
Abstract
The development of effective strategies to restore the biological functioning of aquatic ecosystems with altered flow regimes requires a detailed understanding of flow-ecology requirements, which is unfortunately lacking in many cases. By understanding the flow conditions required to initiate critical life history events such as migration and spawning, it is possible to mitigate the threats posed by regulated river flow by providing targeted environmental flow releases from impoundments. In this study, we examined the influence of hydrological variables (e.g., flow magnitude), temporal variables (e.g., day of year) and spatial variables (e.g., longitudinal position of fish) on two key life history events (migration to spawning grounds and spawning activity) for a threatened diadromous fish (Australian grayling Prototroctes maraena) using data collected from 2008 to 2015 in the Bunyip–Tarago river system in Victoria. Our analyses revealed that flow changes act as a cue to downstream migration, but movement responses differed spatially: fish in the upper catchment showed a more specific requirement for rising discharge to initiate migration than fish in the lower catchment. Egg concentrations peaked in May when weekly flows increased relative to the median flow during a given spawning period. This information has recently been incorporated into the development of targeted environmental flows to facilitate migration and spawning by Australian grayling in the Bunyip–Tarago river system and other coastal systems in Victoria.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Many of the world’s freshwater ecosystems have been severely affected by human activity, especially where water resources are in great demand. Natural flow regimes have often been greatly altered by water storage, extraction and even seasonal inversion, causing major changes to the ecology and functioning of riverine ecosystems (Bunn and Arthington 2002). The strong life history dependence of riverine fish on the natural flow regime has resulted in widespread declines in abundance and species diversity in flow-impacted systems (Murchie et al. 2008; Poff and Zimmerman 2010). For example, increases in river flow that are an important cue for movement to spawning grounds in many freshwater fishes have often been disrupted or completely stopped (e.g., O’Connor and Mahoney 2004). In response, strategies are being developed and implemented in regulated riverine environments such as the provision of flows to conserve or restore important ecological processes and functions (Arthington 2012). However, the development of effective strategies requires a detailed understanding of flow-ecology relationships, which are unfortunately lacking for many species in impacted systems.
Although a growing body of scientific literature exists in relation to fish–flow relationships (e.g., Taylor and Cooke 2012; Stewart-Koster et al. 2013; Koster et al. 2016), studies have often focused on single events with few examples of long-term monitoring (>3 years), which limits the temporal applicability and robustness of findings (e.g., Growns and James 2005; King et al. 2015a), or involve biological metrics (e.g., adult abundance) that make it difficult to establish, which flow components are directly responsible for observed changes (Poff and Zimmerman 2010). Many studies also lack clear hypotheses concerning specific flow impacts on key life history stages that are then explicitly tested with targeted data collection (King et al. 2015b). Such an ad-hoc approach to exploring fish–flow relationships severely limits the development of targeted and effective flow management strategies for fish conservation (Koehn et al. 2014) because there is no underlying conceptual model that allows meaningful predictions of ecological outcomes from flow scenarios to be made (Morrongiello et al. 2011).
In this study, we examined the influence of flow on key aspects of the life history (migration and spawning) of a threatened fish Australian grayling (Prototroctes maraena), in the Bunyip–Tarago river system in Victoria, Australia, using acoustic telemetry and drift sampling of eggs. The Australian grayling is a diadromous species that undertakes downstream migrations to lower fresh water reaches of rivers to spawn in autumn in response to increased flows, and the eggs and larvae drift downstream to the sea, from where juveniles migrate back into fresh water in spring–summer (Berra 1982; Crook et al. 2006; Koster et al. 2013). Promoting migration to spawning grounds and increasing the occurrence of successful spawning events by providing flow cues is a key management action to support the reproductive requirements of this species and assist the recovery of its populations (Backhouse et al. 2008; SKM 2012). Indeed, environmental flows aimed specifically at initiating migration and spawning in Australian grayling have been introduced into the management of the Bunyip–Tarago river system and other coastal systems in Victoria in recent years by the Victorian Environmental Water Holder with planning and delivery in partnership with Catchment Management Authorities and Melbourne Water.
Early environmental flow recommendations for the Australian grayling focused on delivering short-lived (e.g., 1-day) flows to trigger spawning (e.g., SKM 2005; Earth Tech 2006). This was based on research suggesting that adults spawn in upriver reaches and do not migrate to spawn (Berra 1982). However, following the identification of a long-distance downstream migration to spawning grounds just upstream of the estuary in response to increased flow (Koster et al. 2013), it was evident that environmental flow recommendations for the Australian grayling needed to include flows that allowed adult migration to spawning areas. This has recently been achieved through reviews of the environmental flow recommendations for the Australian grayling in the Bunyip–Tarago river system and other coastal systems in Victoria (SKM 2013; Alluvium 2015; Jacobs 2015).
While recent research has resulted in significant new information on the spawning and migration of Australian grayling and their links to flows, evaluations of the flow requirements of this species have been based on short-term studies (e.g., 2 years) and constrained by factors such as small sample sizes (Koster et al. 2013; Amtstaetter et al. 2015; Koster et al. 2017). In this study, we integrated all of the available movement and spawning data collected between 2008 and 2015 on Australian grayling in the Bunyip–Tarago river system, and examined the influence of a priori determined hydrological, temporal and spatial variables on Australian grayling migration to spawning grounds and spawning activity. The study uses data previously analysed by Koster et al. (2013, 2017), but builds on this previous work by incorporating more data and using more sophisticated statistical methods. For migration, we predicted that short-term flow changes act as a cue for movement, although we also tested for absolute flow magnitude effects. For spawning, we predicted that fish respond to changes in flow relative to prevailing conditions, rather than the absolute level of river discharge. This information is needed to improve the understanding about how components of the flow regime such as timing, duration, and magnitude affect migration and spawning in order to develop and refine environmental flow delivery strategies.
Methods
Study Area
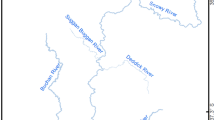
The study was conducted in the Bunyip–Tarago river system in Victoria, south-eastern Australia (Fig. 1). The Tarago river is the main tributary of the Bunyip river and is regulated by the Tarago reservoir. The upper reaches of the Bunyip–Tarago catchment are forested, while the mid to lower reaches consist predominantly of cleared agricultural land and small urban areas. Average annual discharge in the Bunyip River is about 153,000 ML.

The location of the study site in the Bunyip–Tarago river system in Victoria, Australia. Black circle represents the drift sampling site and white triangles the acoustic listening stations
Fish Movement
A total of 81 Australian grayling were collected by electrofishing in 2009, 2010 and 2015 from the Bunyip and Tarago rivers, and acoustic transmitters were surgically implanted in each fish (see Koster et al. 2013 for details). The lengths of fish ranged from 150 to 240 mm, with a mean of 203 mm (±SE = 1.9 mm). Different fish were tagged in each year. Vemco V7 transmitters (22 × 7 mm, frequency 69 kHz, mass 1.8 g in air, estimated battery life 150–170 days; Vemco, Nova Scotia, Canada) were used in 2009 (n = 21) and 2010 (n = 34). In 2015, 26 fish were tagged with newly available Vemco V4 transmitters (11 × 5.7 mm, frequency 180-kHz, mass 0.4 g in air, estimated battery life 50–60 days). An array of Vemco VR2W acoustic listening stations was deployed in the Bunyip River between Bunyip and Koo Wee Rup, and in the Tarago River between Drouin West and the Bunyip River junction (Fig. 1). Each listening station was deployed using a length of plastic-coated steel cable attached to a star picket as an anchor point. Data were downloaded from the listening stations at approximately monthly intervals throughout the study.
Fish Spawning
Fish eggs were collected at one site on the lower reaches of the Bunyip River near Koo Wee Rup using two drift nets (Fig. 1). Previous sampling at other sites had shown that the vast majority of eggs occurred in the lower reaches (Koster et al. 2013; Koster et al. 2017). Sampling was conducted once or twice per week from March to June in each year from 2008 to 2015, except 2012. Drift nets were of 500-µm mesh, 150 cm long with a 50 cm mouth diameter, and had flow meters (General Oceanics, Florida, USA) fitted to the mouth of the net to measure the volume of water filtered. Nets were set in late afternoon (1500–1700 h) and retrieved the following morning (08:00–10:00 h). Drift-net samples were immersed into an anaesthetic solution of 4 mL Alfaxan (Jurox, Rutherford, Australia) per litre water for 10 min to euthanase any fish, and then preserved in 70% ethanol. These samples were sorted in the laboratory under a dissecting microscope.
Data Analysis
We modelled the probability of an individual Australian grayling undertaking a downstream movement on a given day (binary response variable: 0 or 1) during the spawning season or spawning activity (counts of eggs) as a function of hydrological, temporal and spatial predictor variables (Table 1) using generalised linear mixed effects models (GLMMs). The analysis for spawning activity included counts only of eggs, not larvae, as larvae comprised only about 10% of the total catch. Hydrological variables were defined a priori based on our understanding of how Australian grayling are likely to respond to changes in their environment (Koster et al. 2013; Koster et al. 2017). GLMMs were fitted using a binomial distribution in the lme4 package in R 3.2.2 (R Development Core Team 2015).
Movement model
As there were multiple daily observations of an individual’s movement status, a GLMM was fitted with a random intercept for individual fish (Fish ID) (Bestley et al. 2010; Koster et al. 2013). The random intercept induces a correlation among observations within a fish and accounts for the repeated measures nature of the data.
We compared a series of increasingly complex models fitted to the movement data that explored potential spatially and temporally dependent flow responses (flow*DOY (day of year); flow*distance), and year-to-year variation in seasonal migration patterns (Year*DOY). We also allowed for curvilinear flow responses by fitting a quadratic term. All models included Lag-1 to account for a potential Markov process in which future movement states depend on preceding state and temporal autocorrelation in the data (Bestley et al. 2010). Continuous predictors were first scaled [(x-mean)/SD] to ensure model convergence. In the case of FlowMagCurrent and FlowMagNat, data were then log10-transformed to ensure linearity between the predictor and the link function. Because of colinearity among hydrological variables, flow change, FlowMagCurrent and FlowMagNat were fitted in separate models. Competing models were compared using rescaled Akaike’s Information Criterion corrected for small sample sizes (ΔAICc) (Burnham and Anderson 2002). Parameter estimates and 95% credible intervals were derived from the posterior distribution of the fixed effects in the best model using 1000 model simulations generated by the arm R package.
Spawning model
The response variable in the spawning model (number of eggs) is a count with values that ranged from 0 to 3974. As these counts were dependent on the volume of water filtered by each net, we fitted an offset term for filtered flow volume that adjusted counts for sampling effort and still allowed us to predict values on the original, count scale. We used GLMMs that included Year as a random effect (seven levels) as we were not interested in these seven years per se, but rather treated them as a random draw of all possible years. We initially fitted a Poisson GLMM, but these models were significantly over-dispersed and, therefore, a negative binomial GLMM was used.
Because of strong colinearity (r = 0.94), we fitted DOY and water temperature models separately. We included quadratic terms for each variable to allow for curvilinear relationships with spawning magnitude and also tested for interactions between DOY or water temperature and each flow variable. Continuous predictors were scaled, competing models were compared using ΔAICc and parameter estimates and 95% credible intervals were generated using 1000 model simulations derived from the arm R package.
Results
Fish Movement
Tagged adult Australian grayling in the Bunyip–Tarago system undertook rapid downstream migrations over distances of up to 45 km to the lower reaches of the Bunyip River near Koo Wee Rup from late March to late April during increases in flow (Fig. 2). The percentage of tagged fish detected migrating to the lower reaches varied between years: 33% in 2009, 18% in 2010, and 85% in 2015. The much greater percentage of fish detected migrating in 2015 compared with 2009 and 2010 likely reflects a higher rate of transmitter retention in 2015, attributable to refinement of the tagging procedure. Movements to the lower reaches during the spawning season in response to increased flows were partially synchronised: some fish moved together for part of their journey, and although not all moved on the same flow event, all fish migrated at the same time of year.

Examples of movement patterns of individual Australian grayling tagged in 2009, 2010 and 2015 in the Bunyip and Tarago rivers. Black circles show the date and location of tagging and grey circles show detections of tagged fish on the listening stations. Dark blue line denotes mean daily discharge in Bunyip River at Iona. Light blue line denotes mean daily discharge in Tarago River at Drouin West. Dashed red line denotes mean daily water temperature in the Bunyip River at Iona (2009, 2010) and Tarago River at Drouin West (2015). Figure 2a–d reproduced from Koster et al. (2013)
The best movement model included the terms Lag-1 and the interactions Year*DOY and flow change*distance (Table 2). Models including FlowMagCurrent and FlowMagNat received little support given the data (ΔAICc values > 21). Fish that moved at time t—1 (yesterday) were 2.78 times more likely to move at time t (today), indicating that migrations occurred over subsequent days. There was considerable among-year variation in the timing of spawning migrations (Fig. 3). In both 2009 and 2010, the probability of fish moving increased with time (peak spawning on average occurred around day140; see spawning results), whereas in 2015 fish had a greater probability of moving earlier in the season.

Predicted probability of fish migrating as a function of year and day-of-year (+/− 95% CI). Peak spawning occurred around day 140 (see spawning results). All other covariates held at their mean values
Fish were more likely to move downstream when higher in the river system, but this relationship was flow-change dependent (Fig. 4). Fish located higher in the catchment (more than 20 km above the estuary) moved only on the rising limb of flows (positive flow change) and had a higher probability of moving on larger flow changes. Once fish had moved farther down in the system, the probability of moving as a function of flow change was lower, and fish were just as likely to move on negative and positive flow changes (Fig. 4). The downstream migrations corresponded to the timing of spawning (Figs 2 and 5).

Predicted probability of fish moving downstream as a function of distance upstream from the spawning area and % flow change (positive values indicate rising limb, negative values falling limb of hydrograph). Grey scale colour bar denotes probability of movement. All other covariates held at their mean values

Adjusted total density of Australian grayling eggs per megalitre collected in drift nets in the Bunyip River. Dark blue line denotes mean daily discharge in Bunyip River at Iona. Dashed red line denotes mean daily water temperature in the Bunyip River at Iona (2008–2013) and Tarago River at Drouin West (2014–15). Clear triangles on x-axis indicate sampling event. Figure 5a–e reproduced from Koster et al. (2013) and Fig. 5f reproduced from Koster et al. (2017)
Fish Spawning
In total, 11,711 Australian grayling eggs were collected in the drift sampling (Table 3). Eggs were collected from April to June, with peak egg abundances collected between mid April and mid May, coinciding with autumn within-channel flow pulses (Fig. 5). Australian grayling larvae were much less abundant in the samples (n = 1310).
The best spawning model included an interaction between PerFlowMag and DOY, and their quadratic terms (Table 2). Collections of eggs peaked around day 140 (mid to late May) when flows in the week prior to sampling were about 300% higher than the median flow during the spawning period (March–May) (Fig. 6). Collections steadily declined at lower and higher flows, and also earlier and later in the year.

Predicted spawning activity (number of eggs per megalitre) as a function of % flow magnitude and day of year. Grey scale colour bar denotes magnitude of spawning
Eggs were collected over a broad range of water temperatures (as high as 18 °C), with peak abundances at 13–14 °C (Supplementary Fig. 1), but this model (which also included percentage flow magnitude) received little support from the data (ΔAICc > 25). This suggests that spawning is triggered by day length rather than just temperature.
Discussion
Our study analysed data collected over 8 years (2008–2015) for two key life history events associated with reproduction in a threatened diadromous fish. Previous studies of this species (i.e., Koster et al. 2013; Amtstaetter et al. 2015; Koster et al. 2017) were each based on 2–4 years of data only. Combining these data sets and incorporating new data provided temporal replication that allowed us to investigate potentially complex relationships between Australian grayling behaviour and flow variability, and generate a robust synthesis of how flow alteration and restoration affects this species. We confirm the importance of a rise in river flow for downstream migration and spawning of Australian grayling (Koster et al. 2013; Amtstaetter et al. 2015; Koster et al. 2017). More specifically, our analyses show that adult fish undertake partially synchronised downstream spawning migrations to freshwater reaches immediately upstream of the estuary predominantly during periods of increased streamflow. The statistical modelling also demonstrated that the rising limb of the hydrograph is a critical cue for initiating downstream spawning migration. Downstream migrations over long distances to spawn at times of increasing flow occur in many fish species for which transport of eggs or larvae to the sea is an obligatory part of the life cycle (Boubée et al. 2001; Durif and Elie 2008; Crook et al. 2010). For these fishes, movement on the rising hydrograph limb of a flow likely represents an evolutionary adaptation that allows individuals to take maximum advantage of a flow pulse for travelling to spawning grounds (i.e., reduced energy expenditure, improved connectivity) and to assist the subsequent transport of eggs or larvae to the sea (Northcote 1984).
The clear behavioural response of the Australian grayling to increases in river discharge highlights the threats posed by artificially altered flow regimes (i.e., altered frequency, magnitude and duration of flow events) because it shows that spawning and subsequent recruitment are unlikely to occur in the absence of specific flow cues during the autumn spawning period (see also O’Connor and Mahoney 2004). Damping of variations in streamflow caused by the capture of small to moderate rises in river discharge and the development of stable base flows is recognised as one of the most significant ecological effects of river regulation (e.g., McMahon and Finlayson 2003). Our study provides further evidence of how reduced flow variability can deleteriously impact the sustainability of riverine fish populations (Freeman et al. 2001). Our results also provide important insights into the influence of hydrology and its timing on spawning of Australian grayling. It has been suggested that spawning takes place from February to July (Bishop and Bell 1978; Berra 1982; Hall and Harrington 1989; Koehn and O’Connor 1990), but we have shown that the highest concentrations of eggs typically occurred in May when flows in the week prior to sampling are higher than the median flow during the spawning period for that year. Our analyses also identified an upper bound of flow beyond which spawning declines (i.e., when weekly flows greater than 300% of the seasonal median). Together, these results suggest fish respond to increases in flow relative to the prevailing seasonal conditions rather than the absolute level of river discharge, and that larger flows do not necessarily result in more spawning.
By understanding the flow conditions required to initiate migration and spawning, it is possible to mitigate the threats posed by regulated river flow by providing targeted environmental flow releases from impoundments. For example, a flow release designed to maximise the duration of the rising limb of the flow would likely increase movement activity and spawning by Australian grayling, and different flow magnitudes could be delivered in different rainfall years to match the prevailing seasonal conditions (as opposed to providing a single flow magnitude across all years). Consideration of flow duration and timing is now a key component of seasonal watering plans for the Bunyip–Tarago recently developed by the Victorian Environmental Water Holder and Melbourne Water (VEWH 2016), based on the supporting evidence from this study, and the responses of fish are currently being monitored. More broadly, our study demonstrates the importance of quantifying flow-ecology relationships for multiple life history events through targeted research to develop appropriate flow regimes for aquatic biota in regulated rivers (King et al. 2010; Koster et al. 2017). While our study found clear responses to flow, the specific mechanisms for how flow affects movement and spawning behaviours need to be explored further. For example, the roles of hydrodynamic characteristics that constitute flow such as velocity, depth and turbulence are unknown,despite the fact that fish are likely to respond to these rather than flow per se (Silva et al. 2011; Piper et al. 2015).
Another key finding of this study relates to the synchronised movement by Australian grayling to the lower reaches during the spawning season. Some fish moved together for part of their journey, and although not all fish moved on the same flow event, all migrated at the same time of year. Furthermore, once an individual commenced migrating, it was more likely to keep moving. Similar synchronised patterns have been observed for other flow-dependent spawning species, such as Colorado pikeminnow (Ptychocheilus lucius) in White River, Colorado (Irving and Modde 2000), and surubim (Pseudoplatystoma corruscans) in the São Francisco River, Brazil (Godinho et al. 2007). Synchronised behaviour in species with reproductive strategies strongly linked to flow (and, therefore, somewhat opportunistic) may allow individuals to maximise their ability to successfully exploit favourable environmental conditions when they occur, thereby enhancing reproductive success (Lytle and Poff 2004). For species such as the Australian grayling that undertake synchronised movements, the predictability of movement has important implications for conservation management, such as allowing for specific management actions to maintain or protect populations during critical periods (Nemeth et al. 2006).
Our analyses also revealed different responses to flow by Australian grayling in different parts of the river system; fish in the upper reaches showed a more specific requirement for rising discharge to initiate migration than fish in the lower catchment. This is likely because fish in the lower catchment had already initiated downstream movement (fish were more likely to move on subsequent days if they have already begun moving), so flow change appears to become less important as a cue as fish move downstream. Indeed, fish in the lower reaches of the system were just as likely to move in response to negative flow changes as they were to positive flow changes. These findings suggest that environmental flow releases intended to initiate downstream migration to specific spawning areas may need to consider longitudinal variation in the migratory behaviour and requirements of resident fish. Consideration of spatial variation in behaviour may be particularly important for developing environmental flow recommendations in systems whose discharge can be regulated at several locations along the length of a river system. Although intraspecific variation in movement behaviour of fishes is an increasingly recognised phenomenon (e.g., Brodersen et al. 2008; Chapman et al. 2011), there has been little attempt to incorporate information on intra-specific variation into flow management of rivers. This is an important area for future research.
In conclusion, the empirical research and statistical modelling undertaken during this study enabled the prediction of the occurrence of fish spawning behaviour in relation to river discharge in a regulated river system. In particular, the study highlights how specific components of flows (in this case, the rising limb) and timing of flows can affect migration and spawning of fish. It also highlights the importance of considering spatial and temporal aspects of fish distributions and behaviour, and flow effects when implementing and adaptively managing environment flows. Applying this type of information to the development of targeted environmental flows to facilitate migration and spawning by diadromous fish could greatly improve the long-term viability of their populations and related fisheries.
References
Alluvium (2015) Macalister river environmental flows review. Alluvium, Melbourne
Amtstaetter F, O’Connor J, Pickworth A (2015) Environmental flow releases trigger spawning migrations by Australian grayling Prototroctes maraena, a threatened, diadromous fish. Aqua Conserv 26:35–43. doi:10.1002/aqc.2570
Arthington AH (2012) Environmental flows: Saving Rivers in the Third Millennium. University of California Press, California
Backhouse G, Jackson J, O’Connor J (2008) National recovery plan for the Australian Grayling Prototroctes maraena. Department of Sustainability and Environment, Melbourne
Berra TM (1982) Life history of the Australian grayling, Prototroctes maraena (Salmoniformes: Prototroctidae) in the Tambo River, Victoria. Copeia 198:795–805
Bestley S, Patterson TA, Hindell MA, Gunn JS (2010) Predicting feeding success in a migratory predator: integrating telemetry, environment, and modeling techniques. Ecol 91:2373–2384. doi:10.1890/08-2019.1
Bishop KA, Bell JD (1978) Aspects of the biology of the Australian Grayling Prototroctes maraena Günther (Pisces : Prototroctidae). Aust J Marine Freshwater Res 29:743–761. doi:10.1071/MF9780743
Boubée JA, Mitchell CP, Chisnall BL, West DW, Bowman EJ, Haro A (2001) Factors regulating the downstream migration of mature eels (Anguilla spp.) at Aniwhenua Dam, Bay of Plenty, New Zealand. NZ J Marine Freshwater Res 35:121–134. doi:10.1080/00288330.2001.9516982
Brodersen J, Nilsson PA, Hansson L-A, Skov C, Brönmark C (2008) Condition-dependent individual decision-making determines cyprinid partial migration. Ecol 89:1195–1200. doi:10.1890/07-1318.1
Bunn SE, Arthington AH (2002) Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ Manage 30:492–507. doi:10.1007/s00267-002-2737-0
Burnham KP, Anderson DR (2002) Model selection and inference: a practical information-theoretic approach. Springer-Verlag, New York, NY
Chapman BB, Brönmark C, Nilsson J-Å, Hansson L-A (2011) The ecology and evolution of partial migration. Oikos 120:1764–1775. doi:10.1111/j.1600-0706.2011.20131.x
Crook DA, Koster WM, Macdonald JI, Nicol SJ, Belcher CA, Dawson DR, O’Mahony DJ, Lovett D, Walker A, Bannam L (2010) Catadromous migrations by female tupong (Pseudaphritis urvillii) in coastal streams in Victoria, Australia. Marine Freshwater Res 61:474–483. doi:10.1071/MF09170
Crook DA, Macdonald JI, O’Connor JP, Barry B (2006) Use of otolith chemistry to examine patterns of diadromy in the threatened Australian grayling Prototroctes maraena. J Fish Biol 69:1330–1344. doi:10.1111/j.1095-8649.2006.01191.x
Durif CMF, Elie P (2008) Predicting downstream migration of silver eels in a large river catchment based on commercial fishery data. Fisheries Manag Ecol 15:127–137
Earth Tech (2006) Environmental flow determination for the Bunyip and Tarago Rivers - issues paper. Earth Tech Engineering, Melbourne
Freeman MC, Bowen ZH, Bovee KD, Irwin ER (2001) Flow and habitat effects on juvenile fish abundance in natural and altered flow regimes. Ecol Appl 11:179–190. doi:10.1890/1051-0761(2001)011[0179:faheoj]2.0.co;2
Godinho A, Kynard B, Godinho H (2007) Migration and spawning of female surubim (Pseudoplatystoma corruscans, Pimelodidae) in the São Francisco river, Brazil. Env Biol Fish 80:421–433. doi:10.1007/s10641-006-9141-1
Growns I, James M (2005) Relationships between river flows and recreational catches of Australian bass. J Fish Biol 66:404–416. doi:10.1111/j.0022-1112.2005.00605.x
Hall DN, Harrington DJ (1989) Studies on the spawning and early life history of Australian Grayling, Prototroctes maraena Günther, in the Barwon River, Victoria. Arthur Rylah Institute for Environmental Research Technical Report Series No. 84. Department of Conservation, Forests and Land, Heidelberg, Victoria
Irving DB, Modde T (2000) Home-range fidelity and use of historic habitat by adult Colorado pikeminnow (Ptychocheilus lucius) in the White River, Colorado and Utah. West N Am Naturalist 60:16–25
Jacobs (2015) Moorabool River FLOWS study update. Jacobs Engineering Group, Bendigo
King A, Gawne B, Beesley L, Koehn J, Nielsen D, Price A (2015a) Improving ecological response monitoring of environmental flows. Environ Manage 55:991–1005. doi:10.1007/s00267-015-0456-6
King AJ, Gwinn DC, Tonkin Z, Mahoney J, Raymond S, Beesley L (2015b) Using abiotic drivers of fish spawning to inform environmental flow management. J Appl Ecol 53:34–43. doi:10.1111/1365-2664.12542
King AJ, Ward KA, O’Connor P, Green D, Tonkin Z, Mahoney J (2010) Adaptive management of an environmental watering event to enhance native fish spawning and recruitment. Freshwater Biol 55:17–31. doi:10.1111/j.1365-2427.2009.02178.x
Koehn JD, King AJ, Beesley L, Copeland C, Zampatti BP, Mallen-Cooper M (2014) Flows for native fish in the Murray-Darling Basin: lessons and considerations for future management. Ecol Manage Restor 15:40–50. doi:10.1111/emr.12091
Koehn JD, O’Connor WG (1990) Biological information for management of native freshwater fish in Victoria. Victorian Government Printing Office, Melbourne
Koster WM, Amtstaetter F, Dawson DR, Morrongiello JR, Reich P (2017) Provision of environmental flows promotes spawning of a nationally threatened diadromous fish. Marine Freshwater Res 68:159–166. doi:10.1071/MF15398
Koster WM, Dawson DR, Crook DA (2013) Downstream spawning migration by the amphidromous Australian grayling (Prototroctes maraena) in a coastal river in south-eastern Australia. Marine Freshwater Res 64:31–41. doi:10.1071/MF12196
Koster WM, Dawson DR, Liu C, Moloney PD, Crook DA, Thomson JR (2016) Influence of streamflow on spawning-related movements of golden perch Macquaria ambigua in south-eastern Australia. J Fish Biol 90:93–108. doi:10.1111/jfb.13160
Lytle DA, Poff NL (2004) Adaptation to natural flow regimes. Trends Ecol Evol 19:94–100. doi:10.1016/j.tree.2003.10.002
McMahon TA, Finlayson BL (2003) Droughts and anti-droughts: the low flow hydrology of Australian rivers. Freshwater Biol 48:1147–1160. doi:10.1046/j.1365-2427.2003.01098.x
Morrongiello JR, Crook DA, King AJ, Ramsey DSL, Brown P (2011) Impacts of drought and predicted effects of climate change on fish growth in temperate Australian lakes. Glob Change Biol 17:745–755. doi:10.1111/j.1365-2486.2010.02259.x
Murchie KJ, Hair KPE, Pullen CE, Redpath TD, Stephens HR, Cooke SJ (2008) Fish response to modified flow regimes in regulated rivers: research methods, effects and opportunities. River Res Appl 24:197–217. doi:10.1002/rra.1058
Nemeth RS, Blondeau J, Herzlieb S, Kadison E (2006) Spatial and temporal patterns of movement and migration at spawning aggregations of red hind, Epinephelus guttatus, in the U.S. Virgin Islands. Environ Biol Fish 78:365–381. doi:10.1007/s10641-006-9161-x
Northcote TG (1984) Mechanisms of fish migration in rivers. In: McCleave, J, Arnold, G, Dodson, J & Neill, W (eds) Mechanisms of migration in fishes, Plenum, New York, p 317–355
O’Connor JP, Mahoney JC (2004) Observations of ovarian involution in the Australian grayling (Prototroctes maraena). Ecol Freshwater Fish 13:70–73. doi:10.1111/j.0906-6691.2004.00020.x
Piper AT, Manes C, Siniscalchi F, Marion A, Wright RM, Kemp PS (2015) Response of seaward-migrating European eel (Anguilla anguilla) to manipulated flow fields. Proceedings of the Royal Society of London B: Biological Sciences 282:20151098. doi:10.1098/rspb.2015.1098
Poff NL, Zimmerman JKH (2010) Ecological responses to altered flow regimes: a literature review to inform the science and management of environmental flows. Freshwater Biol 55:194–205. doi:10.1111/j.1365-2427.2009.02272.x
R Development Core Team (2015) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Silva AT, Santos JM, Ferreira MT, Pinheiro AN, Katopodis C (2011) Effects of water velocity and turbulence on the behaviour of Iberian barbel (Luciobarbus bocagei, Steindachner 1864) in an experimental pool-type fishway. River ResAppl 27:360–373. doi:10.1002/rra.1363
SKM (2005) Determination of the minimum environmental water requirements for the Yarra River. Minimum environmental water requirement and complementary works recommendations. Sinclair Knight Merz, Melbourne
SKM (2012) Yarra River environmental flow study review. Flow recommendations report. Sinclair Knight Merz, Melbourne
SKM (2013) Review of the environmental flow recommendations for the Bunyip and Tarago rivers. Sinclair Knight Merz, Melbourne
Stewart-Koster B, Olden JD, Gido KB (2013) Quantifying flow–ecology relationships with functional linear models. Hydrol Sci J 59:629–644. doi:10.1080/02626667.2013.860231
Taylor MK, Cooke SJ (2012) Meta-analyses of the effects of river flow on fish movement and activity. Environ Rev 20:211–219. doi:10.1139/a2012-009
VEWH (2016) Seasonal Watering Plan 2016-17. Victorian Environmental Water Holder, Melbourne
Acknowledgements
Melbourne Water and the Department of Environment, Land, Water and Planning funded this study. Lauren Dodd, Renae Ayres, Mike Nicol, Graham Hackett, Jed Macdonald, Damien O’Mahony and Frank Amtstaetter assisted with field work or laboratory sorting. Thanks go to Justin O’Connor, David Meagher and two anonymous reviewers for constructive comments on an earlier version of this manuscript. This study was conducted under Victorian Flora and Fauna Guarantee Permit 10004353 and 10005451, Fisheries Victoria Research Permit RP-827 and ethics permits 07/20, 08/04, 11/11 and 10/28 (ARI Animal Ethics Committee).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no competing interests.
Electronic supplementary material
Rights and permissions
About this article

Cite this article
Koster, W.M., Crook, D.A., Dawson, D.R. et al. Predicting the Influence of Streamflow on Migration and Spawning of a Threatened Diadromous Fish, the Australian Grayling Prototroctes Maraena . Environmental Management 61, 443–453 (2018). https://doi.org/10.1007/s00267-017-0853-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-017-0853-0