
Abstract
A five-year experiment (2002–2006) was conducted to determine apple (cv ‘Golden Smoothee’) and pear (cv ‘Conference’) crop coefficients (Kc) using two large weighing lysimeters. Daily reference evapotranspiration (ETo) and crop evapotranspiration (ETc) were evaluated. Midday canopy light interception of both crops planted in hedgerows, 4 × 1.6 m, was determined on a weekly basis from bud-break until leaf fall from year 2002 (fourth after planting) to year 2006 of both plantations. Relationships between canopy light interception and calculated Kc (ETc/ETo) were evaluated from bud-break until harvest. There were differences in Kc values between apple and pear trees. When daily Kc values from bud-break until harvest were adjusted to hyperbolic functions each year, adjusted curves for pear trees were very similar regardless of year (maximum Kc around 1.0). In apple trees, the maximum values of Kc increased over time, from 0.49 in 2002 to 1.04 in 2006. Midday light interception in both apple and pear trees increased during the 5 years of experiment from 29.0 to 45.6% in apples and from 27.5 to 41.6% in pears in midsummer. Although there was a significant positive correlation between midday canopy light interception and Kc in apple and pear trees, in different times within a specific year, these relationships were different between crops. While the apple data fitted into the same equation regardless of the year, different equations were needed to fit the pear data in different years. This discrepancy may have been related to differences in the canopy properties between apple and pear trees. Pear canopies had higher porosity than apple canopies and thus improved light penetration. Apple trees were more vigorous and produced taller and denser canopies. Pear Kc values were greatly influenced by the evaporative demands of different years and consequently differences in midday canopy light interception did not adequately reflect the differences in Kc across the two species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Agricultural water management requires precise methods for estimating crop water requirements. Crop evapotranspiration can be accurately determined using weighing lysimeters. Although these are expensive and are not generally available to growers, relations derived from weighing lysimeter experiments could be used to develop more simple methods for estimating crop water use. For instance, standard crop coefficients (Kc) have traditionally been developed based on the ratio of crop evapotranspiration (ETc) to the potential evapotranspiration (ETo) (Doorenbos and Pruitt 1977; Allen et al. 1998; Allen and Pereira 2009). Since ETo is only a function of climatic parameters, using weather data and the estimated Kc, crop water use could be calculated as ETc = ETo × Kc. Even so, standard Kc values are general and may not be very representative of all commercial plantations for a specific crop type.
Accurate Kc values are difficult to obtain for fruit trees and grapevines. Kc should account for specific orchard conditions such as cultivar, orchard orientation, plant spacing, training system, and soil management among other factors. For a given orchard, however, it seems that canopy light interception is the main factor influencing the ETc/ETo ratio for peach trees (Johnson et al. 2000, 2002; Ayars et al. 2003), grapevine (Williams et al. 2003; Williams and Ayars 2005), and orange trees (Consoli et al. 2006). It therefore seems feasible to use canopy light interception values to adjust standard Kc data for specific orchard conditions (Johnson et al. 2000, 2002).
Preliminary Kc data from Girona et al. (2004) obtained for young lysimeter-grown pear trees were also positively correlated with midday canopy light interception. We were interested in evaluating the development of this relationship over several years of tree growth by comparing two species with different growth habits. A better understanding of the relationship between midday canopy light interception and crop coefficients may help develop a simple method for estimating crop water requirements under different orchard conditions that might then be extended to assist fruit growers in making irrigation management decisions.
The lysimeter-grown pear trees described by Girona et al. (2004) were located close to an adjacent weighing lysimeter planted in apple. This provided an excellent opportunity for testing variations in Kc in relation to canopy light interception for both pear and apple. We undertook a five-year study in which daily ETo and ETc were determined using these weighing lysimeters. Midday canopy light interception was determined on a weekly basis. The specific objectives of this research were as follows: (1) to measure crop coefficients for apple and pear trees using weighing lysimeters and (2) to compare the relationships between midday light interception and crop coefficients in apple and pear trees.
Materials and methods
Experimental plot
The experiment was conducted over 5 years (2002–2006) at two adjoining experimental apple (Malus x domestica Borkh. cv ‘Golden Smoothee’) and pear (Pyrus communis L. cv ‘Conference’) orchards. The orchards were located at the IRTA-Estaciò Experimental de Lleida (41°37′N; 0°52′E; 260 m a.s.l), Spain. Both orchards were planted in 1999 at a spacing of 4 × 1.6 m in a north–south row orientation. The apple trees were grafted onto ‘M-9’ rootstock and were trained to a modified central-leader system. The pear trees were grafted onto ‘M-A’ rootstock and trained to the same system as apple trees but pruned considering the different growth habits between apple and pear trees. Weather data were collected using an automated weather station located over grass 20 m apart from the orchards (see schematic presentation of the lysimetric station in Girona et al. (2004)) in order to avoid interactions between trees and the weather station. Mean annual precipitation during the five-year experiment was 324 mm. Harvest took place by mid-August in pear trees and by mid-September in apple trees.
Lysimeter-grown trees
We monitored three apple trees and three pear trees growing in large weighing lysimeters. These trees were located in the centres of their respective orchards as described by Girona et al. (2004). The lysimeter-grown trees were planted at the same time as similar trees grown in the experimental orchards, but the training system of apple trees was delayed by one year with respect to the pear trees. Both the lysimeter-grown apple and pear trees were managed according to commercial practices including ground cover elimination below the trees.
Weighing lysimeters
The weighing lysimeters measured 2 m wide, 4.8 m long and 1.7 m deep. The lysimeter tank, which was surrounded by a cement construction, was filled with the soil excavated from the lysimeter site. The original soil layers were preserved. One drainage (200 l) and two irrigation recipients (100 l each) were hung from the centre of the lysimetric tank [see schematic presentation of lysimeter transversal section in Girona et al. (2004)]. The weighing system consisted of four load cells with a unit capacity of 15 t (total weighing capacity 60 t). The sensitivity of the system was 0.5 kg, allowing the detection of a water consumption of 0.053 mm. Load cells were connected to a data logger (CR-10X; Campbell Scientific; Logan, Utah, USA) that continuously recorded the weight of each lysimetric tank throughout the experimental period.
Crop water requirements
The irrigation system consisted of an automated surface drip with 2 emitters per tree (4 l h−1 per emitter). There was a single pipeline, which was located close to the trunk of the trees. Each emitter was located 15 cm apart from the trunk. The number of emitters in the lysimeter trees was increased from 2 to 7 in order to avoid a hypothetical limitation in the volume of wetted soil affecting tree physiology and development. The trees were irrigated on a daily basis. The FAO-56 Penman–Monteith method was used to determine daily ETo. Daily apple and pear ETc was calculated from the weight differences between two consecutive days recorded in the lysimeter-tree unit. At midnight, the irrigation recipients were automatically refilled with the amount of water lost during the previous day plus 10% to prevent progressive soil drying. The water applied to the irrigation recipients was used to irrigate the lysimeter-tree units the following day. This irrigation routine was applied during the whole irrigation season. The irrigation season began at bud-break (beginning of April) and ended by late October (before leaf fall was completed).
Daily apple and pear Kcs were estimated from bud-break until the end of the irrigation season in each of the 5 years of the experiment, by applying the formula:
This Kc value included only the evaporation that occurred within the lysimeters and disregarded that occurring in the middle of the isle. Under semiarid conditions and using drip irrigation system with pipelines located underneath the trees (wetted soil in shade), soil evaporation of the middle part of the isle tends to be affected by rain, and thus its effect tends to be negligible on the whole soil water balance if rainy days were not considered for the calculation of Kc. Therefore, this estimated Kc could be only applied under such conditions. However, they can be converted into FAO-56 parameters by using the basal crop factor (Kcb). After 2007, we did some studies while covering the lysimeter soil surface with plastic. During the summer, soil evaporation from the lysimeter was, on average, 5% of tree transpiration. Therefore, Kcb = 0.95 × Kc. This justifies the elimination of rainy days for the calculation of seasonal trends in Kc.
Canopy light interception
Midday light interception by the lysimeter-tree unit was determined with a linear ceptometer (probe length 80 cm; Accupar Linear PAR; Decagon Devices, Pullman, WA, USA). Measurements were taken on a weekly basis, at solar noon ±30 min on cloudless days, from bud-break until leaf fall in each of the 5 years of the experiment. For each lysimeter, 64 measurements were taken at fixed positions (32 on each side) within the space assigned to the lysimeter trees (4 × 1.6 m). The ceptometer was placed in a horizontal position at ground level and was placed perpendicular to the trees. On each occasion, two further measurements were made at an open site with no interference from the canopy. The percentage of PAR intercepted by the lysimeter-grown trees was estimated as:
Canopy properties
Tree size, in terms of height and width, was estimated with measuring tape each year at midseason (10 July). Leaf area index (LAI) was measured during 2003 by collecting all the leaves trapped by nets covering all trees in the lysimeters before leaf shedding. For the other years, LAI was estimated from light inversion equations (Campbell 1986) that have been parametrically adjusted for the known LAIs of 2003. The LAI values correspond to those of 10 July for each year because on this day midday PAR interception was measured. Noon canopy porosity was estimated for each year at midseason from tree size, row orientation, and geographical parameters following Oyarzun et al. (2007).
Analysis of data
Relationships between the percentage of PAR intercepted by the lysimeter-grown trees at solar noon and the mean daily Kc obtained 3 days before and after the PAR measurements were evaluated in each of the five experimental years. Since rainfall increased the amount of soil evaporation, when significant rainfall was recorded 3 days before or after the PAR measurements, the corresponding mean Kc value was discarded. The equation that gave the best fit for the observed relationships was selected, and the R 2 values were analysed in order to determine how much variation in daily Kc was explained by the percentage of PAR intercepted. Statistical analyses were performed using the SAS software package (SAS Institute, Cary, NC, USA). Statistical significance was established at P ≤ 0.05.
TableCurve® Software was used to adjust hyperbolic functions regarding the response of Kc to time form bud-break until harvest.
Results
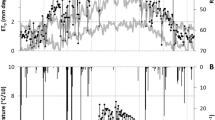
The seasonal patterns of daily ETo obtained from the weather station adjacent to the experimental orchards were similar from one year to another (Fig. 1). Daily ETo values increased with time until they reached a peak of about 6.5–7.0 mm day−1 midway through the year. Thereafter, daily ETo values decreased with time following a similar pattern to their previous increase.

Seasonal variations in daily reference evapotranspiration (ETo) and daily crop evapotranspiration (ETc) for lysimeter-grown apple and pear trees from 2002–2006. Part of the pear ETc data for 2006 were excluded because trees were undergoing a period of water stress
Daily ETc values increased with time from bud-break until full canopy development (Fig. 1). Daily ETc values were then maintained until harvest. After fruit harvest, daily ETc decreased with time until the end of the irrigation season. In pear trees, the seasonal patterns of daily ETc were quite consistent from year to year (Fig. 1). However, there were noticeable differences in maximum values of apple ETc from one year to another (Fig. 1).
Irrespective of the crop, seasonal patterns of daily Kc followed a similar trend from year to year (Fig. 2). At bud-break, initial Kc values were about 0.2 for both apple and pear trees (Fig. 2). They increased with time until reaching maximum values some time before harvest (Fig. 2). When daily Kc from bud-break until harvest was adjusted to an hyperbolic curve, the adjusted curves for pear trees were very similar from one year to another, but those for apple trees showed differences between years (Fig. 3). For pear trees, when five-year data were adjusted to a single hyperbolic curve, mean Kc reached maximum values of about 0.9 with the exception of 2006 when partial blocking of emitters caused water stress in trees. The average midday stem water potential for well-watered orchard pear trees was −0.9 MPa while that of the stressed lysimeter pear trees was −1.4 MPa. The Kc values for the stressed pear trees were excluded from Figs. 1,2, and 3. For apple trees, however, maximum values of Kc were observed at about 0.6, 0.7, 0.9, and 1.0 in 2003, 2004, 2005, and 2006, respectively (Fig. 3). Irrespective of the crop, Kc values exhibited an immediate reduction (Fig. 2) after fruit sink removal at harvest. Fruit removal decreases tree water use (Reyes et al. 2006; Marsal et al. 2008). Near the end of the irrigation season, final Kc values were about 0.4 for both apple and pear trees (Fig. 2).

Daily patterns of crop coefficients (Kc) for lysimeter-grown apple and pear trees from bud-break until the end of the irrigation season throughout the experimental period (2002–2006). The Kc values were not considered when significant rainfall occurred. The Kc data were adjusted to a seven-day moving average. Part of the pear Kc data for 2006 were excluded because trees were undergoing a period of water stress

Daily patterns of crop coefficients (Kc) for lysimeter-grown apple and pear trees from bud-break until harvest throughout the experimental period (2002–2006). For apple trees, Kc data were adjusted to a quadratic curve for each experimental year. For pear trees, mean Kc data from the whole experimental period were adjusted to a single quadratic curve for better visibility of data
Both apple and pear trees exhibited a progressive increase in the percentage of PAR intercepted at solar noon over the course of the five-year experiment (Fig. 4). In the first year (2002), apple trees exhibited lower values of intercepted PAR than pear trees (Fig. 4). However, both apple and pear trees had similar values in the following years (Fig. 4).

Seasonal variations in the percentage of photosynthetically active radiation (PAR) intercepted at solar noon by individual lysimeter-grown apple and pear trees from 2002–2006
There were positive correlations between the percentage of PAR intercepted at solar noon and daily Kc (Fig. 5). Irrespective of the crop, Kc increased with increases in the percentage of PAR intercepted at solar noon. When the range of data was sufficient to establish relationships between the two variables, the best fit was obtained using exponential equations (Fig. 5). There were significant differences in the adjusted equations between apple and pear trees. For a given canopy light interception, pear trees presented higher Kc values than apple trees. Moreover, there were clear differences in the adjusted equations between the apple and pear trees from year to year. The apple data fitted the same equation irrespective of the year. However, there were significant differences in the established relationships between the percentage of PAR intercepted at solar noon and pear daily Kc from one year to another (Fig. 5).

Relationships between percentages of photosynthetically active radiation (PAR) intercepted at solar noon and daily crop coefficients (Kc) for individual lysimeter-grown apple and pear trees from bud-break until harvest. For pear trees, the relationships were fitted into exponential equations for 3 years of evaluation. For apple trees, the relationship was fitted to a single exponential equation for all the years of evaluation. Each Kc value represents the average Kc calculated 3 days before and after the PAR measurements
Apple canopies had different characteristics compared to pear canopies (Table 1). Apple trees tended to be taller, less porous, with higher LAI, and with higher percentage of PAR interception at noon than those of pear trees (Table 1). Estimates of apple Kc indicated progressive increases over the experimental years, and Kc in midseason (Kcmid) was linearly related to the percentage of intercepted PAR (Kcmid = 0.0331·PAR − 0.481; R 2 = 0.97 P < 0.001) (Table 2). This was not the case in pear trees where maximum Kc was observed during 2003, which was characterised with high air temperatures and with ETo being higher than any other year (Table 2). Kcmid in pear trees was significantly related to ETo of each year (Kcmid = 0.03212 ETo − 0.4518; R 2 = 0.93 P < 0.001) (Table 2).
Discussion
Water use by fruit trees has been related to canopy size (Fereres et al. 1982). In this study, there were positive correlations between canopy size (as defined by the percentage of PAR intercepted at solar noon) and tree water use (as related to daily Kc) (Fig. 5). Nevertheless, the relationship between midday canopy light interception and Kc differed between apple and pear trees. While apple data fitted into the same equation irrespective of the year, pear data fitted into different equations for each year of evaluation (Fig. 5). This differential performance reflected significant differences in Kc values from one year to another between apple and pear trees (Fig. 3). Consequently, in pear trees, the progressive increase in midday canopy light interception from year to year (Fig. 4) was not accompanied by a significant increment in Kc (Fig. 3). However, in apple trees, there was synchronisation between the increments in canopy light interception and Kc from year to year (Figs. 3 and 4).
It has been reported that training system has a significant effect on tree water relations (Chootummatat et al. 1990; Li et al. 2002). The different growth habits between apple and pear trees in our study may therefore partially explain the discrepancy between the respective Kc values (Fig. 3). Pear canopies produced more sun flecks underneath (visual observation) and were more porous than apple canopies as reported in Table 1. On the other hand, apple trees were more vigorous, taller, had higher LAI, and a higher percentage of PAR interception than pear canopies. It is therefore expected that an average pear leaf received more light than an average apple leaf. Therefore, it is expected that pear Kc be higher than that of apple per unit of LAI. In addition and as a consequence of these different canopy properties, midday canopy light interception was not well linked to differences in Kc between species. On the other hand, it is also likely that midday canopy light interception may not have reflected differences in tree height between apple and pear trees. This would be in agreement with the findings of Jackson (2003) who reported that identical levels of light interception can be achieved with very different tree forms. For a given midday canopy light interception, apple trees may therefore have had a lower canopy volume and water demand than pear trees (Figs. 3 and 4). Using our results, it would be difficult to accurately estimate Kc for different tree types if the same midday canopy light interception technique were employed. In order to improve predictions of Kc based on midday light interception, adjustments for canopy structure may also be needed (Trout and Gartung 2006). In fruit trees, these adjustments may include information related to the training system, such as tree height or canopy spread, and could be used in modelling as a tool (Johnson et al. 2004). Another approach that may help compare water use in trees with different shapes may be the use of more sets of canopy light interception readings during the day (Goodwin et al. 2006).
However, the above considerations may not fully account for the year-to-year variations in pear Kcmid. The values were more sensitive to ETo than to changes in tree size or percentage of intercepted PAR. The high pear Kcmid sensitivity to ETo or daily vapour pressure deficit (VPD) (Table 2) may have masked any possible relationship between Kcmid and intercepted PAR. The latter relationship did occur for apple (Table 2). The hottest conditions were observed during 2003 (Table 2) when pear trees were small (Table 1). Yet, the pear Kcmid was the highest of all years indicating how the tree was influenced more by temperature than by the intercepted PAR. Besides the above-mentioned canopy factors, higher Kc for high ETo or high VPD found in pear trees may also be related to lower stomatal sensitivity to VPD when compared to apple stomata (Higgins et al. 1992) (Table 2). Low reactive stomata to high VPD would make the system more susceptible to changes in evaporative demand as it seems to have happened for pear especially in the hot conditions of 2003 (Table 2).
From these results, it seems that using midday canopy light interception by itself would not be an appropriate technique for scheduling irrigation of fruit trees whose shape in hedgerows is predominantly vertical. It has already been well established that vertical tree forms require more sets of irradiance readings to be taken at ground level within the shaded area below the canopy throughout the day in order to estimate total daily light interception (Wünsche et al. 1995).
In summary, this research has provided Kc values for apple and pear trees that can be useful for orchards in semiarid areas similar to our conditions and with drip irrigation system being used. Year-to-year changes in apple Kc can be approximated by using the crop intercepted PAR at noon, while in pear this method is not recommended. In general, Kc in pear seemed higher than in apple for a given noon intercepted PAR. Several explanations have been offered in the manuscript to account for this. Differences in canopy porosity and stomatal sensitivity to VPD might have been the most relevant explanations.
References
Allen RG, Pereira LS (2009) Estimating crop coefficients from fraction of ground cover and height. Irri Sci 28:17–34
Allen RG, Pereira LS, Raes D, Smith M (1998) Crop evapotranspiration. Guidelines for computing crop water requirements. FAO irrigation and drainage paper no 56, 300 pp
Ayars JE, Johnson RS, Phene CJ, Trout TJ, Clark DA, Mead RM (2003) Water use by drip-irrigated late-season peaches. Irrig Sci 22:187–194
Campbell GS (1986) Extinction coefficients for radiation in plant canopies calculated using an ellipsoidal inclination angle distribution. Agric For Meteorol 36:317–321
Chootummatat V, Turner DW, Cripps JEL (1990) Water use of plum trees (Prunus salicina) trained to four canopy arrangements. Sci Hortic 43:255–271
Consoli S, O’Connell N, Snyder R (2006) Measurement of light interception by navel orange orchard canopies: case study of Lindsay, California. J Irrig Drain Eng ASCE 132:9–20
Doorenbos J, Pruitt WO (1977) Crop water requirements. FAO irrigation and drainage, paper no 24, 194 pp
Fereres E, Martinich DA, Aldrich TM, Castel JR, Holzapfel E, Schulbach H (1982) Drip irrigation saves money in young almond orchards. Calif Agric 36:12–13
Girona J, Marsal J, Mata M, del Campo J (2004) Pear crop coefficients obtained in a large weighing lysimeter. Acta Hortic 664:277–281
Goodwin I, Whitfield DM, Connor DJ (2006) Effects of tree size on water use of peach (Prunus persica L. Batsch). Irrig Sci 24:59–68
Higgins SS, Larsen FE, Bendel RB, Radamaker GK, Bassman JH, Bidlake WR, Al Wir A (1992) Comparative gas exchange characteristics of potted, glasshouse-grown almond, apple, fig, grape, olive, peach and Asian pear. Sci Hortic 52:313–329
Jackson JE (2003) Biology of apple and pears. Cambridge University Press, United Kingdom
Johnson RS, Ayars J, Trout T, Mead R, Phene C (2000) Crop coefficients for mature peach trees are well correlated with midday canopy light interception. Acta Hortic 537:455–460
Johnson RS, Ayars J, Hsiao T (2002) Modelling young peach tree evapotranspiration. Acta Hortic 584:107–113
Johnson RS, Ayars J, Hsiao T (2004) Improving a model for predicting peach tree evapotranspiration. Acta Hortic 664:341–346
Li F, Cohen S, Naor A, Shaozong K, Erez A (2002) Studies of canopy structure and water use of apple trees on three rootstocks. Agric Water Manag 55:1–14
Marsal J, Mata M, Arbones A, del Campo J, Girona J, Lopez G (2008) Factors involved in alleviating water stress by partial canopy removal in pear trees. Tree Physiol 28:1375–1382
Oyarzun RA, Stökle CO, Whiting MD (2007) A simple approach to modeling radiation interception by fruit-tree orchards. Agric For Meteorol 142:12–24
Reyes VM, Girona J, Marsal J (2006) Effect of late Spring defruiting on net CO2 exchange and leaf area development in apple tree canopies. J Hortic Sci Biotechnol 81(4):575–582
Trout TJ, Gartung JL (2006) Use of crop canopy size to estimate crop coefficient for vegetable crops. Environmental and water resources institute world congress proceedings, Omaha, NE, USA
Williams LE, Ayars JE (2005) Grapevine water use and the crop coefficient are linear functions of the shaded area measured beneath the canopy. Agric For Meteorol 132:201–211
Williams LE, Phene CJ, Grimes DW, Trout TJ (2003) Water use of mature Thompson seedless grapevines in California. Irrig Sci 22:11–18
Wünsche JN, Lakso AN, Robinson TL (1995) Comparison of 4 methods for estimating total light interception by apple-trees of varying forms. HortScience 30:272–276
Acknowledgments
The cost of the weighing lysimeters was covered by INIA. We acknowledge the support of CSD2006-00067 grant of CONSOLIDER-INGENIO 2010. The authors are grateful to two anonymous reviewers for their constructive comments and to Prof. M. H. Behboudian, from Massey University, New Zealand, and Prof. E. Fereres, from University of Cordoba, Spain, for their revision of the last version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Asam-Ali.
Rights and permissions
About this article
Cite this article
Girona, J., del Campo, J., Mata, M. et al. A comparative study of apple and pear tree water consumption measured with two weighing lysimeters. Irrig Sci 29, 55–63 (2011). https://doi.org/10.1007/s00271-010-0217-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00271-010-0217-5