
Abstract
The effects of crop level and irrigation on water relations, yield, grape and wine composition were studied during two seasons in a Tempranillo vineyard in Spain. Irrigation was applied at two levels: R2 (with mild deficit irrigation applied during all the season) and R1 (with more severe water stress applied before veraison). Deficit irrigated vines were compared to a non-irrigated control. Crop levels imposed resulted in 11, 20, and 27 clusters per vine. Over all treatments, yield and ratio of leaf area to yield (LA: Y) were different between years: 4.4 and 16.3 t ha−1 and 1.72 and 0.88 m2 kg−1 in 2005 and 2006, respectively. In 2005, large differences in grape and wine composition occurred among non-irrigated and the irrigated treatments, but not between R1 and R2 treatments. Wines from non-irrigated vines were more acid, had higher total anthocyanins, and higher color intensity. In 2006, irrigation had less effect on grape and wine variables. The effect of shoot and cluster thinning on wine composition was different between seasons due to the different crop load values between years. Grape composition was negatively affected by high crop level only for values of LA: Y lower than 1.5 m2 kg−1.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In a vineyard, the training system employed defines the amount of light intercepted for a certain amount of leaf area and determines bud and fruit exposure to sunlight (Reynolds and Vanden Heuvel 2009). Within each genetic material and terroir, the vine water status and the vine yield (i.e. the crop level) or the balance between the sources (vine capacity to produce photoassimilates) and the sink demand (grape yield) (i.e. the crop load) are probably the major determinants of the vineyard performance and of the fruit composition (Jackson and Lombard 1993).
In arid and semi-arid environments, irrigation is a major tool to regulate soil water availability to vines. Under these conditions, supplying irrigation to ensure the potential vine evapotranspiration increases yield, and in occasions reduced wine quality (Williams and Matthews 1990; Salón et al. 2005; Valdés et al. 2009); though in other cases irrigation applied at 100% of crop evapotranspiration did not negatively affect grape or wine composition (Smart and Coombe 1983; Reynolds et al. 2007). Regulated deficit irrigation can be applied as a strategy to improve vine performance but minimizing the possible negative impact of high water status on wine quality (Acevedo-Opazo et al. 2010; Romero et al. 2010).
Water stress during the period from fruit set to veraison (i.e. the onset of ripening) heavily reduces fruit size, as the detrimental effect of soil water deficit on early fruit growth (Ojeda et al. 2001) cannot be normally compensated even if water supply returns at full dosage later in the season (Poni et al. 1994a). In this sense, a reduction in berry size might have a positive impact on wine phenolics content (Roby et al. 2004) and sugar concentration (Trought and Tannock 1996). If other berry characteristics, such as skin thickness, are not affected by lowering vine water status, smaller berries would have a higher skin-to-pulp ratio. In addition, a certain level of vine water stress during the pre-veraison period might be beneficial to regulate vine growth and vigor, helping to ensure an adequate cluster light environment. In this sense, our previous research (Intrigliolo and Castel 2010) has shown that in two out of four experimental seasons, a certain water restriction during the fruit set to veraison period leads to wines with higher color and larger phenolic and anthocyanins concentrations than the wines made from the well-watered plots.
In addition, because of the high dependence of fruit quality on various environmental and endogenous factors (Jackson and Lombard 1993), the overall effect of irrigation might change according to other cultural practices, particularly those affecting the crop level (Bravdo et al. 1984; Poni et al. 1994b). Vines with higher crop level seem to benefit more of a higher amount of irrigation both in terms of yield (Lakso et al. 1999) and of fruit composition (Hepner and Bravdo 1985). This is normally because under high yield a source limitation for carbohydrates derived from water stress might be more detrimental to proper fruit ripening, hence negatively affecting fruit and wine quality.
In grapevines, crop level can be regulated in several ways. Removing clusters during the growing season should mainly affect the fruit demand consequently decreasing the crop load. When crop is removed, vegetative growth can be stimulated (Poni et al. 1994a; Naor et al. 2002), but generally this effect is small (Freeman et al. 1979; Reynolds et al. 1994) or even nil if crop is removed late in the season (Valdés et al. 2009). In terms of fruit and wine composition, results presented in the literature have reported contrasting results, with cluster thinning leading to better fruit quality in some cases (Prajitna et al. 2007; Guidoni et al. 2002), but with no clear effect in others (Ough and Nagaoka 1984; Keller et al. 2005). In addition, cluster thinning is more expensive than shoot thinning alone, that, if performed very early in the season (e.g. before anthesis), can be carried out quicker and cheaper. However, shoot removal not only reduces the fruit demand (the crop level) but it also affects the plant leaf area; hence, the final effect on vine crop load (e.g. the ratio between leaf area and crop weight) might vary depending on the intensity and timing of this operation. In addition, the individual shoot leaf area might increase as a consequence of the shoot removal operation (Naor et al. 2002; Reynolds et al. 2005).
The main objective of this research was to test two different deficit irrigation regimes based on different volumes of water applied before veraison in comparison with rain-fed vines. Within each irrigation regime, three crop levels were tested, where crop levels were regulated by shoot thinning followed by cluster removal. The effects of these combined irrigation and crop level treatments on vine water status and leaf photosynthesis rates, growth, yield, and fruit and wine composition are analyzed.
Materials and methods
Site description
The experiment was carried out during two seasons (2005 and 2006) in a ‘Tempranillo’ vineyard (Vitis vinifera L.) planted in 1991 on 161–49 rootstock at a spacing of 2.45 by 2.45 m (1,666 vines/ha) in north–south oriented rows. The vineyard was located near Requena (39°29′N, 1°13′W, elevation 750 m), Valencia, Spain. Budbreak for Tempranillo in this area usually occurs by the mid April, anthesis by early June; veraison is reached by early August with harvest during late September and leaf fall at the beginning of November. In 2000, a drip-irrigation system was installed and vines trained to a vertically shoot positioned canopy on a bilateral cordon system. Winter pruning was carried similarly in all treatments by leaving 11–12 spurs per vine with two count buds per spur. Canopy height and width were about 1.3 and 0.40 m, respectively. Canopy management practices, all manually performed, included deshooting and shoot tip topping. Drip irrigation was applied with two pressure-compensated emitters of 2.4 l h−1 located at 60 cm each side of the vine. Irrigation was adjusted for rainfall and its frequency varied from 3 to 5 days per week. Water meters measured the amount applied to each replicate plot. All treatments were fertilized at a rate of 30–20–60–16 kg ha−1 of N, P2O5, K2O, and MgO, respectively.
The soil at the site is a Typic Calciorthid, with a clay loam to light clay texture, highly calcareous and of low fertility (0.66% of organic matter, and 0.04% of total nitrogen). It has a deep soil profile (>2 m), available water capacity is about 200 mm m−1 of soil depth and bulk density 1.43–1.55 t m−3.
Irrigation treatments and experimental design
The irrigation treatments applied water amounts at various fractions of estimated crop evapotranspiration (ETc) during the periods from anthesis to veraison and veraison to harvest. The first irrigation treatment (designated R1) consisted of applied water amounts at 25% of ETc from anthesis to veraison and then at 35% of ETc from veraison to harvest. The second irrigation treatment (R2) consisted of applied water amounts at 50% of ETc from anthesis to veraison and then at 35% of ETc from veraison to harvest. A non-irrigated control (the standard practice for the local viticulture) was also included in the study. The R1 irrigation treatment was slightly modified in 2006, and irrigation did not commence until midday stem water potential (Ψstem) reached −1.0 MPa. Thereafter, irrigation for R1 treatment was the same as that described for the R2 treatment. The selection of −1.0 MPa as a threshold for mild water stress was based on previous results obtained in the same vineyard (Intrigliolo and Castel 2007) and other findings reported by Girona et al. (2006). Despite the R1 irrigation scheduling was slightly modified over years, the objective was in both seasons to reduce water applications during the pre-veraison period, in an attempt to reduce berry size, while the same watering regime than the R2 treatment was applied after veraison during the ripening period.
Crop level treatments were 11, 20, and 27 clusters per vine (designated Low (L), Medium (M) and High (H), respectively) for the R1 and R2 irrigation treatments. Only the Low and Medium crop level treatments were used for the non-irrigated control. Crop level was adjusted by shoot thinning in mid-May and by additional cluster removal in early June if needed.
Each plot consisted of five rows with nine vines per row and the surrounding perimeter vines used as guards. The experiment was performed with a generalized incomplete block design (Steel and Torrie 1980) with three blocks and two replicated treatment combinations within each block for the irrigated treatments and a single replicate per block for the rain-fed treatments.
Crop evapotranspiration was estimated as product of reference evapotranspiration (ETo) and crop coefficient (Kc). Reference ETo was calculated by the Penman–Monteith equation (Allen et al. 1998). Environmental data were collected hourly at the vineyard site to include temperature, humidity, global radiation wind speed and direction, and rainfall with an automated meteorological station over bare soil in the plot. The estimated seasonal Kc (crop coefficient for fully watered vines) used varied with the phenological period and the expected pattern of leaf area development. Thus, from June to July, Kc was gradually increased from 0.16 to 0.70.
Stem water potential and leaf photosynthesis determinations
Stem water potential (Ψstem) was measured with a pressure chamber (Soil Moisture Corp., Santa Barbara, CA) following the procedure described by Turner (1981). Leaves chosen were located in the shaded portion of the canopy and were bagged 2 h before measurements were taken (from 1130 to 1230 h). Leaf blades remained covered with the plastic bags inside the pressure chamber. Four representative vines per treatment and two leaves per vine were measured for both R0 treatments and the low and high crop levels for the R2 irrigation treatment in 2005. During 2006, Ψstem was also measured in the R1 treatment, low and high crop levels, along with the treatments measured the previous year, as two pressure chambers were used simultaneously.
Leaf gas exchange was only determined in treatments R0L, R0M, R2L, and R2H with a portable IRGA system (Model ADC LC Pro+, The Analytical Development Co. Ltd., Hoddesdon, UK) on three mature, well-exposed leaves per vine in the same four vines per treatment used for Ψstem determinations. Leaf gas exchange measurements ended in mid-August of 2005 because of equipment failure.
Yield and vegetative growth determinations
Yield and the number of clusters per vine were determined at harvest on each experimental vine. Berry weight was determined on random samples of about 200 berries per treatment replicate. The exact number of berries was subsequently counted.
On 23 June 2005, a hail storm occurred that partly damaged clusters and shoot development. The impact of the hail storm on clusters was assessed by counting the number of berries affected in a random sample of 20 clusters per replicate.
Pruning weight and leaf area were determined in four selected vines per replicate. Leaf area was estimated after veraison when shoot growth had ceased. Leaf area per vine was estimated from a linear equation relating leaf area (Y, m2 per shoot and total (main plus laterals) shoot length (X, cm). This relationship was obtained, from 10 to 20 representative shoots of different lengths collected after veraison each year. Thus, leaf area per vine was calculated from the sum of each of the measured individual shoot lengths. Leaf area-to-yield ratio (LA: Y) was also calculated on the four selectioned vines per replicate.
Grape juice and wine analysis
Grape juice (i.e. must) components were determined in the same berry samples collected for berry fresh weight determination, which were crushed with a small hand-press, and the juice centrifuged. Soluble solids (Brix) were determined by refractometry. Juice pH and titratable acidity (TA) were determined by an automatic titrator. Organic acids (malic and tartaric in juice and wines) were analyzed by high-performance liquid chromatography following procedures described by Romero et al. (1993). Wine potassium was obtained by atomic absorption spectrophotometry. Ethanol in the wines was analyzed by gas chromatography. Wine color intensity (OD420 + OD520 + OD620), and total phenolics index (OD280) were determined by spectrophotometry in accordance with Ribereau-Gayon et al. (2000) and they were expressed in terms of absorbance units (AU). Anthocyanins (OD520 in HCl media) was also determined by spectrophotometry. All analytical determinations were duplicated.
Microvinification procedures
Grapes from the different treatments were harvested on the same day (or with one day difference), when a minimum 21° Brix was reached, and were transported to the experimental winery in field boxes. Vinifications were performed at “Estación Viticultura y Enología Requena” separately on samples of about 30 kg from each plot, (six and three vinifications per treatment for the irrigated and non-irrigated treatments, respectively). Grapes were mechanically crushed, de-stemmed, and fermented at about 25°C in stainless steel containers. All wine lots were inoculated with a commercial yeast strain (L-2056, Danstar Ferment AC, Zug Switzerland) at 100 mg kg−1. Skin contact time was 7 days, and during this time they were punched down automatically every 4 h. After alcoholic fermentation, they were racked off and malolactic bacteria (Oenococcus oeni) inoculated. They were again racked off, sulfited at 100 mg l−1 K2S2O5, decanted and bottled. Wine storage time was the same for all treatments and years. Analysis of the wines was performed at the same time for all treatments, just before the inoculation with malolactic bacteria about 1 month after grapes were crushed.
Statistical analysis
Analysis of variance was performed using the mixed procedure of the SAS statistical package (version 8.2; SAS Institute, Cary, NC). Data from both seasons were initially analyzed together. Since the treatment-by-year interaction was for most of the variables determined highly significant, each season was then analyzed separately. Within each season, differences among treatments were assessed by Duncan multiple range test at P < 0.05.
Results
Climatic conditions, stem water potential and leaf photosynthesis
The climate at the vineyard site is continental and semiarid with average annual rainfall of 430 mm of which about 65% falls during the dormant period. However, both, 2005 and 2006 were drier seasons, particularly 2005 with rainfall from Nov 04 to Oct 05 of 355 mm. In the 2006 season yearly rainfall was 381 mm, of which 149 mm occurred from April to harvest. During each phenological period, except for the after harvest period, the estimated crop evapotranspiration surpassed the irrigation applied plus rainfall (Table 1). As it was planned, the largest deficit was recorded during the veraison to harvest period.
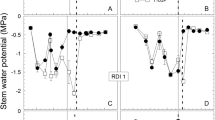
Irrigated vines had higher Ψstem and Pn compared to the non-irrigated control (Fig. 1). By the middle of the growing season (DOY 190 to 230), irrigated vines had Pn ranging from 10 to 15 μmol CO2 m−2 s −1, while Pn for the non-irrigated control ranged from 2 to 6 μmol CO2 m−2 s −1. Midday Ψstem during the summer months for the non-irrigated control vines averaged −1.29 and −1.37 MPa in 2005 and 2006, respectively. The delay in applying water to the R1 treatment in 2006 resulted in a decrease of Ψstem compared to the R2 treatment during that time frame. Once irrigation started, R1 treatment tended to maintain the highest Ψstem among all treatments (Fig. 1).

a and c Stem water potential measured at midday (Ψstem), and b and d leaf net photosynthesis rate (Pn) during the 2005 and 2006 seasons. Values are treatment means ± standard error of 8 and 12 leaves for Ψstem and Pn, respectively. Pn and Ψstem were measured only in the selected treatments shown. Daily rainfall events and amounts are included as bars originating from the x-axis, the downward arrow indicates the initiation of irrigation, and the upward arrow indicates the inception of veraison in (a) and (c)
Within each irrigation regime, the crop level did not affect leaf photosynthetic rate (Fig. 1). Pooling data for seasons and treatments, Pn decreased as vine water status was reduced, particularly when Ψstem went below −0.9 MPa (Fig. 2). However, there were no differences in Pn between crop levels (Fig. 2). Besides, the effect of crop level on Ψstem was not clear in 2005 (Fig. 1). In 2006, only in the non-irrigated vines, crop level had a more consistent effect throughout the season, with the medium crop level having around −0.1 MPa lower Ψstem than the low crop level (Fig. 1).

Relationship between midday measurements of stem water potential (Ψstem) and leaf net photosynthesis (Pn). Values are treatment means across both seasons separated for either the low or medium and high crop level
Vegetative growth, berry growth, and yield
Irrigation stimulated vegetative growth, resulting in greater leaf area per vine both years (Table 2). In 2006, differences in leaf area between R1 and R2 treatments were also significant. The high crop level reduced leaf area per shoot, but leaf area per vine increased in the more cropped vines because of the more shoots per vine left.
Non-irrigated vines had lower berry weight throughout the course of the season (Fig. 3). Before veraison, the R1 treatments had lower berry fresh weight than the R2 ones. The detrimental effect of the water restrictions applied before veraison on the berry fresh weight of the R1 treatments could not be fully compensated during the post-veraison period when both R1 and R2 treatments had the same watering regime. In general, the crop level had a much weaker effect on berry growth than irrigation. Nonetheless, the low crop levels had larger berry weights than the medium and high treatments on both seasons (Table 3). Differences in berry fresh weight due to irrigation or crop level were mainly due to reductions in growth rate, because the irrigation or the crop level did not affect plant phenological development nor the length of the berry growth cycle.

The change in berry fresh weight for the various treatments in 2005 and 2006. Values are treatment means of three replicates per treatment. Bars represent one standard error. Meaning of treatment legends: R0 non-irrigated; R1 irrigated at 25% ETc from anthesis to veraison and at 35% ETc from veraison to harvest; R2 irrigated at 50% ETc from anthesis to veraison and at 35% ETc from veraison to harvest. Crop level treatments were 11, 20, and 27 clusters per vine (designated Low (L), Medium (M) and High (H), respectively)
In both years, irrigation increased yield, with respect to non-irrigated (Table 3). This yield increment was due to the increases in both berry size and berry number per cluster produced by irrigation. In 2005, berries per cluster were higher in the R2 treatments than in the R1, but surprisingly the reverse pattern was observed in the next season. Crop level increased yield in the three irrigation treatments, in proportion to the number of clusters per vine.
There were large differences between seasons in the vine crop load estimated from the LA: Y ratio (Table 2). However, the effect of irrigation was consistent between years with the irrigated vines having lower LA: Y ratio.
Fruit and wine composition
In 2005, must total soluble solids concentration decreased as the irrigation volume applied increased (Table 4). In the following season, irrigation did not affect must sugar levels. Crop level had no effect on must Brix level in 2005, but there was a clear decreasing trend as crop level increased in 2006. In both seasons, irrigation increased the must titratable acidity, but crop level did not clearly affect this variable.
The effect of irrigation and crop level on wine ethanol content was in agreement with differences reported in must sugars levels (Table 5) and showed good correlation (r 2 = 0.52, P < 0.001). The effect of irrigation on the titratable acidity of the wine was only significant in 2005 when irrigation decreased it. Irrigation increased the malic acid concentration but tended to decrease the tartaric acid level in the wines. As a consequence, wine pH was significantly higher in the irrigated vines. On the other hand, crop level did not affect wine titratable acidity or the concentration of the main organic acids. Wine potassium levels were higher in the R1 and R2 treatments than in the R0 treatments. In addition, wine potassium levels increased in the low crop treatments (Table 5).
Supplemental irrigation reduced anthocyanins and total phenolics concentration in the berries as well as in the resulting wines (Tables 6, 7). This was particularly noticeable in 2005, where the R1 and R2 treatments had lower total and extractable berry anthocyanins content, and as consequence, lower color and anthocyanins concentration in the wine. The effect of crop level on fruit and wine phenolics content was instead only clear in 2006, when particularly the low cropping vines had higher concentrations of anthocyanins in wines and whole berries (Tables 6, 7).
Pooling data across irrigation and crop level treatments and seasons, there was a negative linear relationship between vine yield and wine total phenolics and anthocyanins concentration (Fig. 4). These same wine components were instead less related with the vine leaf area-to-yield ratio (Fig. 5).

The relationships between total wine phenols (a) and wine anthocyanins (b) and yield across year and treatment. Data points represent values from each experimental plot. ***Significant at P < 0.001. Meaning of treatment legends as in Fig. 3

The relationships between total wine phenols (a) and wine anthocyanins (b) and the leaf area-to-yield ratio (LA: Y) across year and treatment. Data points represent values from each experimental plot. ***Significant at P < 0.001. Meaning of treatment legends as in Fig. 3
Discussion
Stem water potential and leaf photosynthesis
Results presented in Figs. 1 and 2 suggest a lack of influence of crop level on gas exchange. This is in agreement with reports for other grapevine cultivars (Poni et al. 1994b; Edson et al. 1995). Presumably this was due to the fact that cluster-to-shoot ratios were very similar among treatments, as crop level was regulated mainly by shoot thinning and practically all shoots retained had only one cluster. For instance, Naor et al. (1997) reported that Sauvignon Blanc vines with two clusters per shoot had higher assimilation rates than plants with only one cluster per shoot.
The stem water potential and photosynthesis seasonal variations were mostly dependent on the irrigation regime. However, it should be noted that in 2006 the R1 treatments, once irrigation was resumed at similar rates than the R2 vines, had higher Ψstem (Fig. 1). This was probably a consequence of the lower leaf area of these pre-veraison stressed vines. Similarly in 2006, the fact that in rain-fed vines the medium crop level had lower Ψstem than the low one could be explained considering the larger leaf area of the medium cropping vines because of the more shoots retained during shoot thinning.
With data pooled across seasons, there was not a very tight relationship between plant water status and photosynthesis (Fig. 2). An attempt was made to relate the residuals of the Ψstem-Pn relationships with some environmental variables such as (average air vapor pressure deficit, air temperature, wind speed, and solar radiation). However, no significant relationships were found, suggesting that the Ψstem-Pn relationship might change across the season due to endogenous factors related with differences in plant phenological development (Naor 2008). However, despite the scatter, it could be derived a threshold of −0.9 MPa of Ψstem when a further decrease in plant water status detrimentally affects assimilation rates. Interestingly, in a previous study, we also showed that Ψstem around -1.0 MPa was found as a threshold for the onset of mild water stress based on trunk shrinkage data analysis (Intrigliolo and Castel 2007). A similar threshold of Ψstem-Pn relationship was also obtained for Riesling vines grown in New York (A. N. Lakso, Cornell University, personal communication). On the other hand, in other cases, a more linear relationship between plant water potential and stomatal conductance has been reported with a decline in leaf conductance at stem water potentials higher than what is reported here (Shackel 2007; Naor 2008).
Irrigation effects on grape and wine composition
Irrigation affected grape and wine composition in a slight different fashion in both seasons, with a less negative impact on wine composition, when compared to rain-fed, in 2006 than in 2005. This might be attributed to the differences in crop level and crop load between seasons, as water stress experienced by rain-fed vines was similar in both years. In 2005, crop demand for photoassimilates was very low, because of the low yield levels. Under this situation, the source limitation derived by vine water stress did not impair berry sugar accumulation and ripening. In fact, rain-fed berries and wines were more concentrated in terms of sugar and phenolics than the irrigated ones. In 2006, under a much larger crop demand, the supplemental irrigation applied on vines with limiting LA: Y was probably more necessary than in 2005 to ensure a proper vine source capacity. As a consequence, in 2006, irrigation even increased wine alcohol content and its detrimental effects on wine phenolics concentration were much less noticeable than in the first experimental season.
The effects of irrigation on must and wine organic acid concentrations and on wine pH were consistent between seasons. Irrigation increased wine pH similarly to our previous findings (Intrigliolo and Castel 2008) and also to another experiment with Carignane vines (Freeman and Kliewer 1983). However, previous irrigation trials with Tempranillo vines grown in other regions of Spain (Esteban et al. 1999, 2002) concluded that wine pH decreased with the application of supplemental irrigation. This suggests that even in the same variety the impact of the supplemental irrigation on wine pH might be different according to other factors probably related with the soil and the environmental conditions.
The fact that the irrigation increased wine pH is a negative effect. This is because wines with high pH are more susceptible to microbiological contaminations and hence less prone to aging. This effect was due to the differential effect reported of irrigation on the main wine organic acids. Irrigated vines had more vegetative growth, as reflected by the greater leaf area (Table 2). This larger canopy probably reduced cluster exposure to direct solar radiation and therefore cluster temperature—conditions favorable for the retention of malic acid and counteracting the dilution effect by irrigation because of larger berries. Tartaric acid concentrations decreased in irrigated wines. This was most likely because this acid is less affected than malic acid by environmental conditions (Ruffner 1982), and thus, its concentration was probably more determined by the dilution effect as well as by increased precipitation of bitartrate potassium salts (Iland and Coombe 1988), since wine potassium levels were also higher in the irrigated vines (Table 5). Given that malic is a weaker acid than tartaric (i.e. malic acid has higher pKa and it dissociates incompletely), the overall effect of irrigation on wine pH was to increase it.
In 2006, the delay in the onset of irrigation applied pre-veraison to the R1 treatment reduced berry fresh weight when compared with an earlier irrigation start as applied in the R2 treatments. These results are in agreement with the general observation that even mild pre-veraison water stress reduces berry size (Poni et al. 1994a; McCarthy 1997). However, the pre-veraison irrigation reduction had some positive effect on wine composition, only in 2006 when the R1 treatment had higher amount of wine anthocyanins (Table 7). Further seasons of investigation are hence needed in order to ultimately suggest growers to apply a pre-veraison water shortage under our conditions.
Crop level and crop load effects on berry and wine composition
Results reported corresponded with two seasons that were very different in terms of vine growth, and particularly yield levels. The very low number of berries per cluster reported in 2005 was the main reason for such low yield obtained in the first experimental year. This was in part due to the hail storm event on June 2005 that damaged berries and shoots (on average 15% of the berries were affected). In addition, during fruit set, there were 5 consecutive days with high temperatures recorded (maximum temperatures between 30 and 35°C) and high ETo values (up to 6–6.5 mm). We can speculate that this might have negatively affected fruit set. This unplanned feature allowed to test irrigation and yield effects under very different crop level and crop load conditions. On the other hand, within each season and irrigation regime, particularly between M and H crop levels there were small differences in the vine crop load (i.e. LA: Y). This was mostly because the yield level regulation adopted in this study reduced both the number of shoots and the number of clusters per vine. In addition, it should be noted that the different shoot densities among crop levels probably affected the cluster light exposure. However, no specific determinations were carried out to corroborate this hypothesis.
Among grape growers and wine makers there is a common, and often not scientifically proved, consideration that high yield and large berry size detrimentally affects wine composition (see review by Matthews and Nuzzo 2007). Only the first hypothesis seems to hold true under our experimental conditions. Pooling data across seasons and treatments yield alone could explain up to 82 and 65% of the variability in the wine phenolics and anthocyanins concentrations, respectively (Fig. 4). Yield, regardless of the cultural factor that influences the crop level at harvest, was then a crucial determinant of wine phenolic composition in our experimental conditions.
In contrast, across seasons and treatments, berry fresh weight was not related with wine composition (relationships not-shown), suggesting that berry fresh weight itself was not a determinant factor of wine composition. On the other hand, despite it is often assumed that berry size will decrease with an increase in yield (Matthews and Nuzzo 2007), in the present work, such effect was not found. In fact, for berry size the effect of the year was not statistically significant while the yields were 2.5–3 times larger in 2006 than in 2005 (Table 3).
It is also generally considered that vine performance and fruit and wine composition should be more closely related to any index expressing vine crop load (e.g. leaf area to yield, or the pruning weight to yield ratio) rather than just yield (i.e. crop level itself) (Lakso and Eissenstat 2005). The large difference in yield commented between seasons were related to very different LA: Y. In 2005 the LA: Y ratios were higher than 1.2 m2 kg−1 in all treatments, except in R2H, where it was 1.1 (Table 2). Instead in 2006 crop load was more variable with values ranging from 1.5 to 0.7 m2 kg−1. The lowest values were reached in the more irrigated and heavily cropped vines. These results might explain why grape and wine phenolic composition was more negatively affected by the high crop level in 2006 than in 2005.
Pooling data across seasons and treatments, wine total phenolics content and wine anthocyanins were reasonably well related to crop load (Fig. 5). It seems then that accumulation of phenolics and anthocyanins in wines might be detrimentally affected by high crop load for LA: Y below 1.5 m2 kg−1. This suggests that this value could be employed as a threshold for crop load management for irrigated Tempranillo grapes under our environmental conditions, trained as single narrow vertical shoot positioned curtain. It should be noted that this LA: Y threshold is within the range of values suggested for red wine grapes in warm climates by Kliewer and Dokoozlian (2005). In the rain-fed vines instead it seems that 1.5 m2 kg−1 of LA: Y is insufficient to maximize wine phenolics and lower crop load values (i.e. higher LA: Y) are then suggested in order to achieve the maximum potential under no irrigation conditions. Naor et al. (2002) also showed that for LA: Y above 1.7 m2 kg−1 there was not any clear increase in berry sugar content or in wine quality evaluated by sensory analysis.
Conclusions
The supplemental irrigation applied to vines when compared to rain-fed conditions had a detrimental effect on grape composition that was less pronounced under a large crop demand (around 16 t ha−1). This implies that irrigation should be used with caution and preferentially employed under high crop levels. A pre-veraison water shortage, compared to moderate water application throughout the season, decreased berry size at harvest and improved grape and wine composition only in one out of two experimental years. On the other hand, grape quality was negatively affected by high crop load only for leaf area to yield lower than 1.5 m2 kg−1. This crop load index could be then successfully used to manage vine balance, accordingly to vine water status.
References
Acevedo-Opazo C, Ortaga-Farías S, Fuentes S (2010) Effects of grapevine (Vitis vinifera L.) water status on water consumption, vegetative growth and grape quality: an irrigation scheduling application to achieve regulated deficit irrigation. Agric Water Manage 97:956–964
Allen RG, Pereira LS, Raes D, Smith M (1998) Crop evapotranspiration. Guidelines for computing crop water requirements. FAO Irrigation and Drainage paper No. 56. Rome, Italy
Bravdo B, Hepner Y, Loinger C, Cohen S, Tabacman H (1984) Effect of crop level on growth, yield and wine quality of a high yielding Carignane vineyard. Am J Enol Vitic 35:247–252
Edson CE, Howell GS, Flore JA (1995) Influence of crop load on photosynthesis and dry matter partitioning of Seyval grapevines. II. Seasonal changes in single leaf and whole vine photosynthesis. Am J Enol Vitic 46:469–477
Esteban MA, Villanueva MJ, Lissarrague J (1999) Effect of irrigation on changes in berry composition of Tempranillo during maturation. Sugars, organic acids, and mineral elements. Am J Enol Vitic 50:418–434
Esteban MA, Villanueva MJ, Lissarrague J (2002) Effect of irrigation on changes in the anthocyanin composition of the skin of cv. Tempranillo (Vitis vinifera L.) grape berries during ripening. J Sci Food Agric 81:409–420
Freeman BM, Kliewer WM (1983) Effect of irrigation, crop level and potassium fertilization on Carignane vines. II. Grape and wine quality. Am J Enol Vitic 34:197–206
Freeman BM, Lee TH, Turkington T (1979) Interaction of irrigation and pruning level on growth and yield of shiraz vines. Am J Enol Vitic 30:218–223
Girona J, Mata M, del Campo J, Arbones J, Bartra E, Marsal J (2006) The use of midday leaf water potential for scheduling deficit irrigation in vineyards. Irr Sci 24:115–127
Guidoni S, Allara P, Schubert A (2002) Effect of cluster thinning on berry skin anthocyanin composition of Vitis vinifera cv. Nebbiolo. Am J Enol Vitic 53:224–226
Hepner Y, Bravdo B (1985) Effect of crop level and drip irrigation scheduling on the potassium status of Cabernet Sauvignon and Carignane vines and its influence on must and wine composition and quality. Am J Enol Vitic 36:140–147
Iland PG, Coombe BG (1988) Malate, tartrate, potassium and sodium in flesh and skin of shiraz grapes during ripening: concentration and compartmentation. Am J Enol Vitic 39:71–76
Intrigliolo DS, Castel JR (2007) Evaluation of grapevine water status from trunk diameter variations. Irr Sci 26:49–59
Intrigliolo DS, Castel JR (2008) Effects of irrigation on the performance of grapevine cv. Tempranillo in Requena, Spain. Am J Enol Vitic 59:30–38
Intrigliolo DS, Castel JR (2010) Response of grapevine cv. ´Tempranillo` to timing and amount of irrigation: water relations, vine growth, yield and berry and wine composition. Irr Sci 28:113–125
Jackson DI, Lombard PB (1993) Environmental and management practices affecting grape composition and wine quality-A review. Am J Enol Vitic 44:409–430
Keller M, Mills LJ, Wample RL, Spayd S (2005) Cluster thinning effects on three deficit-irrigated Vitis vinifera cultivars. Am J Enol Vitic 56:91–103
Kliewer WM, Dokoozlian NK (2005) Leaf area/crop weight ratios of grapevines: influence on fruit composition and wine quality. Am J Enol Vitic 56:170–181
Lakso AN, Eissenstat DE (2005) Crop load-physiology, modeling and relationship to root growth in grapevines. In: Proceedings twelfth Australian wine industry technical conference. pp 38–44
Lakso AN, Dunst RM, Fendinger A (1999) Responses to drought of balance-pruned and minimally-pruned ‘Concord’ grapevines. Acta Hortic 493:103–107
Matthews MA, Nuzzo V (2007) Berry size and yield paradigms on grapes and wine quality. Acta Hortic 754:423–435
McCarthy MG (1997) The effect of transient water deficit on berry development of cv. Shiraz (Vitis vinifera L.). Aust J Grape Wine Res 3:102–108
Naor A (2008) Water stress assessment for irrigation scheduling of deciduous trees. Acta Hortic 792:467–481
Naor A, Gal Y, Bravdo B (1997) Crop load affects assimilation rate, stomatal conductance, stem water potential and water relations of field-grown Sauvignon blanc grapevines. J Exp Bot 48:1675–1680
Naor A, Gal Y, Bravdo B (2002) Shoot and cluster thinning influence vegetative growth, fruit yield, and wine quality of `Sauvignon blanc` grapevines. J Am Soc Hortic Sci 127:628–634
Ojeda H, Deloire A, Carbonneau A (2001) Influence of water deficits on grape berry growth. Vitis 40:141–145
Ough CS, Nagaoka R (1984) Effect of cluster thinning and vineyard yields on grape and wine composition and wine quality of Cabernet Sauvignon. Am J Enol Vitic 35:30–34
Poni S, Lakso AN, Turner JR, Melious RE (1994a) The effects of pre- and post-veraison water stress on growth and physiology of potted Pinot Noir grapevines at varying crop levels. Vitis 32:207–214
Poni S, Lakso AN, Turner JR, Melious RE (1994b) Interactions of crop level and late season water stress on growth and physiology of field-grown Concord grapevines. Am J Enol Vitic 45:252–258
Prajitna A, Dami IE, Steiner TE, Ferree DC, Scheerens JC, Schwartz SJ (2007) Influence of cluster thinning on phenolic composition, resveratrol, and antioxidant capacity in Chambourcin Wine. Am J Enol Vitic 58:346–350
Reynolds AG, Vanden Heuvel J (2009) Influence of grapevine training systems on vine growth and fruit composition: a review. Am J Enol Vitic 60:251–268
Reynolds AG, Edwards CG, Wardle DA, Webster DR, Dever M (1994) Shoot density affects ‘Riesling’ grapevines. I. Vine performance. J Am Soc Hortic Sci 119:874–880
Reynolds AG, Molek T, De Savigny C (2005) Timing of shoot thinning in Vitis vinifera: impacts on yield and fruit composition variables. Am J Enol Vitic 56:343–356
Reynolds AG, Lowrey WD, Tomek L, Hakimi J, de Savigny C (2007) Influence of irrigation on vine performance, fruti composition, and wine quality of Chardonnay in a cool, humid climate. Am J. Enol Vitic 58:217–228
Ribereau-Gayon P, Glories Y, Maujean A, Dubourdieu D (2000) Phenolic compounds. In: Ribereau-Gayon P, Glories Y, Maujean A, Dubourdieu D (eds) Handbook of enology, vol 2. The chemistry of wine stabilization and treatments, pp 129–187. John Wiley & Sons, New York
Roby G, Harbertson JF, Adams DA, Matthews M (2004) Berry size and wine water deficits as factors in winegrape composition: anthocyanins and tannins. Aust J Grape Wine Res 10:100–107
Romero EG, Muñoz GS, Ibáñez MDC (1993) Determination of organic acids in grape musts, wines and vinegars by high-performance liquid chromatography. J Chromat 655:111–117
Romero P, Fernández-Fernández JI, Martínez-Cutillas A (2010) Physiological thresholds for efficient regulated deficit-irrigation management in winegrapes grown under semiarid conditions. Am J Enol Vitic 61:300–312
Ruffner HP (1982) Metabolism of tartaric and malic acids in Vitis. A review-Part A1. Vitis 21:247–259
Salón JL, Chirivella C, Castel JR (2005) Response of Vitis Vinifera cv. Bobal to deficit irrigation in Requena, Spain. Water relations, yield and wine quality. Am J Enol Vitic 56:1–18
Shackel KA (2007) Water relations of woody perennial plant species. J Int Sci Vigne Vin 41:121–129
Smart RE, Coombe BG (1983) Water relations of grapevines. In: Kozlowski TT (ed) Water deficits and plant growth. Academic Press, New York, pp 137–196
Steel GD, Torrie JH (1980) Principles and procedures of statistics. McGraw-Hill, USA, pp 195–233
Trought CT, Tannock SJC (1996) Berry size and soluble solids variation within a bunch of grapes. In: Proceedings of the 4th international symposium on cool climate viticulture and enology. V, pp 70–72
Turner NC (1981) Techniques and experimental approaches for the measurement of plant water status. Plant Soil 58:339–366
Valdés ME, Moreno D, Gamero E, Uriarte D, Prieto MH, Manzano R, Picon J, Intrigliolo DS (2009) Effects of cluster thinning and irrigation amount on water relations, growth, yield and fruit and wine composition of tempranillo grapes in extremadura (Spain). J Int Sci Vigne Vin 43:67–76
Williams LE, Matthews MA (1990) Grapevine. In: Stewart BA, Nielsen DR (eds) Irrigation of agricultural crops, agronomy monograph no. 30. ASA-CSSA-SSSA, Madison, pp 1019–1055
Acknowledgments
This research was supported by funds from INIA-FEDER project RTA2008-00037-C04-01, from project Rideco-Consolider CSD2006-0067 and from a collaboration agreement with Caja Campo and the Fundación Lucio Gil de Fagoaga. We are grateful to the “Estación Experimental de Enología y Viticultura Requena” for the vinifications. Thanks are also due to C. Chirivella, E. Navarro, S. Cárcel, I. Yeves, and C. García for help in field determinations and analytical determinations and to Dr. E. Carbonell and J. Pérez for help with the statistical analysis of data.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Azam-Ali.
Rights and permissions
About this article
Cite this article
Intrigliolo, D.S., Castel, J.R. Interactive effects of deficit irrigation and shoot and cluster thinning on grapevine cv. Tempranillo. Water relations, vine performance and berry and wine composition. Irrig Sci 29, 443–454 (2011). https://doi.org/10.1007/s00271-010-0252-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00271-010-0252-2