human progenitor cells and was not biased towards a specific progenitor.

Human BMSC responds to inflammatory conditions in vitro and in vivo. a Tlr1-10 mRNA expression in human BMSCs (black bars), peripheral blood dendritic cells (PB DCs, grey bars), and natural interferon-producing cells (PB IPCs, white bars). Expression levels are normalized against 18S RNA. n.d. not detectable within 35 cycles of amplification. Mean ± SEM of one representative out of three independent experiments is shown. Each experiment was performed with three different bone marrow donors and one buffy coat donor. b Cytokine levels in supernatants from unstimulated (white bars) and LPS-stimulated (10 μg/ml, 48 h, grey bars) human BMSC cultures. Mean ± SEM of supernatants from five experiments, each with different donor BMSC, is shown. n.d. not detectable within the sensitivity of the assay. Statistically significant differences are indicated (*p < 0.05; **p < 0.01). c CFU activity of 500 sorted human CB CD34+ cells per well in the presence of supernatants from unstimulated and LPS-stimulated BMSC. Controls included cultures containing a full cytokine mix (SCF, FLT3L, TPO, IL-3, IL-6, IL-11, GM-CSF, EPO), unstimulated BMSC supernatant with addition of LPS, and cultures with only LPS added. All cultures were supplemented with a minimal cytokine mix containing SCF, TPO, and FLT3L. Mean ± SEM of a representative out of three independent experiments, performed with three different bone marrow donors, and pooled CD34+ cells is shown. Statistically significant differences are indicated (**p < 0.01). d CFU activity of human cord blood CD34+ cells after 12 days of cytokine-supplemented culture in the absence or presence of LPS (10 μg/ml). One thousand five hundred human CD34+ cells were plated per well, and colonies formed were counted in triplicate wells. CFU-E CFU-erythrocyte. Four independent experiments with each CD34+ cord blood cells from different donors are shown
We thus conclude that LPS stimulation of primary human BMSC induces enhanced production and release of functional myeloid cell differentiation-supporting cytokines that are capable of stimulating a quantitative increase of CFU-GM/G/M from a given number of human hematopoietic stem and progenitor cells.
LPS stimulation of human BMSCs enhances hematopoietic progenitor cell maintenance in vitro
Given the increase in the secretion of early acting cytokines IL-11 and IL-6 and the constitutive secretion of SCF, we evaluated the capacity of human BMSCs to maintain immature CB CD34+ cells in co-cultures with or without addition of LPS. Within 12 days of co-culture, total hematopoietic cell numbers were augmented (2–3 population doublings), but the increase was most pronounced in BMSC-free cultures supplemented only with SCF, TPO, and FLT3L (5–6 population doublings) (Fig. 2a). No difference in total cell numbers was seen with respect to TLR4 activation in either BMSC co-cultures or cytokine-only cultures (Fig. 2a). However, addition of LPS to BMSC and CB CD34+ cell co-cultures led to an approximate twofold higher percentage of recovered CD34+ cells as compared to LPS-free co-cultures. No significant effect on the maintenance of CB CD34+ cells was observed upon LPS addition to cultures supplemented with SCF, TPO, and FLT3L cytokines only (Fig. 2b). Importantly, as measured by CFSE dilution, LPS-stimulated BMSCs retained up to eightfold more CD34+ cells in divisions 0–3 as compared to co-cultures without LPS addition. By contrast, 0–3-fold dividing CD34+ cells were hardly detectable in cultures supplemented with cytokines only, irrespective of LPS addition (Fig. 2c).

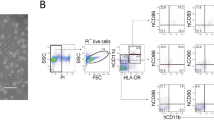
LPS-stimulated human BMSCs maintain human early hematopoietic progenitors with myeloid and lymphoid differentiation capacity. a Population doublings and b CD34+ cell percentages of sorted human CB CD34+ cells cultured for 12 days on human BMSCs or only in the presence of cytokines (SCF, TPO, FLT3L) without BMSCs. LPS (10 μg/ml, grey bars) was added from the beginning of cultures as indicated. Mean ± SD of three independent experiments with each different CD34+ cord blood cells and different BMSC donors is shown. c Proliferation of human CD34+ CB cells as measured by dilution of CFSE at day 12 of culture on BMSCs or only in the presence of cytokines (SCF, TPO, FLT3L) with or without addition of LPS (10 μg/ml) as indicated. Gates and numbers indicate percentage of CD34+ cells that divided ≤3 and ≥4 times. Results from one representative out of three independent experiments with each different CD34+ CB cells and BMSC donors are shown. d CFU activity of hematopoietic cells previously cultured on BMSCs for 12 days with or without LPS, and cells previously cultured for 12 days only in the presence of cytokines with or without LPS as described in (a). Each 2 × 103 cells were subjected to methylcellulose cultures containing SCF, FLT3L, TPO, IL-3, IL-6, IL-11, GM-CSF, and EPO. One representative experiment out of three with each different CD34+ cord blood cells and different BMSC donors is shown. e and f Analysis of the BM and thymus of Rag2−/−γc −/− mice 4 weeks after receiving transplants of 2–3 × 105 human hematopoietic cells recovered after 12 days of culture on BMSCs in the presence or absence of LPS. Representative analysis out of two independent experiments with five transplanted mice per experiment is shown. Statistically significant differences are indicated (*p < 0.5)
To assay myeloid differentiation potential, remaining hematopoietic cells from primary BMSC co-cultures or cytokine-only cultures were subjected to cytokine-supplemented CFU assays for full myeloid read out. Consistent with higher CD34+ cell content, hematopoietic cells recovered from LPS-stimulated BMSC co-cultures produced about 2.5-fold more myeloid colonies as compared to cells from unstimulated BMSC co-cultures. Furthermore, mixed myeloid and erythroid colonies could only be detected from cells derived from LPS-stimulated BMSC co-cultures (Fig. 2d). In contrast, CFU activity from cells that were pre-cultured in cytokines only was overall lower compared to CFU-activity from co-cultures, where no mixed myeloid and erythroid colonies could be detected and no relevant difference with respect to LPS addition was observed (Fig. 2d).
To next test lymphoid cell differentiation capacity, BMSC co-cultured cells were transplanted into irradiated newborn Rag2−/−γc −/− mice [14]. Low levels of human B cell and T cell engraftment were detected only in animals receiving hematopoietic cells from LPS-stimulated BMSC co-cultures (Fig. 2e, f and Table 1). These results demonstrate that upon LPS stimulation, primary human BMSCs produced soluble or membrane-bound factors that increased maintenance of immature human hematopoietic cells with myeloid and lymphoid developmental potential.
Collectively, our findings suggest that upon TLR activation, BMSC differently support hematopoiesis: (1) They increase the secretion of cytokines, which directly support myeloid differentiation. (2) They enhance the production of factors that are able to maintain immature hematopoietic progenitor cells. Whereas the first mechanism contributes to the rapid replenishment of consumed effector cells during an ongoing innate immune response, the latter seems to prevent the exhaustion of the HSPC pool during inflammation. Increased maintenance of HSPCs in LPS-stimulated co-cultures parallels recent in vivo finding of LPS injection driving dormant HSC into cycle and self-renewal [22]. The results shown here point to an indirect activation of HSC maintenance by a most likely TLR4-expressing nonhematopoietic cellular compartment, which drives the production of various maintenance-associated factors. In addition, toll-like receptor ligation has been shown to affect the osteogenic potential of BMSCs, eventually reducing their differentiation capacity. Osteoblasts are important positive regulators of hematopoiesis as shown in mouse models of osteoblast-deficiency. LPS treatment could therefore negatively affect the HSC-supporting ability by reducing the number of mature osteoblasts. It is therefore tempting to speculate that BMSC function in parallel as positive and negative regulators of hematopoiesis, thereby counterbalancing each other in order to ensure a controlled activity during an ongoing immune response. Inflammation not only leads to a massive enhancement of granulopoiesis but also induces changes in systemic iron handling, inhibition of erythrocyte production, and reduction of erythrocyte life span. Anemia is therefore a comorbid condition in clinical settings which associates with poor outcomes. Bacterial infection drives the production and release of pro- as well as anti-inflammatory cytokines not only in the BM, the primary site of hematopoiesis, but also at multiple sites throughout the body. Cytokines which restrict erythropoiesis or which promote the turnover of erythrocytes could therefore be produced in the BM or reach the BM via the blood stream and interfere with the general improvement of hematopoiesis elicited by BMSC.
As BMSC are known to enhance the expansion of HSCs during ex vivo co-culture [23, 24], the additional use of TLR-ligands could further optimize the expansion of HSCs for later clinical use.
References
Sacchetti B, Funari A, Michienzi S et al (2007) Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell 131:324–336
Sugiyama T, Kohara H, Noda M, Nagasawa T (2006) Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 25:977–988
Mendez-Ferrer S, Michurina TV, Ferraro F et al (2010) Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 466:829–834
Ding L, Saunders TL, Enikolopov G, Morrison SJ (2012) Endothelial and perivascular cells maintain haematopoietic stem cells. Nature 481:457–462
Greenbaum A, Hsu YM, Day RB et al (2013) CXCL12 in early mesenchymal progenitors is required for haematopoietic stem-cell maintenance. Nature 495:227–230
Park D, Spencer JA, Koh BI et al (2012) Endogenous bone marrow MSCs are dynamic, fate-restricted participants in bone maintenance and regeneration. Cell Stem Cell 10:259–272
Walenda T, Bork S, Horn P et al (2010) Co-culture with mesenchymal stromal cells increases proliferation and maintenance of haematopoietic progenitor cells. J Cell Mol Med 14:337–350
Wagner W, Roderburg C, Wein F et al (2007) Molecular and secretory profiles of human mesenchymal stromal cells and their abilities to maintain primitive hematopoietic progenitors. Stem Cells (Dayton, Ohio) 25:2638–2647
Kadereit S, Deeds LS, Haynesworth SE et al (2002) Expansion of LTC-ICs and maintenance of p21 and BCL-2 expression in cord blood CD34(+)/CD38(−) early progenitors cultured over human MSCs as a feeder layer. Stem Cells (Dayton, Ohio) 20:573–582
McNiece I, Harrington J, Turney J, Kellner J, Shpall EJ (2004) Ex vivo expansion of cord blood mononuclear cells on mesenchymal stem cells. Cytotherapy 6:311–317
Robinson SN, Ng J, Niu T et al (2006) Superior ex vivo cord blood expansion following co-culture with bone marrow-derived mesenchymal stem cells. Bone Marrow Transplant 37:359–366
Wang X, Cheng Q, Li L et al (2012) Toll-like receptors 2 and 4 mediate the capacity of mesenchymal stromal cells to support the proliferation and differentiation of CD34(+) cells. Exp Cell Res 318:196–206
Manz MG, Miyamoto T, Akashi K, Weissman IL (2002) Prospective isolation of human clonogenic common myeloid progenitors. Proc Natl Acad Sci U S A 99:11872–11877
Traggiai E, Chicha L, Mazzucchelli L et al (2004) Development of a human adaptive immune system in cord blood cell-transplanted mice. Science (New York, NY) 304:104–107
Bianco P, Robey PG, Simmons PJ (2008) Mesenchymal stem cells: revisiting history, concepts, and assays. Cell Stem Cell 2:313–319
Dominici M, Le Blanc K, Mueller I et al (2006) Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 8:315–317
Tomchuck SL, Zwezdaryk KJ, Coffelt SB, Waterman RS, Danka ES, Scandurro AB (2008) Toll-like receptors on human mesenchymal stem cells drive their migration and immunomodulating responses. Stem Cells (Dayton, Ohio) 26:99–107
Pevsner-Fischer M, Morad V, Cohen-Sfady M et al (2007) Toll-like receptors and their ligands control mesenchymal stem cell functions. Blood 109:1422–1432
Liotta F, Angeli R, Cosmi L et al (2008) Toll-like receptors 3 and 4 are expressed by human bone marrow-derived mesenchymal stem cells and can inhibit their T-cell modulatory activity by impairing Notch signaling. Stem Cells (Dayton, Ohio) 26:279–289
Sioud M, Floisand Y, Forfang L, Lund-Johansen F (2006) Signaling through toll-like receptor 7/8 induces the differentiation of human bone marrow CD34+ progenitor cells along the myeloid lineage. J Mol Biol 364:945–954
De Luca K, Frances-Duvert V, Asensio MJ et al (2009) The TLR1/2 agonist PAM(3)CSK(4) instructs commitment of human hematopoietic stem cells to a myeloid cell fate. Leukemia 23:2063–2074
Takizawa H, Regoes RR, Boddupalli CS, Bonhoeffer S, Manz MG (2011) Dynamic variation in cycling of hematopoietic stem cells in steady state and inflammation. J Exp Med 208:273–284
Li N, Feugier P, Serrurrier B et al (2007) Human mesenchymal stem cells improve ex vivo expansion of adult human CD34+ peripheral blood progenitor cells and decrease their allostimulatory capacity. Exp Hematol 35:507–515
Fei XM, Wu YJ, Chang Z et al (2007) Co-culture of cord blood CD34(+) cells with human BM mesenchymal stromal cells enhances short-term engraftment of cord blood cells in NOD/SCID mice. Cytotherapy 9:338–347
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest. All authors have read the journal’s authorship agreement and policy on disclosure of potential conflicts of interest.
Funding
This work was in part funded by a START grant of the Medical faculty of RWTH Aachen University to P.Z.
Additional information
Markus G. Manz and Tim H. Brümmendorf contributed equally to this work.
Rights and permissions
About this article

Cite this article
Ziegler, P., Boettcher, S., Takizawa, H. et al. LPS-stimulated human bone marrow stroma cells support myeloid cell development and progenitor cell maintenance. Ann Hematol 95, 173–178 (2016). https://doi.org/10.1007/s00277-015-2550-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00277-015-2550-5