
Abstract
In this paper, based on the analysis of a long-term energy balance monitoring programme, a Bowen ratio-based method (BR) was proposed to resolve the lack of closure of the eddy covariance technique to obtain reliable sensible (H) and latent heat fluxes (λE). Evapotranspiration (ET) values determined from the BR method (ETc,corr) were compared with the upscaled transpiration data determined by the sap flow heat pulse (HP) technique, evidencing the degree of correspondence between instantaneous transpirational flux at tree level and the micrometeorological measurement of ET at orchard level. Using the BR-corrected λE fluxes, a crop ET model implementing the Penman–Monteith approach, where the canopy surface resistance was determined from standard microclimatic variables, was applied to determine the crop coefficient values. The performance of the model was evaluated by comparing it with the sap flow HP data. The results of the comparison were satisfactory, and therefore, the proposed methodology may be considered valid for characterizing the ET process for orange orchards grown in a Mediterranean climate. By contrast to reports in the FAO 56 paper, the crop growth coefficient of the orange orchard being studied was not constant throughout the growing season.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Analyses carried out on water use in the Mediterranean region have shown that on average 72 % of available water is used for irrigation purposes (Hamdy 1999) and about 99 % of water uptake from the soil by plants is lost as evapotranspiration (ET). Therefore, accurately estimating ET rates is crucial in water resource management in arid and semi-arid environments where irrigation is necessary and water is quite scarce and expensive.
At present, different methods are available for determining ET. Some methods are more suitable than others because of their accuracy or cost, or because they are particularly suitable for the given space and time scales. Often it is necessary to predict ET, so it must be modelled. Among the indirect methods of measuring ET rates, the most accurate techniques are the micrometeorological ones (i.e. Bowen ratio, eddy covariance, aerodynamic methods, radiation temperature methods, etc.) (Heilman et al. 1996; Villalobos et al. 2000; Wullschleger et al. 2000), and thus, from an energy point of view, ET may be considered as equivalent to the energy used to transport water from the inner cells of the leaves and plant organs and from the soil to the atmosphere. In this case, it is ‘latent heat’ (λE, with λ latent heat of vaporization equal to 2.45 × 106 J kg−1 at 20 °C) and expressed as energy flux density (W m−2). When ET is considered as latent heat flux density, it is worth considering all the energy components acting above a vegetated surface, that is, the energy balance may be written as:
where all the terms are in W m−2, R n is the net radiation from balancing all the radiations above the crop and is directly measurable, G is the soil heat flux, also directly measurable, and H is the sensible heat flux, which can be directly measured and/or estimated by micrometeorological techniques. R n − G is commonly termed available energy, and the surface heat storage at daily or longer time scales is negligible. The energy available at the land surface used for photosynthesis (typically less than 1 % of net radiation) can be neglected. The fluxes may be considered conservative, that is, when they are only vertical and expected to remain constant, independent of their height, over a horizontal homogeneous surface.
Since the mid-1990s, the Fluxnet network has provided long-term high-quality observations of heat and mass exchanges between the land surface and atmosphere, for a wide range of ecosystems. The methods which measure land surface–atmosphere exchanges of heat and mass are numerous and have been summarized in a number of publications (Twine et al. 2000; Baldocchi 2003; Finnigan 2004). Eddy covariance (EC) is the most widely used technique at flux-tower sites worldwide. However, an analysis by Wilson et al. (2002) of surface flux measurements at 27 EC sites distributed across North America and Western Europe showed that energy budget closure (Eq. 1) evaluations were absent for all the investigated sites. Aubinet et al. (2000) reported a similar finding based on analyses of EC data collected at European sites where typically, annual energy closure ranges between 5 and 30 % (Guo et al. 2009). Many studies have provided explanations for energy imbalance (Twine et al. 2000; Oncley et al. 2007), including suggestions that there are: (1) sampling errors associated with different measurement source areas for the energy components, (2) systematic bias caused by calibration, inherent time response and the mounted structures of instruments, (3) neglected energy sinks, (4) the loss of low- and/or high-frequency contributions to turbulent flux, and (5) neglected advection of scalars. Consequently, the sensible heat and latent heat associated with turbulent movement are systematically underestimated. The current inability of EC data to close the energy budget is a well-known issue, which has led several authors to emphasize the necessity of finding a way to deal with it (Mahrt 1998; Wilson et al. 2002; Baldocchi 2003; Liu et al. 2006). In order to improve the usefulness of eddy covariance measurements, Twine et al. (2000) suggested adjusting sensible and latent heat to force the energy equilibrium. However, some researchers have pointed out the difficulty of applying and evaluating ecosystem models using flux measurements which lack energy closure (Kustas et al. 1999; El Maayar et al. 2008) and the imperative need for resolving observed energy budget imbalances in measured data prior to their use in test models. Despite all these recommendations, researchers are still testing land surface models in which the energy budget is assumed to be closed, using EC data which do not generally satisfy the conservation of energy (Zhang et al. 2005; Ju et al. 2006; Kucharik et al. 2006).
Therefore, micrometeorological approaches are often difficult to apply, both from theoretical and technical points of view. They also require large flat fields and constant technical assistance. Nevertheless, micrometeorological methods are currently indispensable for the calibration of other methods.
A variety of methods are available to measure, or estimate, plant water use in the field. Sap flow, for example, is a suitable and relatively simple method for measuring plant transpiration (Cohen et al. 1993; Trambouze et al. 1998; Rana et al. 2005; Motisi et al. 2012). Sap flow measurements can be taken both in herbaceous and woody plants and in any conductive organ, including roots. Depending on the method, measurements are taken in the part of the conductive organ where the sensors are located, or in the whole perimeter of the conductive organ. Some methods are suitable for stems of small diameters, while others can be used in large trees. Calibration is convenient in all cases, being compulsory for the invasive methods, since the insertion of the probes alters the xylem characteristics.
The heat pulse velocity (HPV) system presented by Green and Clothier (1988) is based on the compensation heat pulse (CHP) method (Swanson and Whitfield 1981). The method allows to monitor tree transpiration and explores the short-term water-use dynamics of trees (Green et al. 1989; Moreno et al. 1996).
However, since sap flow is measured at the branch or trunk scale, transpiration measurements should be scaled up to the whole stand (Zhang et al. 1997). Scaling-up procedures may vary according to the characteristics of the stand (Smith and Allen 1996).
The crop coefficient (K c) approach of estimating evapotranspiration in well-watered conditions is the most practical and recommended method (Allen et al. 1998). The concept of K c was introduced by Jensen (1968) and further developed by the other researchers (Doorenbos and Pruitt 1975, 1977; Burman et al. 1980; Allen et al. 1998). The crop coefficient is the ratio of the actual crop evapotranspiration (ETc) to reference crop evapotranspiration (ET0), and it integrates the effects of characteristics that distinguish field crops from grass, like ground cover, canopy properties and aerodynamic resistance. For irrigation scheduling purposes, daily values of crop ETc can be estimated from crop coefficient curves, which reflect the changing rates of crop water use over the growing season, if the values of daily ET0 are available. FAO paper 56 (Allen et al. 1998) presents a procedure to calculate ETc using three K c values that are appropriate for four general growth stages (in days) for a large number of crops.
To make the use of K c operational, research and experiments have been carried out worldwide, and they have led to the determination of the average value that K c may take in the course of the season over the years. It is worth highlighting that the K c is affected by all the factors that influence soil water status, for instance, the irrigation method and frequency (Doorenbos and Pruitt 1977; Wright 1982; Consoli et al. 2006), the weather factors, the soil characteristics and the agronomic techniques that affect crop growth (Stanghellini et al. 1990). Consequently, the crop coefficient values reported in the literature can vary even significantly from the actual ones if growing conditions differ from those where the mentioned coefficients were experimentally obtained (Tarantino and Onofrii 1991).
Therefore, the general objective of this study was to identify a useful procedure for correcting the energy budget measurements of sensible and latent heat fluxes to measure and model the evapotranspiration (ET) of irrigated orange orchards in semi-arid Mediterranean conditions. The key points of the paper may be outlined as follows:
-
The evaluation of the Bowen ratio (BR) correction method to solve the unclosed surface energy balance at the study site.
-
The evaluation of the performance of the sap flow method (T SF) to measure actual evapotranspiration by comparison with the micrometeorological method (corrected eddy covariance data).
-
The development of a model of orange orchard evapotranspiration applying a Penman–Monteith approach, using standard meteorological variables as input to determine canopy resistance.
-
The evaluation of the crop coefficient method (K c) effectiveness to determine the water requirements of the orange orchard and to verify the suitability of the K c value proposed for a generic citrus crop.
Materials and methods
Site information and field measurements
The trial was carried out during the monitoring period 2010–2011 within an orange orchard located in Sicily, Southern Italy (Lentini, lat. 37°16′N, long. 14°53′E). This area has a semi-arid Mediterranean climate (annual mean air temperature 17 °C and rainfall less than 600 mm in the period 1990–2011). The experimental field (of about 20 ha) was planted with 15–25-year-old orange trees (Citrus sinensis (L.) Osbeck, cv. Tarocco Ippolito), grown in an orchard of about 120 ha. The site is flat and the orchard has trees with similar spatial distribution of about 4 m between trunks within rows and 5.5 m between rows. The soil surface is partially covered (10–15 %) by natural grass for most of the year. The mean canopy height was 3.75 m. Leaf area index (LAI, m2 m−2) was estimated using a Licor LAI-2000 (LI-COR® Biosciences) digital analyser and was found to be in the range of 4.0–4.7. For the dominant wind direction (mainly W and NW), the fetch was larger than 550 m. For the other sectors, the minimum fetch was 400 m (SE) (Castellví et al. 2012).
The crop was well-watered by irrigation supplied every day during the hot months (May–October). Water was supplied by drip irrigation, with 4 online labyrinth drippers per plant, spaced at 0.80 m, with a discharge rate of 4 l/h at a pressure of 100 kPa. Total irrigation, during the two seasons, amounted to about 5,500 m3 ha−1.
Continuous energy balance measurements were made from January 2010 to December 2011. Net radiation (R n, W m−2) was measured with two CNR 1 Kipp&Zonen (Campbell Scientific Ltd) net radiometers at a height of 8 m. Soil heat flux density (G, W m−2) was measured with three soil heat flux plates (HFP01, Campbell Scientific Ltd) placed horizontally 0.05 m below the soil surface. Three different measurements of G were selected: in the trunk row (shaded area), at 1/3 of the distance to the adjacent row, and at 2/3 of the distance to the adjacent row. The soil heat flux was measured as the mean output of three soil heat flux plates. Data from the soil heat flux plates were corrected for heat storage in the soil above the plates. The heat storage (ΔS) was quantified in the upper layer by measuring the rate of temperature change. The net storage of energy (ΔS) in the soil column was determined from the temperature profile taken above each soil heat flux plate. Three probes (TCAV, Campbell Scientific Ltd) were placed in the soil to sample soil temperature. The sensors were placed 0.01–0.04 m (z) below the surface; the volumetric heat capacity of the soil C v was estimated from the volumetric fractions of minerals (V m), organic matter (V o) and volumetric water content (θ). Therefore, G at the surface was estimated by measuring G′ at a depth of 0.05 m and the change in temperature over time of the soil layer above the heat flux plates to determine ΔS.
The air temperature and the three wind speed components were measured at two heights, 4 and 8 m, using fine wire thermocouples (76 μm diameter) and sonic anemometers (Windmaster Pro, Gill Instruments Ltd, at 4 m, and a CSAT, Campbell Sci., at 8 m). A gas analyser (CSAT, Campbell Sci.) operating at 10 Hz was deployed at 8 m. The raw data were recorded at a frequency of 10 Hz using two synchronized data loggers (CR3000, Campbell Sci.).
Low-frequency measurements were taken for air temperature and humidity (HMP45C, Vaisala), wind speed and direction (05103 RM Young), and atmospheric pressure (CS106, Campbell Scientific Ltd) at 4 and 8 m. Rainfall (AGR 100 Waterra, UK) was measured nearby.
The freely distributed TK2 package (Mauder and Foken 2004) was used to determine the first- and second-order statistical moments and fluxes on a half-hourly basis following the protocol used as a comparison reference described in Mauder et al. (2007). This software corrects for the errors in the wind speed vertical components, sensor separation and path-length averaging and eliminates spurious flux values.
The micrometeorological data set used for comparison included samples (of high- and low-frequency data) that passed the Foken’s quality control test up to level 7 (Mauder and Foken 2004). The test checks the assumptions of steady flow and developed turbulence invoked in the EC method. Then, variances and covariances are discriminated in levels of reliability. Up to level 7 (i.e. from 1 to 7), the data set includes high-quality flux measurements recommended for research purposes (up to level 3) and measurements that can be considered useful for routine applications and gap filling (from 4 to 7). The range −20 Wm−2 ≤ H EC ≤ 20 Wm−2 was excluded [taken as the EC measurement error (Foken 2008)].
Energy closure at the orange orchard and correction of measured sensible and latent heat fluxes
The EC method applied at the study site allows for estimates of H and λE by directly measuring fluctuations in the vertical wind velocity and scalar concentration (Arya 1988; Campbell and Norman 1998). By contrast with the Bowen ratio (BR) method, EC does not force the energy budget to close though, beneficially, it offers separate estimates of H and λE.
At this point, the problem, of course, is how best to close the energy balance for the EC method. As reported by Twine et al. (2000), there are two approaches. The first assumes that measurements of H are accurate so that λE can be calculated merely by subtracting G and H from R n (Eq. 1). This approach is known in the literature as the residual method. However, because no compelling evidence exists to confirm that the EC method only underestimates λE (Katul et al. 1999; Twine et al. 2000), a second approach uses the Bowen ratio to close the energy budget.
The BR approach assumes that for relatively homogeneous vegetated surfaces, R n − G measurements may be considered reliable and representative of the EC flux footprint. That is, any errors in R n − G measurements are necessarily much smaller than the energy imbalance determined from simultaneous EC measurements.
The BR approach then assumes that the EC technique provides correct estimates of the Bowen ratio (β = H/λE) even though it underestimates H and λE, as some studies tend to confirm (Barr et al. 1994; Blanken et al. 1997; El Maayar et al. 2008). Thus, rearrangement of Eq. 1 yields the following:
Then corrected estimates of λE are assumed to be given by Eq. 2, after which H can also be inferred from:
Equations (2) and (3) effectively redistribute the imbalance to H and λE according to their relative measured proportions.
In the following, the corrected latent and sensible heat fluxes refer to λE corr and H corr as calculated from data using Eqs. (2) and (3).
Crop evapotranspiration (ETc,corr) was calculated by transforming λE corr into millimetres of water. Calculating ETc,corr on a daily time scale was performed by adding hourly values for 24-h period.
Sap flow and soil evaporation measurements
Measurements of water consumption at tree level were performed by using the HPV (heat pulse velocity) technique. For HPV measurements, two 4-cm sap flow probes with 4 embedded thermocouples (Tranzflo NZ Ltd, Palmerston North, NZ) were inserted into the trunks of three trees. The probes were positioned at the north and south sides of the trunk at 50 cm from the ground and wired to a data logger (CR1000, Campbell Sci.) for heat pulse control and measurement; the sampling interval was 30 min. Data from the two probes were processed according to Green et al. (2003) to integrate sap flow velocity over sapwood area and calculate transpiration. So, the sapwood fraction of water was determined both on sample trees during the experiment and directly on the trees with the sap flow probes, at the end of the observation period. Wound-effect correction (Green et al. 2003) was performed on a per tree basis.
Scaling up the sap flow from a single tree to the field scale requires analysing plant size variability, to determine the mean of those monitored. This was obtained by analysing the spatial variability of plant leaf area (Jara et al. 1998). Thus, scaling was performed only on the basis of the ratio between orchard LAI and tree leaf area.
During each irrigation season, daily soil evaporation was observed over a 7-day period in spring/summer (June–July), summer (August–September) and autumn (October) with microlysimeter measurements for a total of 42 observations. Small undisturbed soil samples were located in rings of limited height which were closed at the bottom, weighed and reinstalled in the field. Measurements of soil evaporation E s were obtained using sets of four microlysimeters replicated three times for a total of 12. Microlysimeters, made from 3-mm-thick aluminium pipe, were 12.5 cm long with a diameter of 11.5 cm. A portable electronic balance was used to weigh the microlysimeters daily between 07.30 and 09.30 h solar time (Han and Felker 1997). Two microlysimeters were placed in dry soil and two in the saturated zone near the wet spots of the emitters.
Modelling crop evapotranspiration
Orange orchard crop evapotranspiration (ETc,mod) was analysed using the Penman–Monteith model. ETc,mod was calculated hourly using:
where A = R n − G (W m−2), ρ is the air density in kg m−3, Δ is the slope of the saturation pressure deficit versus temperature function in kPa °C−1, γ is the psychrometric constant in kPa°C−1, Cp is the specific heat of moist air in J kg−1°C−1, D is the vapour pressure deficit of the air in kPa, r c is the bulk canopy resistance in s m−1, and r a is the aerodynamic resistance in s m−1.
As highlighted by Rana et al. (2005), r c is not constant for irrigated crops, but varies depending on the available energy and vapour pressure deficit. Katerji and Perrier (1983) proposed calculating r c as:
where a and b are empirical calibration coefficients that require experimental determination; r* (s m−1) is given as (Monteith 1965):
In our study, the canopy resistance was calculated with Eq. 4, by introducing the λE corr values calculated with Eq. 2, together with the measured values of D and A, and the estimated values of r a:
where z is the reference point above the canopy, d (m), the zero plane displacement is estimated as a portion of the canopy height where an intermediate scaling is d = 0.75 h c (Brutsaert 1988), h c is the mean height of the orchard (3.75 m), k = 0.4 is the von Karman constant, and u* is the friction velocity (m s−1) measured by the EC method.
The obtained values of r c were combined with Eq. 5 to estimate parameters a and b.
This model is particularly suited to canopy crops covering orchard soils, as suggested by Villalobos et al. (2000), Wullschleger et al. (2000) and Rana et al. (2005).
The model was calibrated using 3 months of data (June–August) during the 2010 irrigation season. The results are shown in Fig. 1, where the r c/r a ratio is related to the r*/r a ratio. A linear curve fit resulted in a = 0.364 and b = 0.0422 (coefficient of determination R 2 = 0.6287).

Linear relationship between r c/r a and r*/r a during June–August 2010; r c, r a and r* are canopy, aerodynamic and climatic resistance, respectively
The final expression of the model on an hourly time scale is:
The calculation of the orange orchard ETc,mod on a daily time scale was obtained by adding hourly values of λEmod (from Eq. 8), after dividing by λ.
Determining the crop coefficient Kc
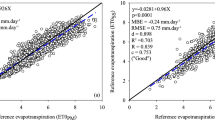
Crop evapotranspiration (ETc,mod) on a daily time scale was calculated with Eq. 8. Generally, crop coefficients are determined by calculating the ratio K c = ETc,mod/ETo, where ETc,mod is the evapotranspiration of a well-watered crop, and ET0 is the reference evapotranspiration calculated by the Penman–Montetih method (Allen et al. 1998). The variables used for ETo determination were measured in an agrometeorological station of the Sicilian Agrometeorological Service (SIAS) located 3.0 km away from the experimental field. The station was equipped with instruments for measuring standard meteorological variables (solar radiation, wind speed and direction, air temperature, relative humidity).
Results and discussion
Weather conditions
Figure 2 shows the daily weather variables during the two study periods. Air temperature (T a) reached a maximum in July, approximately 30 °C, while the minimum value occurred during February (about 5 °C). The relative humidity (RH) presented the inverse behaviour. The maximum wind speed (u) at 2 m above a standardized grass field was about 4.5 m s1 during January–June and October–November. Most precipitation (P) was during January–February and October–November; the irrigation seasons (May–September) were almost dry. The accumulated rainfall during 2010 was 588 mm, whereas for 2011 it was only 300 mm. Reference evapotranspiration (ETo) followed the oscillation of solar radiation (data not shown) with a maximum of about 9 mm day−1 during July and a mean value of about 4.2 mm day−1.

Daily values of weather variables during the study periods in 2010 (a, b) and 2011 (c, d). T a mean air temperature (°C), RH mean relative humidity (%), P precipitation (mm day−1), u mean wind speed (m s−1), ET o reference evapotranspiration (mm day−1)
BR-correction of measured sensible and latent heat fluxes
The lack of closure in the energy budget is commonly quantified by the relative difference between (R n − G) and (H + λE), expressed as a percentage: 100 × [((R n − G)/(H + λE)) − 1]. Figure 3 shows the average variation of monthly observed energy imbalance at the selected site. Assuming that R n and G measurements are rather accurate (Twine et al. 2000; Wilson et al. 2002) during both monitored years (H + λE) is underestimated at the site. This may arise from an underestimation of H or λE, or both. The mean annual energy imbalance is 29.3 % during 2010 and 31.1 % in 2011, showing peaks during winter and a variation of about 30 %.

Average monthly energy imbalance in measured data during 2010 and 2011
Despite eddy covariance (EC) being among the most advanced in situ measurement techniques which directly provides λE, the lack of closure of the EC-based energy balance is widely known (Wilson et al. 2002; Testi et al. 2006; de Teixeira et al. 2008). The λE data quality from the orange orchard was verified by studying the energy balance closure; fluxes (H + λE) and available energy (R n − G) were compared for the entire period (years 2010 and 2011) of measurement on an hourly time scale. The energy balance ratio, that is, the ratio of turbulent energy fluxes to available energy, was 0.89 in 2010 and 0.86 in 2011. The RMSE (root mean square error) for 1 h values of turbulent fluxes was 0.13 and 0.12 MJ m−2 h−1, during 2010 and 2011 highlighting how good the data set was.
The diurnal flux trend of individual components of the energy balance for the orange orchard showed that the latent heat flux (λE) was always in excess of the sensible heat flux (H) during daylight hours and the H was higher than the soil heat flux (G). At night, the eddy covariance results showed H and LE approaching zero.
Daily averages of energy balance fluxes are given in Table 1. Unstable atmospheric conditions predominated above the orchard, with the sensible heat flux (H) accounting for about 30 % of R n during both the monitoring periods. The significant leaf area index (LAI) of the orange crop (LAI of about 4–4.7 m2 m−2) caused little solar radiation to penetrate the canopy. As a consequence, the soil heat flux (G) on a daily scale was small and negative, with daily average values less than 1 % of R n. Negative values for G could be the result of both large LAI and frequent drip irrigation which keeps the soil thermal conductivity high. The largest part of R n was used as latent heat flux (λE), representing on average 67 % of R n during 2010 and 57 % of R n during 2011. The corresponding evaporative fractions (E F = λE/(R n − G)) were 0.70 and 0.60.
Monthly data show (Fig. 4) that after forcing (the BR approach) the measured energy balance data to close, the discrepancy between the available energy (R n − G) and turbulent fluxes (H corr + λE corr) tends to be neglected. In particular, H corr increased by 8.6 and 10 % in 2010 and 2011 with respect to EC measurements of H; λE tends to increase by 2 and 3.7 % in 2010 and 2011.

BR-corrected and uncorrected measurements from EC of monthly total sensible heat flux (H) and latent heat flux (λE) during 2010 (a) and 2011 (b) monitoring periods
Comparison between BR-corrected eddy covariance and sap flow measurements of evapotranspiration
Scatter plots of T SF versus ETc,corr hourly values showed general dependence with a saturation-type response, a good degree of linearity at low ETc,corr values and a lack of increase in T SF at high ETc,corr flux values (Fig. 5). For orange trees, fairly good linearity was observed both in the morning and afternoon values, whereas the midday values showed a weak relationship, with a small slope value, denoting lower xylem flux (SF values) in comparison with canopy transpiration as estimated by BR-corrected EC.

Scatter plots of hourly values of daytime upscaled trunk SF versus BR-corrected EC measurements of ET in orange. Observation period was July–August 2010
Analysis of the daily variation of average ETc,corr and T SF fluxes (Fig. 6) shows that T SF divergence from ETc,corr begins at about 09:00 local time. Midday T SF fluxes were almost steady for most of the period, while ETc,corr ones followed the daily trend of atmospheric evapotranspiration demand. Midday differences between T SF and ETc,corr denote a depletion of plant water content in relation to the imbalance between tree crown water loss by transpiration, as estimated by EC, and water transport from the tree’s root mass as estimated by SF. This imbalance is recovered in the afternoon and nocturnal hours, with higher T SF than ETc,corr fluxes.

Diurnal changes of upscaled SF in orange. Each data point represents the average of all the values in the observation period (July–August 2010)
Most of the differences here in water-use dynamics could be interpreted by tree capacitance. The imbalance between canopy transpiration and tree water uptake is revealed by a large hysteresis (data not shown), with higher afternoon SF values. It is interesting to note that the hysteresis loop appears specularly reflected, with a larger hysteresis in the morning-midday hours (Motisi et al. 2012).
Furthermore, as orchards in general are well coupled to the atmosphere, their transpiration is mainly regulated by the resistance that water vapour encounters in its movement from inside the leaf to the atmosphere (Villalobos et al. 2000). Canopy conductance (g c) is, therefore, an essential parameter for understanding the mechanisms of plant evaporation in orchards, but it is very difficult to measure at the correct scale.
Villalobos et al. (2009), analysing the canopy conductance of mandarin, found that it always peaked in the morning, followed by a lower plateau, and (sometimes) another small peak in the afternoon. This pattern of g c, with an early maximum later depressed in the hotter midday, is typical for water-stressed plants. A similar diurnal course of g c was also found in unstressed olives, another Mediterranean sclerophyllous tree crop, both at orchard (Villalobos et al. 2000) and leaf level (Moriana et al. 2002). Leaves, which reach low water potential during hot dry afternoons, accumulate water during the night. In the morning, the guard cells of the stomata are turgid and can open quickly. Later, when vapour pressure deficit (VPD) increases, the transpiration rate surpasses uptake. The leaf water potential drops and the stomata close, thus reducing g c and transpiration. This process goes on until water uptake can sustain the transpiration rate again and a new dynamic equilibrium is reached. This feedback system increases transpiration efficiency because the stomata are well open only during the coolest hours of the day (when the evaporative demand of the atmosphere is lower) and carbon can be assimilated at a lower cost in terms of water. The sensitivity of the feedback indicates adaptation to dry climates.
In this study, the difference between total values of T SF and ETc,corr during 2010 and 2011 was about 10 % which may be attributed to soil evaporation and which is not taken into account by the sap flow method.
The distribution of the evapotranspiration rate between soil evaporation (E s) and transpiration is difficult to compare to other studies, because of the small number of E s measurements available in our study and the difficulties of extrapolating them from different ground cover fractions. In this experiment, evaporation from the soil was 11 and 12 % of the total ETc,corr in 2010 and 2011. From the data of Moreshet et al. (1983), obtained in an orange orchard with the same percentage of ground cover by vegetation (about 80 %) as in this study, it is possible to calculate that their Es varied between 0.24 and 0.61 mm day−1. The lower values of E s are related to the lower energy available to evaporate water at the soil level due to the high canopy ground cover.
Crop evapotranspiration by a Penman–Monteith-type model
The comparison between daily crop evapotranspiration (ETc,mod) simulated by the model in Eq. 8 and T SF measured by the sap flow method is shown in Fig. 7. In particular, the empirical calibration coefficients of Eq. 5 were used to model ETc in 2011 too. For the available data set, the daily values of T measured by sap flow and modelled ET are fairly close during the crop cycle. The total values of the entire experimental periods are 913 and 883 mm for the sap flow measurements of transpiration during 2010 and 2011, and 1,008 and 984 mm for the modelled ETc during the 2 years with small differences, no higher than 3 %. The average daily values of ETc,mod and T SF are 3.9 and 3.4 mm day−1, during May–October 2010 and 3.7 and 3.2 mm day−1 in 2011.

Comparison between the transpiration values measured by the sap flow method (T SF) and evapotranspiration rates calculated by the model of Eq. 8 (ETc,mod) during 2010 (a) and 2011 (b)
The values of ETc,mod followed atmospheric demand in both growing seasons, being higher during May–October (from flowering to fruit maturation), with peaks of 6.7 and 6.0 mm day−1 during 2010 and 2011. The minimum values were around 1.2 mm day−1.
With regard to similar studies found in the literature, which used a calibrated heat pulse method, Cohen (1991) found average transpiration of about 3.0 mm day−1 in a high-density grapefruit orchard. Consoli et al. (2006) and Snyder and O’Connell (2007) reported daily ET peaks near 6.0 mm day−1 for clean-cultivated mature orange orchards in California using energy balance techniques. Rana et al. (2005) using EC measurement systems in citrus (Clementine) orchard in Mediterranean conditions (Southern Italy) found ET rates ranging from 2.0 to 7.0 mm day−1. Paco et al. (2006), using the same method in a peach orchard in Portugal, found ET values ranging from 1.4 to 4.0 mm day−1. de Teixeira et al. (2008), evaluating ET rates using micrometeorological measurements, showed average daily values of mango orchard ET ranging from 1 to 6.3 mm day−1. Sammis et al. (2004) used EC measurements to determine ET rates of flood-irrigated pecans in the USA; they found an average total ET of about 1,100 mm year−1.
Therefore, the methodology proposed in this work may be valid for characterizing the evapotranspiration process on a field scale.
Analysis of crop coefficient values
Figure 8 compares the crop coefficient calculated daily with the K c value given by Allen et al. (1998) for a generic citrus crop. These authors report a constant value of the crop coefficient between 0.45 and 0.65 (it is 0.65 in our experimental conditions), depending on the percentage of crop cover throughout the growth cycle. In this study, K c varies between 0.20 and 1.10, with a mean value of 0.68.

Comparison between the calculated daily crop coefficient (K c) and the value (constant) given by FAO 56 for the whole experimental period
During rainy periods, mainly at the start of the year, ETc.,mod rates exceeded ET0, producing daily K c values exceeding 1. As reported by Rana et al. (2005), the higher values of K c in the period January–June could be due to the following reasons: (1) the period coincides with phenological stages, the ‘flowering’ and ‘swelling of buds’ of active growth, when stoma conductance is usually high (Bethenod et al. 2000); (2) the period corresponds to days with high wind speed (Fig. 2) and vapour pressure deficit, which can cause high tree ET rates which are much greater than ETo.
Similar results were reported by Azevedo et al. (2003), who find higher K c values for mango orchard with peaks around 0.71 during the crop stages.
Many studies highlight greater accuracy when the crop coefficient curves are plotted with variables more closely related to crop development: LAI, canopy percentage shading the ground or thermal-based variables. This approach is considered an improvement over the FAO guidelines which suggest estimating K c values as a function of the length of the four phenological stages into which crop development is divided.
Linear relationships are reported between K c and LAI values for green bean and melon (Orgaz et al. 2006), grapevine (Williams et al. 2003), young olive orchard (Testi et al. 2004), and orange orchard (Consoli et al. 2006). In particular, Testi et al. (2004) find that the K c values determined in late autumn, winter and spring are usually high, variable and relatively independent of LAI or ground cover; during the summer, soil evaporation decreases and K c is lower, far less variable and LAI-dependent. These K c values are linearly correlated with LAI or ground cover: the authors proposed a linear model to predict it. This model has shown great robustness despite its empirical nature. Ayars et al. (2003) found that K c was a linear function of the amount of light intercepted by peach (Prunus persica L.) trees. Consoli et al. (2006) found nearly linear relationships between the normalized ratio for orange trees (K c over peak K c) and LAI, ground cover percentage (C g) and PAR light interception (LI). For comparable C g values of 20, 50 and 70 %, the observed K c values were 27, 31 and 43 % higher in the experiment than the K c values reported in FAO 56 (Allen et al. 1998).
Conclusions
Accurate knowledge of the partitioning of available energy between sensible and latent heat is needed to develop reliable tools for the study of short-term or long-term processes within agricultural crop ecosystems, which are particularly fragile when water availability is limited. This accurate knowledge requires the best possible data measurement and requires that the problem of closing measured energy budgets be resolved using rigorous procedures which respect the principle of energy conservation.
In our study, a long-term energy balance monitoring programme, integrated with sap flow measurements and biophysical ancillary data, was used to obtain reliable evapotranspiration estimates of irrigated orange orchards. In the implemented procedure, the Bowen ratio method was used to correct the eddy covariance measurements of sensible and latent heat fluxes for energy closure proving efficacious for orange orchards. In particular, after forcing the measured energy balance data to close, sensible heat flux increased by an average of about 9 % with respect to EC measurements of H; latent heat flux tends to increase by about 3 % during the monitoring period.
The BR-corrected crop evapotranspiration (ETc) rates were compared with upscaled transpirational data by sap flow heat pulse (T SF) method, showing a saturation-type response, with a good degree of linearity at low ETc,corr values and a lack of increases of T SF at high ETc,corr flux values. The difference between total values of T SF and ETc,corr during the monitoring period was about 10 %, which may be attributed to soil evaporation and which is not taken into account by the sap flow method.
By using the BR-corrected latent heat flux values, crop evapotranspiration rates (ETc,mod) were analysed and modelled, starting from a simple formulation based on the Penman–Monteith model, where canopy resistance was determined as a function of standard microclimatic variables. The calibration coefficients of the proposed model depend only on the crop and are valid for the study site. Modelled ETc helped calculate the crop coefficient (K c) of orange orchards during the growing seasons; this was compared with the FAO 56 approach based on a constant K c over the different growth stages.
Modelled ETc values were compared with daily transpiration (T SF) data measured by the sap flow method, with fairly good results. Simultaneous use of ETc,mod and T SF measurements provides an interesting experimental insight into the biophysical behaviour of tree crops.
References
Allen RG, Pereira LS, Raes D, Smith M (1998) Crop evapotranspiration, guidelines for computing crop water requirements. Paper No. 56, Rome
Arya SP (1988) Introduction to micrometeorology. Academic Press, London
Aubinet M, Grelle A, Ibrom A, Rannik U, Moncrieff J, Foken T, Kowalski AS, Martin PH, Berbigier P, Bernhofer C, Clement R, Elbers J, Granier A, Grunwald T, Morgenstern K, Pilegaard K, Rebmann C, Snijders W, Valentini R, Vesala T (2000) Estimates of the annual net carbon and water exchange of European forests: the EUROFLUX methodology. Adv Ecol Res 30:114–175
Ayars JE, Johnson RS, Phene CJ et al (2003) Water use by drip-irrigated late-season peaches. Irrig Sci 22:94
Azevedo PV, Silva BB, da Silva VPR (2003) Water requirements of irrigated mango orchards in northeast Brazil. Agric Water Manage 58:241–254
Baldocchi DD (2003) Assessing ecosystem carbon balance: problems and prospects of the Eddy covariance technique. Glob Change Biol 9:479–492
Barr AG, King KM, Gillespie TJ, denHartog G, Neumann HH (1994) A comparison of Bowen ratio and eddy correlation sensible and latent heat flux measurements above deciduous forest. Bound Layer Meteorol 71:21–41
Bethenod O, Katerji N, Goujet R, Bertolini JM, Rana G (2000) Determination and validation of corn crop transpiration by sap flow measurement under field conditions. Theor Appl Climatol 67:153–160
Blanken PD, Black TA, Yang PC, Newmann HH, Nesic Z, Staebler R, denHartog G, Novak MD, Lee X (1997) Energy balance and canopy conductance of a boreal aspen forest: partitioning overstory and understory components. J Geophys Res 102(D24):28915–28928
Brutsaert W (1988) Evaporation into the atmosphere. D Reidel Publishing Company, Dordrecht
Burman RD, Wright JL, Nixon PR, Hill RW (1980) Irrigation management-water requirements and water balance. In: Irrigation, challenges of the 80’s. Proceedings of the second national irrigation symposium, American Society of Agricultural Engineers, St. Joseph, MI, pp 141–153
Campbell GS, Norman JM (1998) An introduction to environmental biophysics, 2nd edn. Springer, New York
Castellví F, Consoli S, Papa R (2012) Sensible heat flux estimates using two different methods based on Surface renewal analysis. A study case over an orange orchard in Sicily. Agric For Meteorol 152:58–64
Cohen Y (1991) Determination of orchard water requirement by a combined trunk sap flow and meteorological approach. Irrig Sci 12(2):93–98
Cohen M, Girona J, Valancogne C, Ameglio T, Cruizat P, Archer P (1993) Water consumption and optimisation of the irrigation in orchards. Acta Hort 335:34–357
Consoli S, O’Connell N, Snyder R (2006) Estimation of evapotranspiration of different-sized navel-orange tree orchards using energy balance. J Irrig Drain Eng 132(1):2–8
de Teixeira AHC, Bastiaanssen WGM, Moura MSB, Soares JM, Ahmad MD, Bos MG (2008) Energy and water balance measurements for water productivity analysis in irrigated mango trees, Northeast Brazil. Agric For Meteorol 148:1524–1537
Doorenbos J, Pruitt WO (1975) Guidelines for predicting crop water requirements. Irrigation and Drainage Paper no. 24, FAO-ONU, Rome, Italy
Doorenbos J, Pruitt WO (1977) Guidelines for predicting crop water requirements, FAO-ONU, Rome. Irrigation and Drainage Paper no. 24 (rev.)
El Maayar M, Chen JM, Price DT (2008) On the use of field measurements of energy fluxes to evaluate land surface models. Ecol Model 214:293–304
Finnigan JJ (2004) A revaluation of long-term flux measurement techniques. Part II: coordinate systems. Bound Layer Meteorol 113:1–41
Foken T (2008) Micrometeorology. Springer, New York
Green SR, Clothier B (1988) Water use of kiwifruit vines and apple trees by the heat-pulse technique. J Exp Bot 39:115–123
Green SR, McNaughton KG, Clothier BE (1989) Observations of night-time water use in kiwifruit vines and apple trees. Agric For Meteorol 48:251–261
Green S, Clothier B, Jardine B (2003) Theory and practical application of heat pulse to measure sap flow. Agron J 95:1371–1379
Guo JX, Bian LG, Dai YJ (2009) Multiple time scale evaluation of the energy balance during the maize growing season, and a new reason of energy imbalance. Sci China Ser D Earth Sci 52:108–117
Hamdy A (1999) water resources in the Mediterranean region: from ideas to action. In: Background documentation for thematic session. Consultation of experts from MENA region on: World Water Vision-Water for Food and Rural Development. CIHEAM-IAMB, Valenzano, 27–29 May 1999, pp 1–23
Han H, Felker P (1997) Field validation of water-use efficiency of the CAM plant Opuntia ellisiana in south Texas. J Arid Environ 36:133–148
Heilman JL, McInnes KJ, Gesch RW, Lacano RJ, Savage MJ (1996) Effects of trellising on the energy balance of a vineyard. Agric For Meteorol 81:79–93
Jara J, Stockle CO, Kjelgaard J (1998) Measurement of evapotranspiration and its components in a corn (Zea Mays L) field. Agric For Meteorol 92:131–145
Jensen ME (1968) Water consumption by agricultural plants. In: Kozlowski TT (ed) Water deficits and plant growth, vol II. Academic Press, Inc., New York, pp 1–22
Ju W, Chen JM, Black TA, Barr AG, Liu J, Chen B (2006) Modelling multi-year coupled carbon and water fluxes in a boreal aspen forest. Agric For Meteorol 140:136–151
Katerji N, Perrier A (1983) Modélisation de l’évapotranspiration réelle ETR d’une parcelle de luzerne: rŏle d’un coefficient cultural. Agronomie 3(6):513–521
Katul G, Hsieh C-I, Bowling D, Clark K, Shurpali N, Turnipseed A, Albertson J, Tu K, Hollinger D, Evans B, Offerle B, Anderson D, Ellsworth D, Vogel C, Oren R (1999) Spatial variability of turbulent fluxes in the roughness sublayer of an even-aged pine forest. Bound Layer Meteorol 93:1–28
Kucharik C, Barford C, El Maayar M, Wofsy SC, Monson RK, Baldocchi DD (2006) Evaluation of a dynamic global vegetation model (DGVM) at the forest stand-level: vegetation structure, phenology, and seasonal and inter-annual CO2 and H2O vapor exchange at three AmeriFlux study sites. Ecol Model 196:1–31
Kustas WP, Prueger JR, Humes KS, Starks PJ (1999) Estimation of surface heat fluxes at field scale using surface layer versus mixed layer atmospheric variables with radiometric temperature observations. J Appl Meteorol 38:224–238
Liu H, Randerson JT, Lindfors J, Massman W, Foken T (2006) Consequences of incomplete surface energy balance closure for CO2 fluxes from open-path CO2/H2O infrared gas analyzers. Bound Layer Meteorol 120:65–85
Mahrt L (1998) Flux sampling errors for aircraft and towers. J Atmos Ocean Technol 15:416–429
Mauder M, Foken T (2004) Documentation and instruction manual of the eddy covariance software package TK2. Universität Bayreuth, Abt.Mikrometeorologie, Arbeitsergebnisse 26-44. http://www.geo.unibayreuth.de/mikrometeorologie/ARBERG
Mauder M, Foken T, Clement R, Elbers JAEW, Grünwald T, Heusinkveld B, Kolle O (2007) Quality control of CarboEurope flux data—part 2: inter-comparison of eddy-covariance software. Biogeosciences 5:4067–4099
Monteith JL (1965) Evaporation and environment. In: Fogg GE (ed) The state and movement of water in living organism. Proceedings of the XIX symposium on society for experimental biology. Academic Press, New York, pp 205–234
Moreno F, Ferna′ndez JE, Clothier BE, Green SR (1996) Transpiration and root water uptake by olive trees. Plant Soil 184:85–96
Moreshet S, Cohen Y, Fuchs M (1983) Response of mature "Shamouti" orange trees to irrigation of different soil volumes at similar levels of available water. Irr Sci 3:223–236
Moriana A, Villalobos FJ, Fereres E (2002) Stomatal and photosynthetic responses of olive (Olea europaea L.) leaves to water deficits. Plant Cell Environ 25(3):395–405
Motisi A, Consoli S, Rossi F, Minacapilli M, Cammalleri C, Papa R, Rallo G, D’urso G (2012) Eddy covariance and sap flow measurement of energy and mass exchange of woody crops in a Mediterranean environment. In: 8th international workshop on sap flow. Volterra (Italy), May 8–12
Oncley SP, Foken T, Vogt R et al (2007) The energy balance experiment EBEX-2000. Part I: overview and energy balance. Bound Layer Meteorol 123:1–28
Orgaz F, Testi L, Villalobos FJ, Fereres E (2006) Water requirements of olive orchards. II. Determination of crop coefficients for irrigation scheduling. Irrig Sci 24(2):77–84
Paco TA, Ferreira MI, Conceicao N (2006) Peach orchard evapotranspiration in a sandy soil: comparison between eddy covariance measurements and estimates by the FAO 56 approach. Agric Water Manag 85(3):305–313
Rana G, Katerji N, de Lorenzi F (2005) Measurements and modelling of evapotranspiration of irrigated citrus orchard under Mediterranean conditions. Agric For Meteorol 128:199–209
Sammis TW, Mexal JG, Miller D (2004) Evapotranspiration of flood-irrigated pecans. Agric Water Manage 69:179–190
Smith DM, Allen SJ (1996) Measurement of sap flow in plant stems. J Exp Bot 47:1833–1844
Snyder RL, O’Connell NV (2007) Crop coefficients for microsprinkler-irrigated, clean-cultivated, mature citrus in an arid climate. J Irrig Drain Eng 133(1):43–52
Stanghellini C, Bosma AH, Gabriels PCJ, Werkoven C (1990) The water consumption of agricultural crops: how crop coefficient are affected by crop geometry and microclimate. Acta Hortic 278:509–516
Swanson RH, Whitfield DWA (1981) A numerical analysis of heat-pulse velocity theory and practice. J Exp Bot 32:221–239
Tarantino E, Onofrii M (1991) Determinazione dei coefficienti colturali mediante lisimetri. Bonifica 7:119–136 (in Italian)
Testi L, Villalobos FJ, Orgaz F (2004) Evapotranspiration of a young irrigated olive orchard in southern Spain. Agric For Meteorol 121:1–18
Testi L, Orgaz F, Villalobos FJ (2006) Variations in bulk canopy conductance of an irrigated olive (Olea europaea L.) orchard. Environ Exp Bot 55:15–28
Trambouze W, Bertuzzi P, Voltz M (1998) Comparison of methods for estimating actual evapotranspiration in a row cropped vineyard. Agric For Meteorol 91:193–208
Twine TE, Kustas WP, Norman JM et al (2000) Correcting eddy-covariance flux underestimates over a grass land. Agric For Meteorol 103:279–300
Villalobos FJ, Orgaz F, Testi L, Fereres E (2000) Measurement and modeling of evapotranspiration of olive (Olea europea L.) orchards. Agric For Meteorol 13:155–163
Villalobos FJ, Testi L, Moreno-Perez MF (2009) Evaporation and canopy conductance of citrus orchards. Agric Water Manag 96:565–573
Williams DG, Cable W, Hultine K, Yepez EA, Er-Raki S, Hoedjes JCB, Boulet G, de Bruin HAR, Chehbouni A, Timouk F (2003) Suivi de la repartition de l’evapotranspiration dans une oliveraie (Olea europaea L.) a l’aide des techniques de l’eddy covariance, des flux de seve et des isotopes stables. Vemes Journees de l’Ecologie Fonctionnelle, 12 au 14 Mars 2003 a Nancy
Wilson K, Goldstein A, Falge E et al (2002) Energy balance closure at fluxnet sites. Agric For Meteorol 113:223–243
Wright JL (1982) New evapotranspiration crop coefficients. J Irrig Drain Div ASCE 108:57–74
Wullschleger SD, Wilson KB, Hanson PJ (2000) Environmental control of whole-plant transpiration, canopy conductance and estimates of the decoupling coefficient for large red maple trees. Agric For Meteorol 104:157–168
Zhang H, Simmonds LP, Morison JIL, Payne D (1997) Estimation of transpiration by single trees: comparison of sap flow with combination equation. Agric For Meteorol 87:155–169
Zhang Y, Grant RF, Flanagan LB, Wang S, Verseghy DL (2005) Modelling CO2 and energy exchanges in a northern semiarid grassland using the carbon- and nitrogen-coupled Canadian Land Surface Scheme (C-CLASS). Ecol Model 181:591–614
Acknowledgments
This work was carried out under the auspices of the Project Innovazioni e strumenti per l’adattamento dell’agricoltura ai cambiamenti climatici (ISAACC) (Innovations and tools for adapting agriculture to climatic change) under grant no. 594/2011 of the Sicilian Region. The authors wish to thank Azienda Tribulato (Lentini, SR) for its hospitality. The authors are also grateful to the Agrometeorological Service (SIAS) of the Sicilian Region and to CSEI Catania for their cooperation and support.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. Li.
Rights and permissions
About this article
Cite this article
Consoli, S., Papa, R. Corrected surface energy balance to measure and model the evapotranspiration of irrigated orange orchards in semi-arid Mediterranean conditions. Irrig Sci 31, 1159–1171 (2013). https://doi.org/10.1007/s00271-012-0395-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00271-012-0395-4