
Abstract
The effects of deficit irrigation (DI) and partial rootzone drying (PRD) on apple (Malus domestica Borkh. Cv. ‘Fuji’) yield, fruit size, and quality were evaluated from 2001 to 2003 in the semi-arid climate of Washington State. PRD and DI were applied from about 40 days after full bloom until just before (2001, 2002) or after (2003) harvest and compared to a control irrigation (CI). Irrigation was applied once a week using two micro-sprinklers per tree. Soil-water content in CI was maintained above 80% of field capacity using micro-sprinklers on both sides of a tree. The DI and PRD were irrigated at about 50% (2001–2002) and 60% (2003) of the CI, but differed in placement of irrigation. For DI both micro-sprinklers were operated whereas PRD was irrigated using only one micro-sprinkler wetting half the rootzone compared to CI and DI. Wetting/drying sides of PRD trees were alternated every 2–4 weeks (2001, 2002) or when soil-water content on the drying side had reached a threshold value (2003). Seasonal (1 May–31 October) potential evapotranspiration (ET0) was 967, 1002, and 1005 mm for 2001, 2002, and 2003, and rainfall totaled 58, 39, and 21 mm, respectively. Irrigation amounts applied were 596, 839, and 685 mm in the CI; 374, 763, and 575 mm in the DI; and 337, 684, and 513 mm in the PRD for the 2001, 2002, and 2003 seasons. Higher irrigation volumes in 2002 were due to excessive (177–324 mm) irrigations after harvest. No significant differences were found in yield and fruit size among treatments in 2001 and 2003. In 2002, DI had significantly lower yield than CI, while the yield of PRD did not differ from CI and DI. Fruit from DI and PRD were firmer and had higher concentrations of soluble solids than fruit from CI, both at harvest and following short-term storage at 20°C, but differences to CI were significant in 2002 only. Treatment effects on fruit titratable acidity were inconsistent. Additional water was preserved in the soil profile under PRD compared to DI in 2001 and 2003, but no statistical differences were found between PRD and DI in 2002. Approximately 45–50% of irrigation water was saved by implementing newly developed DI and PRD irrigation strategies without any significant impact on fruit yield and size with PRD. However, apple yield was reduced by DI compared to CI in the second year.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Irrigated agriculture in the Pacific Northwest region of the United States is facing a number of political, economic, and environmental challenges. The success and survival of many agricultural and horticultural industries in this semi-arid region depends on the continued supply of water for irrigation. However, good quality water is a scarce resource and competition between resource users will continue to increase. With increasing water demands from cities and recreational and environmental groups, less of this finite resource will be available for agricultural use. Clearly, new approaches for irrigation management are required that will reduce both water consumption and the detrimental environmental effects of current agricultural practices. While irrigation strategies such as regulated deficit irrigation (RDI) (Chalmers et al. 1981) have been used successfully with a number of crops and can significantly reduce water use in crops such as peaches [Prunus persica L. Batsch] (Mitchell and Chalmers 1982; Li et al. 1989; Boland et al. 1993), pears [Pyrus spp.] (Mitchell et al. 1989; Caspari et al. 1994; Marsal et al. 2002), and olives [Olea europaea L.] (Alegre et al. 1997; Goldhamer 1997), they have not been successful with apples due to a negative impact on fruit size and yield. In fact, many studies have shown that water deficits reduce final fruit size in apples, irrespective of timing (Landsberg and Jones 1981; Lötter et al. 1985; Ebel et al. 1993, 1995, 2001; Mpelasoka et al. 2000, 2001). Partial rootzone drying (PRD) is a new deficit irrigation strategy that has been recently developed for grapevines (Vitis vinifera L.) in Australia (Dry et al. 1996, 2000a, b, 2001; Loveys et al. 1997, 1998; Dry and Loveys 1998) and may allow the use of deficit irrigation (DI) on crops where other deficit strategies such as RDI lead to negative outcomes. With PRD, irrigation is withheld from a part of a plant’s rootzone while the remaining part is kept well watered. Briefly, the proposed physiological mechanism of PRD is that roots in drying soil synthesize a hormonal signal (abscisic acid, ABA), which is transported to the shoots, indicating a developing soil-water deficit. In the leaves, ABA induces partial stomatal closure, which increases water-use efficiency. Increases in ABA concentration in response to soil drying and the effects on growth and gas exchange are well documented (Davies and Zhang 1991; Tardieu et al. 1992a,b), and have also been shown for grapevines (Dry et al. 1996). However, as the remaining part of the rootzone is kept well watered with PRD, the effect on plant water potential is minimal. In order to maintain the ABA signal, irrigation is alternately applied to each side of the rootzone, allowing the wet side to dry while the dry side is wetted. Such application of PRD to grapevines has resulted in water savings of up to 50% with significant reductions in vegetative vigor and improved fruit quality, but without loss of yield (Dry et al. 1996; Loveys et al. 1998). The potential to increase water use efficiency in agriculture through the exploitation of such chemical signaling has recently been reviewed by Davies et al. (2002).
Since 1995, a number of trials with PRD on apples in the Marlborough region of New Zealand, and one preliminary trial in Washington State have been conducted. In the New Zealand experiments with ‘Gala’, ‘Fuji’, and ‘Braeburn’ apples, seasonal irrigation input was reduced by 30–50% without loss in fruit size or yield (Caspari et al. 2003; Caspari and Neal, unpublished data). Similarly, PRD applied for the final 7 weeks prior to harvest did not reduce fruit size and yield of ‘Golden Delicious’ apple grown near Prosser, Washington, while saving 50% of irrigation water over this same period (Caspari and Lang, unpublished data).
Many studies have compared one regime or another of deficit irrigation and a well-watered control, but only a few studies have included more than one type of deficit irrigation. During the 1999–2001 growing seasons, three different regimes of deficit irrigation of ‘Braeburn’ apples were compared to a well-watered control in the Marlborough region of New Zealand (Caspari et al. 2003). All deficit treatments were irrigated at 50% of the control but differed in respect of placement and/or timing of irrigation. Fruit size and yield were reduced by omitting every other irrigation and by applying 50% of the water to the entire planting area as compared to the control, but fruit size and yield were not affected by PRD. There was no significant treatment effect on soluble solids, firmness, color, starch pattern index, and the development of fruit disorders.
The temperate climate of New Zealand differs greatly from the semi-arid conditions of eastern Washington State. The objective of this study was to determine the impact of DI and PRD on apple yield, mean fruit size, fruit quality, and orchard water requirements in a semi-arid region where annual precipitation may provide less than 10% of orchard water use.
Materials and methods
Experimental site and irrigation regimes
This study was conducted from 2001 to 2003 at Washington State University’s Roza research site near Prosser in southeastern Washington State. At the beginning of the 2001 growing season, a block of 6-year-old ‘Fuji’ apple trees containing 120 trees on a 2.74 m × 4.88 m spacing was converted from furrow to micro-sprinkler irrigation. The micro-sprinklers were placed in the tree rows midway between trees. Micro-sprinklers had a circular wetting pattern with a diameter of 2.7 m. This configuration isolated the irrigation to each side of a tree to allow for PRD. Water supply pipes were installed to create three irrigation regimes: (1) control irrigation (CI), (2) DI, and (3) PRD in a completely randomized block (CRB) design with four replications. Each plot consisted of at least nine trees (three trees per row in three adjacent rows). Generally, data were collected from the central row while the outside trees acted as guards. Soil management consisted of a permanent grass cover crop with a ∼2.0 m wide herbicide strip under the tree row. At full leaf area development the tree canopy covered 65–70% of ground area.
The soil-water content in the CI plots was maintained close to field capacity. Irrigation was applied once a week. The irrigation volume applied in the control was only 60–70% of estimated crop evapotranspiration (ETc) for an apple orchard without a cover crop (James et al. 1989), as calculated by the Public Agricultural Weather System (PAWS). The meteorological data for ET calculations were collected from a PAWS weather station (http://frost.Prosser.wsu.edu/) located at the research orchard. In the CI blocks, irrigation was applied using two sprinklers per tree (one on either side of the trunk). Early in the season, PRD and DI treatments were irrigated like the control. Starting in early June, about 40 days after full bloom, PRD and DI plots were irrigated as described below until late September (2001) or early (2002) or mid October (2003) when all plots were returned to field capacity for winter dormancy.
The irrigation approach for PRD and DI differed slightly between years. In 2001 and 2002, both PRD and DI were irrigated at a fixed ratio (50%) of the volume applied to the control, but differed in placement of irrigation water. For the PRD blocks, only one sprinkler per tree was operated for the same length of time as the CI. The irrigated and dry sides of the PRD trees were alternated every three−four weeks in 2001, and every two weeks in 2002. In the DI blocks, sprinklers on both sides of the tree were operated for half the time of the control during 2001 and 2002. However, measurements of sprinkler outputs in 2002 indicated that the output was about 8% higher in DI than in PRD and CI, and DI application times were reduced accordingly during 2003. Further, results from the 2002 season indicated that the fixed ratio of 50% was insufficient to maintain one side of the PRD rootzone ‘wet’ (see later). Consequently, in an attempt to maintain one side of the PRD rootzone close to field capacity at all times, irrigation volumes for PRD were adjusted as follows: from early June to mid July, PRD and DI were irrigated at 50% of control. On 18 July, 3 days before the second switch of the wet/dry sides, an additional irrigation was applied to ensure that the soil moisture of the wet side was close to field capacity, and the process was repeated on 7 August, 4 days prior to the third switch. Finally, on 16 and 23 September (prior to and coinciding with the final switch of the season), PRD was irrigated at 87% of control. At all other times, PRD was irrigated at 50% of the CI’s volume and at the same time as CI. The DI volume was equally adjusted to match the irrigation input and timing of the PRD at all times. Averaged over the treatment period in 2003, the PRD and DI regime received 60% of the CI’s volume (compared to 50% for 2001 and 2002). Cumulative values for irrigation, potential evapotranspiration (ET0), ETc, and precipitation for the 2001, 2002, and 2003 seasons are presented in Fig. 1.

Accumulated reference evapotranspiration (ET0), predicted water loss of an apple orchard without cover crop (ETc), irrigation volumes applied in the CI, DI, and PRD regimes, and rainfall during 2001, 2002, and 2003 for the Roza Research Station near Prosser, WA. Arrows at the top indicate the period of differential irrigation
Soil-water content and soil-water potential
Soil-water content was monitored on a weekly basis using the neutron probe method (Gardner 1986). Neutron probe access tubes were located midway between the micro-sprinklers and the trees. One access tube was installed in each experimental unit with the exception of the PRD plots where two access tubes were installed to measure soil-water content on the irrigated and non-irrigated sides of a tree’s rootzone. Access tubes were inserted to a depth of 0.9–1.2 m depending on the soil depth encountered. Soil-water content was measured at 0.15 m intervals and approximately 2–3 days after each weekly irrigation.
Soil-water potential (ψs) was measured at 0.15, 0.45, and 0.75 m soil depths in one plot per treatment using Watermark sensors connected to a Hansen data logger. Data were recorded every 8 h, and instantaneous values were also recorded manually once a week while taking neutron probe readings. Due to a computer problem, the data recorded by the data logger during 2001 were lost.
The soil used in this study was classified as Warden silt loam soil (coarse, silty, mixed, mesic, and Xerollic Camborthid), developed from lacustrine sediment and has a mantle of loess parent material with a field capacity of ∼0.27 m3 m−3 and a permanent wilting point of ∼0.08 m3 m−3 .
Fruit growth, yield, and fruit quality
Fruit size was monitored weekly using a Cranston diameter gauge on ten selected fruits per tree, one tree per plot. The same ten fruits were measured throughout the growing season.
All fruits were harvested from each experimental tree on October 3, 2001, October 17, 2002, and October 14, 2003. In 2001, two of the four replicates in the DI treatment were carrying a much higher crop load than any of the other trees. To further evaluate the influence of crop load and irrigation on fruit size, an additional four to five trees per treatment were harvested using the guard trees. For each tree, fruits were sorted by size with a Cranston diameter gauge and each size group was counted and weighed. In all the years, a sub-sample of fruits was taken to the laboratory for further analysis. Fruit maturity was assessed 1 day after harvest, and after 14 (2001), 7 (2002), or 10 days (2003) at room temperature (20°C). Measurements were made of fruit firmness using the Fruit Texture Analyzer (Güss), soluble solid concentration (SSC) using a digital refractometer (Atago), starch index (0–6 scale for ‘Fuji’), density, titratable acidity, percentage of red color, and internal ethylene concentration using a gas chromatograph (Hewlett-Packard GC 5830 with a PLOT column). Ten apples per tree were used for each measurement and time. Six apples stored for 14 days in respiration chambers at room temperature (20°C) in 2001 were monitored for carbon dioxide (CO2) and ethylene (C2H4) evolution.
Results
Irrigation
Seasonal (May 1–Oct 31) reference ET0 and ETc were 967 and 758 mm in 2001, 1002 and 792 mm for 2002, and 1005 and 811 mm for 2003 (Fig. 1). Seasonal precipitation totaled 59, 39, and 21 mm, respectively. Cumulative irrigation inputs for 2001, 2002, and 2003 were 596, 839, and 685 mm for CI, 374, 753, and 575 mm for DI, and 337, 684, and 513 mm for PRD.
Soil-water content
All three irrigation regimes showed a small drop in soil-water content in May (Fig. 2). However, the larger decline in soil-water contents in 2001 was due to a temporary hold of water deliveries to conserve reservoir water for use later in the growing season due to drought conditions in Washington State during 2001. Furthermore, due to the drought, water delivery ended earlier than normal in late September 2001. Consequently, soil-water content was raised back up to field capacity earlier in 2001 than in 2002 and 2003 to prepare for the dormant season (Fig. 2). With the exception of these drought-induced changes, the soil-water content in the wetted zone for the CI treatment was kept near field capacity (0.24–0.26 m3 m−3) during the first two growing seasons. In 2003, soil-water content in CI was maintained a little lower at about 80–90% of field capacity from mid June to mid August before being raised to field capacity for the remainder of the season.

Effect of irrigation regime on the mean soil moisture content of a ‘Fuji’ apple orchard near Prosser, WA. Solid line at the top indicates the period of differential irrigation. Asterisks at the top of each graph indicate irrigation events, where large-sized asterisks indicate a switch of the irrigated side for the PRD treatment. Vertical bars indicate LSD (P≤0.05)
Following the start of irrigation treatments (June 4, 2001; June 11, 2002; June 3, 2003), both the DI and PRD showed a consistent decrease in soil-water content (Fig. 2). However, soil-water depletion was greater with DI than PRD in all years, although differences were comparatively small in 2002. Soil-water depletion in the wetted profile at the end of the treatment periods for DI and PRD was 133 and 80 mm in 2001, 123 and 113 mm in 2002, and 125 and 55 mm in 2003. It is worth noting that the actual irrigation volumes applied, as determined by measuring sprinkler outputs, were slightly higher in DI than PRD, and the DI received an additional 27 and 32 mm during the periods of differential irrigation in 2001 and 2002, respectively. With the adjustment in application time for DI during 2003, the difference was reduced to only 5 mm. Thus, the DI differed from PRD in apparent water use, defined here as the sum of soil-water depletion and irrigation input, by 80, 42, and 75 mm in 2001, 2002, and 2003.
Differences between DI and PRD as well as year-to-year differences for the PRD are also evident from the depth-wise pattern of soil-water depletion (Fig. 3). In 2001, soil-water content at 0.3 m was close to that of CI, i.e. close to field capacity, in at least one part of the PRD rootzone throughout the season. However, soil moisture content on the ‘dry’ PRD side declined rapidly after a switch, and reached lower levels than the DI before recovering to near-CI levels following the next switch. Likewise, soil moisture at 0.6 m was close to that of CI until mid August when it started to decline. Soil moisture content at 0.9 m for the PRD tended to decline all through the season with only a small increase in response to switches. This indicates that the irrigation volume applied was insufficient to re-wet the lower part of the soil profile. Soil moisture content declined more with DI than with PRD and was lower at all layers with DI from mid July until re-watering in late September.

Effect of irrigation regime on the mean soil moisture content at 0.3 (top), 0.6 (middle), and 0.9 m (bottom) of a ‘Fuji’ apple orchard near Prosser, WA during the 2001, 2002, and 2003 seasons. Soil moisture content is shown for both sides (North, South) of the PRD regime. Solid line at the top indicates the period of differential irrigation. Asterisks at the top of each graph indicate irrigation events, where large-sized asterisks indicate a switch of the irrigated side for the PRD treatment
Despite higher irrigation volumes applied in 2002 compared to 2001, soil-water content for both PRD and DI reached levels lower than in 2001, and there was little difference in soil-water depletion between the two regimes (Fig. 3). While the effect of switching is evident at 0.30 m for the PRD, the soil-water content continued to decline over time. Not enough water was applied in the PRD to cause any significant increase in soil moisture at 0.6 and 0.9 m following a switch. Consequently, the pattern of soil moisture depletion on both sides of the PRD mirrored that of DI. In contrast, the change in irrigation strategy for the PRD regime in 2003 from a fixed 50% ratio to a ‘response-type’ approach again led to pronounced differences in soil-water depletion between PRD and DI. As in 2001, soil moisture at 0.3 m in one part of the PRD rootzone was kept close to the CI for most of the season. In addition, sufficient irrigation was applied to maintain soil-water content at 0.6 and 0.9 m higher than in 2001 and 2002, particularly late in the season. In contrast, and despite the increased irrigation volume, the soil moisture depletion in the DI was similar to that of the previous two years with late-season values for 0.6 and 0.9 m being even lower than in 2001, but slightly higher than in 2002.
Soil-water potential
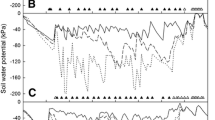
Measurements in one plot per treatment confirm that switching the irrigated and non-irrigated sides of PRD resulted in a substantial drop in ψs on the dry side with a concomitant increase on the wet side (Figs. 4, 5). Fluctuations were largest at 0.15 m soil depth, intermediate at 0.45 m, and least at 0.9 m in all years. In 2001, ψs on at least one side of the PRD rootzone was always higher (less negative) than that of CI at 0.15 m, and higher than DI for all depths (Fig. 4). Soil-water potentials at 0.45 and 0.75 m were lower in the DI than in even the dry side of the PRD from 2 August 2001 (day of year [DOY] 214) until after the soil profile had been re-wetted by the final irrigation on 24 September.

Effect of irrigation regime on the soil-water potential at 0.15 (top), 0.45 (middle), and 0.75 m (bottom) of a ‘Fuji’ apple orchard near Prosser, WA during the 2001 season. Soil-water potential is shown for both sides (North and South) of the PRD regime. Solid line at the top indicates the period of differential irrigation. Asterisks at the top of each graph indicate irrigation events, where large-sized asterisks indicate a switch of the irrigated side for the PRD treatment

Effect of irrigation regime on the soil-water potential at 0.15 (top), 0.45 (middle), and 0.75 m (bottom) of a ‘Fuji’ apple orchard near Prosser, WA during the 2002 and 2003 seasons. For PRD only the maximum (least negative) soil-water potential is shown. Lines are the values recorded by a datalogger while symbols are instantaneous readings taken once per week. Solid line at the top indicates the period of differential irrigation. Asterisks at the top of each graph indicate irrigation events, where large-sized asterisks indicate a switch of the irrigated side for the PRD treatment
In 2002, the decline in ψs at 0.45 and 0.75 m in DI and PRD was more pronounced than in 2001 (Fig. 5). The maximum ψs at 0.15 m for PRD was always higher than those of CI and DI, with the exception of DOY 209−212 when ψs of PRD was up to 8 KPa lower than that of CI. However, the maximum values of ψs for PRD at 0.45 and 0.75 m were lower than that of CI by as much as 35 and 45 KPa, respectively. In addition, minimum values of ψs for PRD were up to 54 and 38 KPa lower than maximum values (data not shown). With DI, ψs at 0.45 m dropped sharply within three weeks of the start of the irrigation treatments and reached −200 KPa, which is the lower limit for the type of sensor used in our study, by August 11 (DOY 212). Soil-water potential at 0.75 m started to decline about two weeks later and leveled off around −160 KPa. The pattern for DI was very similar in 2003, except for a brief recovery at 0.45 m following the second additional irrigation applied just before the fourth switch for PRD. This additional irrigation also slowed the decline of ψs at 0.75 m, which reached −200 KPa in late September. In contrast, maximum ψs values with PRD were near those of CI at all depths and never dropped below −24, −36, and −36 KPa at 0.15, 0.45, and 0.75 m, respectively.
Fruit growth
In 2001, fruit diameter early in the growing season was similar for all treatments but was smaller for DI at harvest (Fig. 6). However, the reduced fruit growth and final size in DI was due to a much higher crop load on two out of the four DI trees, and no treatment differences were found when taking into account crop loads. Both absolute and relative fruit growth rates (data not shown) in the DI, excluding the two over-cropped trees, and PRD regimes were lower than the CI during the first three weeks in September. Cumulative growth over this period was about 15% less with DI and PRD compared to CI. It is worth noting that the mean fruit growth rate in the final week, i.e., the week after re-wetting the soil profile, was about 25–50% higher in PRD and DI than in CI.

Effect of irrigation regime on the mean fruit diameter of ‘Fuji’ apples grown near Prosser, WA. Vertical bars indicate LSD (P≤0.05)
In 2002, fruit size was again similar between treatments early in the season (Fig. 6) but fruit growth rate was lower in both DI and PRD compared to CI from late July until the final week before harvest. As in 2001, the mean fruit growth rate during the week following re-wetting of the soil profile was 10–20% higher in DI and PRD than in CI (data not shown). Final fruit diameter was 80.1 mm for CI, 77.9 mm for PRD, and 77.6 mm for DI. There were no treatment effects on fruit diameter and fruit growth rate during 2003 (Fig. 6).
Fruit size and yield
Harvest data (Table 1) show a trend toward lower fruit size with DI and, to a lesser extent, with PRD. The DI had the lowest mean fruit size and the lowest yield in all years. Cumulative yield was about 10% lower than CI and PRD. A tendency to reduce fruit size with DI is also evident from the crop profile (Fig. 7). Note that 2001 data from the two over-cropped DI trees have been omitted in Fig. 7. If included, the size distribution for DI is 24.3, 22.5, 34.7, 19.2, and 9.3% for fruit sizes of <70, 70–76.2, 76.2–82.5, 82.5–89, and >89 mm, respectively. In 2002, more than 40% of the fruit from PRD and DI were smaller than 76.2 mm compared to 24% for CI (Fig. 6). However, except for a significantly lower yield with DI compared to CI in 2002, treatment differences in yield, mean fruit size, and crop load were not significant (Table 1).

Effect of irrigation regime on fruit size distribution of ‘Fuji’ apples grown near Prosser, WA
Fruit quality
In all years and at all sampling times, DI fruit were firmer and had higher soluble solids than CI, although differences were often not significant (Table 2). In 2001, there were no significant differences in fruit quality parameters among the three irrigation regimes except for soluble solids and titratable acidity. It should be noted that in 2001, apples were harvested in a single picking about two weeks prior to their optimum maturity. Fruit from DI had higher soluble solids than fruit from CI both at 1 and 14 days after harvest, while PRD fruit had soluble solids that were not significantly different from either DI or CI (Table 2). Titratable acidity was significantly lower in DI and PRD than in CI at both sampling times, and was even lower in DI than in PRD at 14 days after harvest. Ethylene and CO2 evolution of apples stored for 14 days in respiration chambers at room temperature (20°C) were more than two times higher for DI and PRD fruit than CI fruit.
In contrast to 2001, titratable acids were significantly higher in DI and PRD fruit than CI fruit 1 day after harvest in 2002, while there were no treatment differences 14 days after harvest (Table 2). Similar to 2001, DI and PRD fruit had a significantly higher concentration of soluble solids than CI fruit at both sampling times. Fruit firmness was lower in CI than in both DI and PRD only at one day after harvest. No significant treatment effects were found in 2003.
Discussion
Irrigation, soil water, and orchard water use
Higher irrigation inputs in DI and PRD for 2002 are due, in part, to a longer irrigation season compared to 2001, but the length of the irrigation season was similar in 2003. The main reason for the comparatively high volumes applied in 2002, however, was excessive irrigation late in the season to re-wet the soil profile prior to winter dormancy. Re-wetting the soil profile prior to winter is a common practice in regions throughout the Western United States that are semi-arid with cold winters. Empirical evidence suggests that winter injury to trees and vines is higher under conditions of dry soil. As irrigation water is generally not available during the winter months and winter precipitation is often insufficient to refill a dry soil profile, growers refill the soil profile with the final irrigation(s) of the season. In our study, measurements of soil-water content (Fig. 2) just prior to re-wetting indicate a soil-water depletion of approximately 120 mm in the DI and PRD, yet more than 300 mm were applied. Such large application negated most of the water savings achieved with DI and PRD during the treatment period. In contrast, the irrigation volume applied in CI to re-wet the soil profile was only 160 mm, which was still excessive as the soil water content in the CI was close to field capacity prior to re-wetting (Fig. 2).
Measurements of soil-water content (Figs. 1, 2) and ψs (Figs. 3, 4) show that soil moisture in CI was kept close to field capacity. We propose that orchard water use can be reasonably accurately calculated from irrigation inputs, rainfall, and changes in soil-water content in the wetted zone, at least for the final two years. We argue that ignoring the contribution from the non-irrigated inter-row area introduces only a minor error under the semi-arid conditions of our site. Soil-water content in the inter-row area was measured from 2001 to 2003 in a companion study with ‘Fuji’/M9 with similar soil (both type and profile depth) and climatic conditions. At that site, soil-water content in spring was always at or above field capacity due to the use of sprinkler irrigation for frost control. Sprinklers were used only for frost control and re-wetting in the fall, while irrigation from early May to October was by sub-surface drip. Measurements over three years show that soil-water content in the inter-row area declined from field capacity values in spring to about 50% of field capacity within a single season (Caspari et al., unpublished data). At our site, precipitation from November 1 to April 30 was insufficient to completely replenish such a deficit, while ET0 over this period was 1.7–2.7 times that of precipitation. Also, a large proportion of any rainfall would have been used by the established grass cover crop rather than being available to the trees. Further, apple trees have been shown to quickly adjust their root water uptake in response to changing soil-water conditions by increasing uptake from a wet part of the rootzone while reducing uptake from a drying part (Green and Clothier 1999; Caspari and Lang, unpublished data). So, omitting the contribution from the inter-row area is likely to cause only a small underestimation of actual orchard water use.
Irrigation volumes required to keep soil moisture content of CI near field capacity from early June to the end of the third week in September were 498 mm in 2001, 488 mm in 2002, and 476 mm in 2003. Thus, irrigation requirements differed by less than 5% between years. Including cumulative rainfall in this period of 29, 26, and 12 mm indicates a mean daily orchard water use of 4.4−4.6 mm, or 70% of the mean daily ET0 of 6.24 mm. Irrigation scheduling based on soil-water measurements required 26% less water than what was predicted by the ETc model for an apple orchard without a cover crop by PAWS, which uses locally derived crop coefficients (James et al. 1989) similar to those proposed by FAO (Doorenbos and Pruitt 1977; Allen et al. 1998). Taking into account the soil moisture depletion with DI and PRD, a further 10–20% of irrigation water was conserved with DI and PRD strategies with only minor impacts on fruit size and yield.
The surface area wetted with DI was double that of PRD. Therefore, the higher apparent water use with DI is likely due in part to higher soil evaporation from the larger area wetted by irrigation (Esw). However, the differences appear to be too large to be explained by differences in Esw alone. For example, a difference of 80 mm in 2001 would imply that Esw was 160 mm for DI, which would represent 59% of the irrigation volume applied to DI over the period. For 2002 and 2003, it would represent 29 and 46% of applied irrigation lost to Esw. Bonachela et al. (2001) developed a model to estimate soil evaporation from the area wetted by drip irrigation under the semi-arid conditions of Cordoba, Spain. For the period 4 June−27 September, these authors estimated a total Esw of 42 mm in a mature olive orchard with 45% ground cover where the fraction of area wetted by daily drip irrigation was 10%. Although the larger area wetted by the micro-sprinklers in our study increases Esw, it also has a negative effect on Esw by decreasing the microadvective effect on E (Bonachela et al. 2001). Further, ground cover was 65–70% in our study and the wetted surface area was under the tree canopy. And with the exception of two additional irrigations in 2003, irrigation was applied only once a week. During the summer months, the wetted soil surface area dried off within 24–48 h following an irrigation so that soil evaporation proceeded at stage 2, the‘falling rate stage’ (Allen et al. 1998), for 5–6 days of a week. For 2003, the difference of ∼0.045 m3 m−3 in mean soil moisture between DI and PRD was already established by 1 August, i.e., after only 10 out of a total of 20 irrigation events during the treatment period. It would require an Esw of 90 mm, or 63% of irrigation volumes applied in DI, if this difference was solely due to soil evaporation.
The lower soil-water depletion with PRD may in part also be due to a more effective and/or prolonged delivery of a chemical signal, presumably ABA, from roots to shoots under PRD than DI. ABA is produced in roots when exposed to soil drying and is transported in the xylem into leaves. In the leaves, ABA induces partial stomatal closure, which increases water use efficiency (Davies and Zhang 1991; Tardieu et al. 1992a, b). Such adjustment in response to partial drying of the roots has also been observed with apple. Using apple seedlings with a split root system where one half of the root system was allowed to dry, Gowing et al. (1990) demonstrated a signal originating from the drying roots, although they did not identify the chemical involved. Dry et al. (1996), using PRD on split-rooted grapevines, showed that the ABA signal was transient and that the ABA signal would disappear if the soil moisture in the dry half was maintained continuously at a deficit. These authors concluded that to maintain transport of an ABA signal to the shoots, a fraction of the rootzone has to be in drying, not dry, soil. Frequent switches of irrigation from one side of the PRD trees to the other may have maintained a larger portion of the root system exposed to drying soil compared to DI where more of the root system may have been in dry soil. This larger portion of roots exposed to drying soil with PRD may have resulted in more ABA arriving in the leaves and affecting stomatal opening, and hence water loss. However, we do not have any ABA data from the current study to test this hypothesis. Irrespective of the mechanism(s), measurements of soil-water content (Figs. 2, 3) and soil-water potential (Figs. 4, 5) show different patterns in soil-water depletion between DI and PRD. The larger depletion with DI required an additional 80, 42, and 75 mm in 2001, 2002, and 2003, respectively, to return the soil to field capacity.
Fruit growth and yield
Measurements of soil-water content (Figs. 1, 2) and ψs (Figs. 3, 4) show that soil moisture in CI was always high enough so as to not limit fruit growth. Ebel et al. (2001) reported that fruit growth rate of ‘Delicious’ apple growing at the same research orchard as in our study was still 97% of that of a well-watered control at 35% of total available water (∼55% of field capacity or 0.15 m3 m−3). Mean soil-water content in CI never fell below 75% of field capacity in our study. In 2002, mean fruit growth rates of DI and PRD started to decline from that of CI in late July, and were 10–25% less than that of CI until October. Mean soil-water content was 0.15 m3 m−3 on August 2 and remained between 0.13 and 0.15 m3 m−3 until re-wetting in October. Likewise, mean fruit growth rate with DI was up to 8% lower than CI from late August to early October 2003, when soil moisture content in DI was below 0.15 m3 m−3 . Our data with ‘Fuji’ are in close agreement with Ebel et al. (2001) and confirm that apple fruit growth is reduced when soil-water content falls below 35% of total available water. A very similar threshold has been found during 2001–2003 in our companion study with ‘Fuji’/M9 (Caspari et al., unpublished data).
A reduction in final fruit size is one of the most consistently reported effects of deficit irrigation of apples (Lötter et al. 1985; Ebel et al. 1993, 1995; Mpelasoka et al. 2000, 2001). Since 1997, we have conducted a number of trials with ‘Fuji’ and ‘Braeburn’ to determine if PRD is different from other forms of deficit irrigation. In the New Zealand climate, we have found only small differences between DI and PRD to a well-watered control, or between PRD and DI (Caspari and Neal, unpublished). However, under very dry conditions such as during the 2000/2001 season, mean fruit size and marketable yield of ‘Braeburn’ was reduced with DI but not PRD (Caspari et al. 2003). In the current study, fruit growth (Fig. 6), harvest data (Table 1), and crop profile show a tendency for smaller fruit and lower yield with DI, although higher crop load in DI in 2001 may have contributed to the reduction in fruit size. The effects on fruit growth (Fig. 6), final fruit size (Table 1), and crop profile (Fig. 7) were less pronounced with PRD than with DI, but there was no significant difference between PRD and DI in any year. Although fruit size and yield tended to be less affected by PRD than by DI in this and previous studies (Caspari et al. 2003; Caspari and Neal, unpublished data), further research is required to determine if PRD is indeed different from other types of DI. Aside from the differences in soil-water depletion discussed above, it is worth noting that we have found very distinct differences between PRD and DI in leaf color and leaf drop late in the 2002 and 2003 seasons in our companion study with ‘Fuji’/M9 and one ‘Bing’ sweet cherry (Prunus avium L.) trial in Washington State (Caspari, unpublished data; Whiting, unpublished data).
Fruit quality
Our results agree with previous studies that have shown an increase in the concentration of soluble solids in apple in response to DI (Proebsting et al. 1984; Irving and Drost 1987; Ebel et al. 1993; Mpelasoka et al. 2000). In contrast to soluble solids, the effect of reduced irrigation on titratable acidity is less clear. While some studies have shown a reduction of titratable acids in response to water deficits (Guelfat Reich et al. 1974; Drake et al. 1981; Proebsting et al. 1984), others have found no effect (Irving and Drost, 1987; Kilili et al. 1996; Mpelasoka et al. 2000). Our own data are inconclusive as titratable acidity was affected in opposite ways in the first two seasons, and we did not determine titratable acidity in 2003. An increase in fruit firmness observed with DI may be an indirect effect due to a reduction in fruit size as small fruit tend to be firmer (Ebel et al. 1993; Mpelasoka et al. 2000; Volz et al. 2003). However, Mpelasoka et al. (2000) compared fruits of similar size, and found fruits from early or late DI treatments to be firmer than control fruit at harvest, as well as after 12 weeks of cold storage. In our study, fruits were of similar size (Table 2), and the results are in agreement with Mpelasoka et al. (2000). The DI fruit was firmer than the CI fruit at all sampling times, with significant differences 1 day after harvest in 2002. In 2001 and 2003, PRD fruit had similar or slightly lower firmness than CI but were significantly firmer than CI 1 day after harvest in 2002. Preliminary data from the two sampling dates in 2003 show higher fruit dry matter concentration with DI that may have contributed to higher firmness. Higher dry matter concentration in fruit from deficit treatments has previously been reported (Proebsting et al. 1984; Mpelasoka et al. 2000).
Starch pattern index and ethylene concentration are used as indicators of apple maturity (Kingston 1992), and the former is widely used in the Washington State apple industry to assess fruit maturity. There was no treatment effect on the starch pattern index in any year, suggesting that fruit ripening was not affected. In contrast, higher ethylene and CO2 evolution in DI and PRD fruit held at room temperature in 2001 suggest more advanced maturity with DI and PRD. The limited data on fruit maturity from our study are inconclusive.
Summary and conclusions
‘Fuji’ apple trees grown in deep soils with high water holding capacity showed no statistically significant differences in yield, fruit size, and crop yield among control irrigation (CI), deficit irrigation (DI) or partial rootzone drying (PRD) treatments in 2001 and 2003. However, yield was significantly reduced by DI compared to that by CI in 2002, while PRD did not differ significantly from CI or DI. Further, the concentration of soluble solids tended to be higher in PRD and DI than in CI. These differences from CI were significant in the first two years for DI and in the second year only for PRD. Treatment effects on titratable acidity were inconclusive, being significantly lower in DI and PRD than in CI in the first year and significantly higher in the second year. Fruit from DI was always firmer than that from CI, with fruit from PRD being intermediate.
Irrigation volumes applied in CI were only 70–75% of the modeled ETc from PAWS weather data in all the years excluding the excessive late-season irrigation in 2002. The soil-water content in CI was kept near field capacity throughout all seasons. Thus, basing irrigation decisions on soil-water monitoring rather than modeled ETc resulted in significant water conservation. A further 20–25% of irrigation water was conserved with DI and PRD with no negative impacts on yield and fruit size with PRD. However, yield was reduced with DI compared to CI in the second year.
Our results indicate the potential of DI and PRD strategies for large water savings with no or small impacts on yield and fruit size. They also confirm that DI strategies can be used to alter fruit quality. It required only a small increase in irrigation volumes in DI and PRD to avoid reductions in yield and fruit size in the final year while maintaining the effects on fruit quality. Similar to previous studies with apple (Caspari et al. 2003; Caspari and Neal, unpublished data) and the results presented here, de Souza et al. (2003) and dos Santos et al. (2003) reported only subtle differences comparing PRD and DI with grape varieties ‘Moscatel’ and ‘Castelão’ in southern Portugal. Further experiments comparing PRD and DI are required to determine the importance of placement versus volume of irrigation water.
Twenty years ago, after a 3-year study on DI with ‘Delicious’ and ‘Golden Delicious’ apples, Proebsting et al. (1984) concluded that ‘minimizing plant water deficits throughout the season, the predominant irrigation philosophy, may prove to be only one of several possible strategies for water management in apple orchards’. We agree.
References
Alegre S, Girona J, Marsal J, Arbones A, Motilva MJ, Romero MP (1997) Regulated deficit irrigation in olive trees. Acta Hort 474:373–376
Allen RG, Pereira LS, Raes D, Smith M (1998) Crop evapotranspiration—Guidelines for computing crop water requirements. FAO Irrigation and Drainage Paper No. 56. Food and Agricultural Organisation of the United Nations, Rome
Boland AM, Mitchell PD, Jerie PH, Goodwin I (1993) The effect of regulated deficit irrigation on tree water use and growth of peach. J Hort Sci 68:261–274
Bonachela S, Orgaz F, Villalobos F, Fereres E (2001) Soil evaporation from drip-irrigated olive orchards. Irrig Sci 20:65–71
Caspari HW, Behboudian MH, Chalmers DJ (1994) Water use, growth, and fruit yield of ‘Hosui’ Asian pears under deficit irrigation. J Am Soc Hort Sci 119:383–388
Caspari HW, Neal S, Alspach P (2004) Partial rootzone drying—a new deficit irrigation strategy for apple? Acta Hort 646:93–100
Chalmers DJ, Mitchell PD, Heek L van (1981) Control of peach tree growth and productivity by regulated water supply, tree density, and summer pruning. J Am Soc Hort Sci 106:307–312
Davies WJ, Zhang J (1991) Root signals and the regulation of growth and development of plants in drying soil. Ann Rev Plant Physiol Plant Mol Bio 42:55–76
Davies WJ, Wilkinson S, Loveys B (2002) Stomatal control by chemical signalling and the exploitation of this mechanism to increase water use efficiency in agriculture. New Phytol 153:449–460
de Souza CR, Maroco JP, dos Santos TP, Rodrigues ML, Lopes CM, Pereira JS, Chaves MM (2003) Partial rootzone drying: regulation of stomatal aperture and carbon assimilation in field-grown grapevines (Vitis vinifera cv. Moscatel). Funct Plant Biol 30:653–662
Doorenbos J, Pruitt WO (1977) Guidelines for predicting crop water requirements. FAO Irrigation and Drainage Paper No. 24. Food and Agricultural Organisation of the United Nations, Rome
dos Santos TP, Lopes CM, Rodrigues ML, Souza CR, Maroco JP, Pereira JS, Silva JR, Chaves MM (2003) Partial rootzone drying: effects on growth and fruit quality of field-grown grapevines (Vitis vinifera). Funct Plant Biol 30:663–671
Drake SR, Proebsting EL, Mahan MO, Thompson JB (1981) Influence of trickle and sprinkle irrigation on ‘Golden Delicious’ apple quality. J Am Soc Hort Sci 106:255–258
Dry PR, Loveys BR (1998) Factors influencing grapevine vigour and the potential for control with partial rootzone drying. Aust J Grape Wine Res 4:140–148
Dry PR, Loveys BR, Botting DG, Düring H (1996) Effects of partial root-zone drying on grapevine vigour, yield, composition of fruit and use of water. In: Stockley CS, Sas AN, Johnstone RS, Lee TH (eds) Proceedings of the 9th Australia Wine Industrial Technical Conference. Winetitles, Adelaide, Australia, pp128–131
Dry PR, Loveys BR, Düring H (2000a) Partial drying of the rootzone of grape. I. Transient changes in shoot growth and gas exchange. Vitis 39:3–7
Dry, PR, Loveys BR, Düring H (2000b). Partial drying of the rootzone of grape. II. Changes in the pattern of root development. Vitis 39:9–12
Dry PR, Loveys BR, McCarthy MG (2001) Strategic irrigation management in Australian vineyards. J Intl Sci Vin 35:129–139
Ebel RC, Proebsting EL, Patterson ME (1993) Regulated deficit irrigation may alter apple maturity, quality, and storage life. Hort Sci 28:141–143
Ebel RC, Proebsting EL, Evans RG (1995) Deficit irrigation to control vegetative growth in apple and monitoring fruit growth to schedule irrigation. Hort Sci 30:1229–1232
Ebel RC, Proebsting EL, Evans RG (2001) Apple tree and fruit responses to early termination of irrigation in a semi-arid environment. Hort Sci 36:1197–1201
Gardner WH (1986) Water content. In: Klute A (ed) Methods of soil analysis Part 1 Physical and mineralogical methods (Agronomy monograph 9). ASA and SSSA, Madison, WI, pp 635–662
Goldhamer DA (1997) Regulated deficit irrigation for California canning olives. Acta Hort 474:369–372
Gowing DJG, Davies WJ, Jones HG (1990) A positive root-sourced signal as an indicator of soil drying in apple, Malus × domestica Borkh. J Exp Bot 41:1535–1540
Green SR, Clothier BE (1999) The root zone dynamics of water uptake by a mature apple tree. Plant Soil 206:61–77
Guelfat’Reich S, Assaf R, Bravdo BA, Levin I (1974) The keeping quality of apples in storage as affected by different irrigation regimes. J Hort Sci 49:217–225
Irving DE, Drost JH (1987) Effects of water deficit on vegetative growth, fruit growth and fruit quality in Cox’s Orange Pippin apple. J Hort Sci 62:427–432
James JG, Erpenbeck JM, Bassett DL, Middleton JE (1989) Irrigation requirements for Washington—estimates and methodology EB 1513 Cooperative Extension, College of Agric Home Econom. Washington State University, Pullman, WA
Kilili AW, Behboudian MH, Mills TM (1996) Composition and quality of ‘Braeburn’ apples under reduced irrigation. Sci Hort 67:1–11
Kingston CM (1992) Maturity indices for apple and pear. Hort Rev 13:407–432
Landsberg JJ, Jones HG (1981) Apple orchards. In: Kozlowski T (ed) Water deficit and plant growth, vol 6. Academic Press, New York, pp 419–469
Li S-H, Huguet J-G, Schoch PG, Orlando P (1989) Response of peach tree growth and cropping to soil water deficit at various phenological stages of fruit development. J Hort Sci 64:541–552
Lötter JDeV, Beukes DJ, Weber HW (1985) Growth and quality of apples as affected by different irrigation treatments. J Hort Sci 60:181–192
Loveys BR, Grant WJR, Dry PR, McCarthy MG (1997) Progress in the development of partial root-zone drying. Aust Grapegrow Winemak 403:18–20
Loveys B, Stoll M, Dry P, McCarthy M (1998) Partial rootzone drying stimulates stress responses in grapevine to improve water use efficiency while maintaining crop yield and quality. Aust Grapegrow Winemak Annu Tech Issu 108–113
Marsal J, Mata M, Arbonés A, Rufat J, Girona J (2002) Regulated deficit irrigation and rectification of irrigation scheduling in young pear trees: an evaluation based on vegetative and productive response. Eur J Agric 17:111–122
Mitchell PD, Chalmers DJ (1982) The effect of reduced water supply on peach tree growth and yields. J Am Soc Hort Sci 107:853–856
Mitchell PD, Ende B van den, Jerie PH, Chalmers DJ (1989) Responses of ’Bartlett’ pear to withholding irrigation, regulated deficit irrigation, and tree spacing. J Am Soc Hort Sci 114:5–19
Mpelasoka BS, Behboudian MH, Dixon J, Neal SM, Caspari HW (2000) Improvement of fruit quality and storage potential of ‘Braeburn’ apple through deficit irrigation. J Hort Sci Biotechnol 75:615–621
Mpelasoka BS, Behboudian MH, Green SR (2001) Water use, yield and fruit quality of lysimeter-grown apple trees: responses to deficit irrigation and to crop load. Irr Sci 20:107–113
PAWS, Public Agricultural Weather System-WSU-PAWS Washington State Univer. Prosser, WA. http://frost.Prosser.wsu.edu/
Proebsting EL, Drake SR, Evans RG (1984) Irrigation management, fruit quality, and storage life of apple. J Am Soc Hort Sci 109:229–232
Tardieu F, Zhang J, Davies WJ (1992a) What information is conveyed by an ABA signal from maize roots in drying field soil? Plant Cell Environ 15:185–191
Tardieu F, Zhang J, Katerji N, Bethenod O, Palmer S, Davies WJ (1992b) Xylem ABA controls the stomatal conductance of field-grown maize subjected to soil compaction or soil drying. Plant Cell Environ 15:193–197
Volz RK, Harker FR, Lang S (2003) Firmness decline in ‘Gala’ apple during fruit development. J Am Soc Hort Sci 128:797–802
Acknowledgements
Recognition is given to the Washington Tree Fruit Research Commission and Washington State University Irrigated Agriculture Research and Extension Center (IAREC) for supporting this research effort. We gratefully acknowledge the technical support of Gary Matthews, and David Strausz III from WSU-Prosser, and Gregory Peck, Margaret Collier, and Scott Mattinson from WSU-Pullman.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by E. Fereres
Rights and permissions
About this article
Cite this article
Leib, B.G., Caspari, H.W., Redulla, C.A. et al. Partial rootzone drying and deficit irrigation of ‘Fuji’ apples in a semi-arid climate. Irrig Sci 24, 85–99 (2006). https://doi.org/10.1007/s00271-005-0013-9
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00271-005-0013-9