
Abstract
We examined the effects of the Zemko Dam removal on the Eightmile River system in Salem, Connecticut, USA. The objective of this research was to quantify spatiotemporal variation in fish community composition in response to small dam removal. We sampled fish abundance over a 6-year period (2005–2010) to quantify changes in fish assemblages prior to dam removal, during drawdown, and for three years following dam removal. Fish population dynamics were examined above the dam, below the dam, and at two reference sites by indicator species analysis, mixed models, non-metric multidimensional scaling, and analysis of similarity. We observed significant shifts in fish relative abundance over time in response to dam removal. Changes in fish species composition were variable, and they occurred within 1 year of drawdown. A complete shift from lentic to lotic fishes failed to occur within 3 years after the dam was removed. However, we did observe increases in fluvial and transition (i.e., pool head, pool tail, or run) specialist fishes both upstream and downstream from the former dam site. Our results demonstrate the importance of dam removal for restoring river connectivity for fish movement. While the long-term effects of dam removal remain uncertain, we conclude that dam removals can have positive benefits on fish assemblages by enhancing river connectivity and fluvial habitat availability.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Dams dramatically alter river systems by creating physical barriers to water movement. Changes in the natural variation in flow regime as a consequence of river impoundment can result in many cascading effects including changes in the distribution of pool-riffle channels (Gregory et al. 1994), alteration of sedimentation and nutrient cycling (Humborg et al. 1997; Vorosmarty et al. 2003), and shifts in thermal profile (Olden and Naiman 2010; Lytle and Poff 2004; Magilligan and Nislow 2005). This, coupled with the restriction of species movement pathways and propagule dispersal abilities from damming (Ward and Stanford 1995; Raymond 1979; Agostinho et al. 2004), can spur dramatic changes in aquatic ecosystem structure and function (de Jalón and Sanchez 1994; Kinsolving and Bain 1993).
Even small dams, where water flows over the crest of the structure (i.e., run-of-river or overflow weirs), can have major impacts on riparian environmental conditions and community composition (Velinsky et al. 2006; Neil and Mazari 1993). There are more than 42,000 small, run-of-river dams in the United States, and most of them are concentrated in the East (Army Corps of Engineers U 2013). Many dams were built between the early 1700 s and the present to create backwater conditions in river channels as water intake sources for mills, industrial plants, and water reservoirs (Csiki and Rhoads 2010). While run-of-river dams continue to alter stream hydrology through the artificial creation of lentic, warm water environments, most dams are no longer used for their original purposes, and they now exist in various states of neglect or abandonment (Graf 2001; Johnson and Graber 2002; Bowman 2002).
Resource managers and communities are increasingly interested in small dam removal as a strategy for dealing with degraded structures to reduce the risk of flooding from dam failure and to restore the ecological integrity of aquatic ecosystems (Poff et al. 1997; Hart and Poff 2002; Gardner et al. 2013; Hogg et al. 2013). Yet, there is a lack of information about the ecological impacts of dam removal on river systems. Many river restoration projects are conducted without a scientific context (Wohl et al. 2005). Although a range of studies have quantified how dam removal impacts river hydrology and species composition, we still know very little about the biophysical consequences of removing dams (Grant 2001; Gregory et al. 2002).
Dam removal constitutes a geomorphic perturbation that converts a reservoir back into a river (Stanley and Doyle 2003). The effects of this disturbance pervade both up- and downstream. Dam removal lifts the constraints on species colonization, but it also stimulates notable shifts in erosional and depositional processes that influence water quality (Grant 2001; Burdick and Hightower 2006). Changes in community composition can occur almost immediately after dam removal (Stanley et al. 2002; Doyle et al. 2000; Pess et al. 2008), but the responses of riverine biota to dam removal are often unpredictable and site-specific (Kanehl et al. 1997; Gregory et al. 2002; Stanley et al. 2002).
A number of studies have characterized changes in river channeling and geomorphology in response to dam removal (Bednarek 2001; Gregory et al. 2002; Stanley and Doyle 2003), and several studies have also monitored changes in fish community composition in response to small dam removals for several years after dam drawdown (Kanehl et al. 1997; Burdick and Hightower 2006; Quist et al. 2005; Agostinho et al. 2004; Gardner et al. 2013; Hogg et al. 2013). Yet, the small number of studies on this topic and the site-specific responses of fishes to such perturbations signal the need to forge a solid peer-reviewed, scientific foundation for dam removal activities (Doyle et al. 2000) to increase understanding of the spatiotemporal responses of biological communities to dam management.
This study builds upon the growing body of multi-year research on the effects of small dam removal by examining changes in fish species composition as a result of a dam removal in a small New England stream. The objective of this study was to quantify the spatiotemporal variation in fish assemblages in response to the removal of the Zemko Dam in the east branch of Eightmile River System in Salem, Connecticut, USA. We evaluate fluctuations in fish species composition both above and below the Zemko Dam site and at two additional reference sites. We address three key research questions. How quickly do shifts in fish species composition occur after dam removal in the Eightmile system? How does fish species composition in the dam and reference sites fluctuate over time? Does the species composition of the dam sites shift toward that of the reference sites over time? The study spanned 6 years (2005–2010) and covered the period prior to dam removal (1 year), during drawdown (2 years), and after dam removal (3 years). We hypothesized that changes in fish species composition would occur immediately after dam drawdown, and that we would observe a shift in species composition in the previously dammed sites toward the composition of the reference sites.
Methods
Site Descriptions
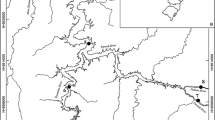
The Eightmile River is a 28-km-long tributary to the lower Connecticut River in southern New England (Fig. 1). It is situated in a relatively undeveloped, well-forested portion of southeastern Connecticut. The Eightmile River was designated a Wild and Scenic River by Congress in 2008, and a variety of local and state conservation groups have worked to protect the area since 1995 because of its unique geology, water quality, hydrology, and biota (Fosburgh et al. 2006). At the onset of this study, the East branch of the Eightmile had several obstacles to free fish passage, one of which was the Zemko dam. The two other remaining dams on the Eightmile River were Bill’s Dam (11 km downstream from the reference sites) and Hamburg Town Dam (10 km downstream from Bill’s Dam) (Fig. 1). Bill’s Dam contains a fish ladder for fish passage, and the Hamburg Town Dam is an overflow dam that does not entirely block fish passage, even during periods of low streamflow.

Map of the study Eightmile River study area and the sample sites included in the study with the locations of the remaining impoundments to streamflow following the Zemko Dam removal which was complete in October, 2007. Bill’s Dam contains a fish ladder, and the Hamburg Town Dam only partially blocks flow but not fish migration, even under low water flow conditions
The Zemko Dam was located approximately 0.75 km below the source of the Eightmile River. A dam for milling was first constructed there in the 1720’s and existed in various states of repair until the 1960’s, when the present-day dam was constructed. Since the original dam construction, the area above the dam has remained an open, lentic pool, while the reach below the dam has maintained characteristics of a small, free-flowing stream with heavy canopy cover. The reservoir behind the dam was periodically drained during the lifetime of the dam. At the time of removal, the dam was 1.5 m high, 3.7 m wide, and 24.5 m long stone- and earth-fill structure that also served as an unimproved road crossing. The center of the dam had a 5.6 m long spillway with an embedded rubble top. Dam drawdown and removal occurred over a 1-year period from summer 2006 to October 2007. Drawdown of the water level began by lowering the water level of the dam slowly over time by sequentially removing wooden “stoplogs” from the spillway. This drawdown step occurred over the course of a year, and the water level above the impoundment was lowered from 7 to 3 m deep during that time period. In October 2007, the base of the structure was completely removed.
Sample Site Descriptions
The Above-Dam Site
The above-dam site (41°29′41″N, 72°16′59″W) was characterized as a large, slow-flowing pond with heavy sediment loads prior to dam removal. The substrate was largely organic; this transitioned to silt and sand as the water levels decreased after the dam was removed, and the channel’s flow was restored. The stream bed and channel continued to experience changes throughout the study period. Water depth decreased from an average depth of 39.6 cm in 2005 to 34.8 cm in 2010, and water velocity increased after dam removal from a thalweg velocity of 0.19 m per second in 2005 to 0.33 m per second in 2010 based on depth and flow measurements taken in the middle of each reach at each sampling interval throughout the study. During later years of the study after dam removal, partially submerged cobble-sized rocks emerged at the upstream edge of the pond which partially resembled a natural riffle. This area was often blanketed in algae, and a recent tree fall impeded flow and trapped sediment toward the upstream end of the fishing reach. The river was surrounded by an emergent, wet meadow. Beyond the immediate open area, the site was bounded by woods on one side, and an old field on the other.
The Below-Dam Site
The below-dam site (41°29′34″N, 72°16′58″W) was characterized by a gravel and cobble bed with intermittent sand deposits in pools and along banks. This site contained a riffle at the end of a run with numerous pools and some boulders. This site was not deliberately altered during the drawdown and dam removal process, but this section of the river experienced major changes during this time period through the channel scouring and deposition of sand and silt in the pool directly below and downstream from the dam. Mean stream depth at the below-dam site decreased after dam removal from a depth of 14.1 cm in 2005 to 10.2 cm in 2010, likely in response to sediment transport from upstream. Thalweg water velocity slowed over the course of the study from a velocity of 0.5 s per meter in 2005 to 0.16 s per meter in 2010. In contrast to the open canopy established by the millpond above the dam, the reach below the dam had nearly continuous tree canopy cover by red maple (Acer rubrum Linnaeus), spicebush (Lindera benzoin L.), shagbark hickory (Carya ovata Mill.), and sweet pepperbush (Clethra alnifolia L.).
The Reference Sites
Two reference sites were included in the study: (i) one site 9.2 km downstream of the former dam in the eastern branch of the Eightmile River (41°26′31″N, 72°18′22″W); and a second site in the western branch (41°26′30″N, 72°19′59″W) (Fig. 1). The reference sites were chosen because they were relatively undeveloped and not obviously altered. Sampling in 2004 prior to this study confirmed, by a variety of water quality and ecosystem health metrics, that the site was relatively unimpaired by the effects of human activity (Chernoff, unpublished data). Stream depth and velocity did not fluctuate significantly over the course of the study. The sites were heavily wooded with yellow birch (Betula lenta L.), shagbark hickory (Carya ovata), witch hazel (Hammamelis virginiana L.), green ash (Fraxinus pennsylvanica Marsh.), and mountain laurel (Kalmia latifolia L.). The Eightmile west reference site was more open than the Eightmile east reference site, especially at the southern end of the site which abutted a heavily-vegetated steep bank. The reference sites lacked obvious alteration and showed little evidence of human impacts.
Fish Assemblage Surveys
We conducted monthly fish inventories between May and October from 2005 to 2010 above the dam site, below the dam site, and at the two reference sites. Fishes were collected for a 30-minute period during each of the sampling dates with a Smith-Root LR-24 backpack electroshocker (CT Dept. Env. Protection, Div. Nat. Res. permits SC-07014, SC-08022; Wesleyan University Animal Care and Use Protocol IACUC 20110225ChernoffA) and 0.16 cm stretch mesh nets. This timespan allowed for covering the entire sampling area, and all of the available habitats were sampled at each sampling interval. Some of the larger fishes were also inventoried by netting and identifying them in the water.
Statistical Analyses
Indicator Species Analysis
Fish relative abundance was calculated by species for each sampling interval at each site to make direct comparisons among sample sites over time. Relative abundance was calculated using count data for each species at each sampling interval. This value was expressed as the percentage abundance of each species relative to the total fish abundance for a sampling interval. We used indicator species analysis (Dufrene and Legendre 1997) and PC-Ord Software (McCune and Mefford 2011) to identify key indicator species across all years for the reference, above-dam, and below-dam sites. The goal of indicator species analysis is to identify species that occur in a particular habitat or location with high fidelity. The method combines species abundance data from a site and faithfulness of occurrence of a species in a particular site. The output identifies indicator species for each site based on a species’ faithfulness of occurrence and abundance in a particular site. Indicator values (ranging from 0 to 100) are simply estimated as the relative frequency of the species in sites belonging to a particular target site group. The fidelity of a species’ occurrence at one site (above-dam, below-dam, or reference site) is then tested with a Monte Carlo permutation which produces a P value indicating whether a particular species is a significant indicator of a particular site.
Species-Specific Trends
The relative abundances of each species’ were analyzed by site over time (2005–2010) via a mixed model procedure. We used R (R Development Core Team 2012) and the lme4 (De Boeck et al. 2011), languageR (Baayen 2007), and multilevel (Bliese 2006) packages to perform linear mixed effects analyses of the relationships among a species’ relative abundance, year, and sample site. The relative abundance data used in this modeling effort had many zeroes for species that were not encountered at certain sampling intervals. We chose not to transform the data prior to running the models because O’Hara and Kotze (2010) demonstrated that count data need not be transformed prior to subsequent analysis by linear models if the residuals are normally distributed. We included site and the interaction of site and year as fixed effects. Random effects were considered for the intercept, the slope of the site–year relationships, and for the sampling interval nested within year. The residuals of each model were inspected for deviations from homoscedasticity, and only models containing residuals without obvious deviations from normality were kept in our analysis. The final structure of the fixed effects for each model was selected by sequentially dropping non-significant terms from the full model, by measuring changes the significance of conditional F-tests for each term (Pinheiro and Bates 2000). The intra-class correlation (a measure of among-group site similarity) was also estimated for each model in order to assess the amount of variance in the response variable that can be attributed to the random effects in a model. The models describing the data most adequately were then selected by analysis of the Akaike information criterion (AIC) (Akaike 1974). The significance of individual sites and site-year combinations was assessed after final model selection via Markov-chain Monte Carlo methods (MCMC). MCMC is an iterative estimation technique which is used to build an empirical distribution of the multiple parameters of a linear mixed model.
Spatiotemporal Variation in Fish Community Structure
We used global non-metric multidimensional scaling (NMS) to summarize among-site similarities (Bray-Curtis distances) in fish assemblages over time in response to dam removal. NMS is an iterative search for the best positions of n entities on k axes that minimize the stress of the k-dimensional configuration. Stress is a measure of departure from monotonicity in the relationship between the distance in the original p-dimensional space and distance in the reduced k-dimensional ordination space (McCune et al. 2002). We used the slow and thorough autopilot mode of PC-ORD for NMS with the Sørenson (Bray–Curtis) distance metric in NMS space and a random starting configuration. We ran NMS for 250 iterations and with a final stability criterion of 0.0000001. Stress was evaluated using a Monte Carlo randomization procedure (P < 0.05) that was based on 500 randomized runs which indicated that a 3-dimensional solution was optimal, and that higher dimensional solutions did not result in significant reductions in stress.
We examined differences in fish community composition among sites and over time with Bray-Curtis similarity matrices and a two-way analysis of similarity (ANOSIM; α = 0.05; 999 permutations) using Past paleontological statistical software v2.17 (Hammer et al. 2001). Spatiotemporal change in community structure was further analyzed by performing a similarity percentage analysis (SIMPER) (Clarke 1993). SIMPER was used to identify major species contributing to >50 % of the total dissimilarity among sites for each year. This method computes the percentage contribution of each species to the dissimilarities between all pairs of sampling units (i.e., sampling intervals) in different sites. Species with a large contribution to dissimilarity between sampling units in different sites were those species that best discriminated among sites.
Results
A total of 23 species were captured during the course of the study (Table 1). The reference sites contained seven significant indicator species (Table 2), although the importance of brown trout in this site was artificial and not representative of physical habitat conditions because it was a non-native, invader species. Most of the species we encountered at the reference sites were fluvial, cool water specialists that were either moderately tolerant or intolerant of pollution (sensu Parasiewicz et al. 2007) (Table 1). The above-dam site hosted four significant indicator species which were mostly tolerant or moderately tolerant species that were cool or warm water specialists. The below-dam site had three significant indicator species, and all of these species were tolerant, warm water mesohabitat generalists.
The mixed model analysis identified significant spatiotemporal changes in fish relative abundance (Fig. 2). Brown bullhead, American eel, tessellated darter, redfin pickerel, bluegill, and fallfish displayed major fluctuations in abundance among sites and over time (P < 0.05). Brown bullhead densities in the reference sites were significantly lower (P = 0.001) than the above- and below-dam sites across all years of the study. Bullhead relative abundance spiked during dam drawdown, but then declined significantly (P < 0.05) after dam drawdown and removal in the above- and below-dam sites. Fallfish was abundant in the reference sites throughout the study, and its abundance increased significantly (P < 0.05) in both of the dam sites in the years following dam removal. However, only the smallest of juveniles (<0.22 mm length) were present in later years of the study in just the downstream end of the site; no larger juveniles, sub-adults, or adults were encountered. Tessellated darter relative abundance increased significantly following dam removal at the above- and below-dam sites, although it began to taper in 2010. The above- and below-dam sites displayed significant temporal fluctuations in American eel abundance (P < 0.05). Shifts in eel relative abundance were most noticeable in the above-dam site which experienced significant increases (P < 0.05) in eels after dam drawdown and removal. Relative abundance of redfin pickerels in the reference sites differed significantly from the other two sites (P = 0.0014). Year-to-year redfin pickerel variation was significant in the above-dam site, but pickerel population changes were not directional over the study period. Bluegill abundance at the below-dam site was significantly higher (P = 0.007) over the study period than the other two sites due to the influx (through colonization and/or recruitment) of this species in the years following dam removal, although a peak in abundance was also observed in 2009. Blacknose and longnose dace displayed significantly higher relative abundance at the reference sites relative to the two dam sites over the course of the study even though both dam sites experienced recruitment of dace in later sampling years.

Boxplots of fish relative abundance by site over time for species that displayed significant spatiotemporal fluctuations in relative abundance according to the mixed model procedure (i.e., species that displayed significant interactions for site and year). The box extends to ±1 SE, and the boxplot whiskers depict ±2 SE. Significant changes (P < 0.05) in species abundance by site and year according to Markov-chain Monte Carlo significance tests are depicted with an (*). The dam was intact in 2005. Dam drawdown occurred in 2006 and 2007, and the dam was completely removed for sampling intervals from 2008 on
The NMS corroborated the results from the mixed model analysis by demonstrating that all of the sample sites experienced shifts in species composition over time (Fig. 3). The temporal changes in species composition in the reference sites were much smaller than the two dam sites (Fig. 3d). The species composition at sites above and below the former dam remained relatively distinct from the reference sites over the course of the study even though the below-dam site displayed some overlap in species composition that was due to influxes and recruitment of juvenile fallfish, longnose dace, and blacknose dace in the later years of the study (Table 1; Fig. 3a–d). The NMS results also suggested that the species composition of the above- and below-dam sites was shifting toward the reference sites in the years after dam removal because of increases in fallfish and tessellated darter.

Non-metric multidimensional scaling of fish relative abundance showing site-level shifts in species composition from 2005 to 2010 (5 = 2005, 6 = 2006, 7 = 2007, 8 = 2008, 9 = 2009, and 10 = 2010) for the a reference sites, b above-dam site, c below-dam site, and d all sites displayed in relation to their distance from the site centroid and plotted with 95 % confidence ellipses for each site. Species names are displayed as four letter acronyms of the first two letters of the genus followed by the first two letters of the species as described in Table 2
The two-way ANOSIM results demonstrated that fish species composition differed significantly both among sites (P = 0.001, R = 0.71) and over time (P = 0.001, R = 0.36). The SIMPER results indicated that dissimilarity in species composition decreased over time both above and below the dam relative to the reference sites (Table 3). However, species’ dissimilarity remained relatively constant over time for the above-dam–below-dam sites comparisons. Prior to dam removal (2005), the reference and above-dam sites displayed the highest dissimilarity, while the above- and below-dam sites were the least dissimilar in species composition (Table 3). Brown bullhead and American eel were the two species that explained greater than 50 % of the differences in species composition in 2005. The sites displayed comparable differences in species composition during dam drawdown (2006). Brown bullhead and American eel remained important contributor species to the differences in species composition among sites during this time step. Tessellated darter was also an important species that was present in the reference sites, but not in the below-dam site in 2006. Species dissimilarity decreased after dam removal in 2007 for the reference–below–dam site and the above–below–dam site comparisons. In addition to bullhead and eel, fallfish appeared in this time step as a species that accounted for more than 50 % of the dissimilarity between reference–above–dam and reference–below–dam pairwise comparisons. In the years following dam removal, blacknose dace, longnose dace, and Tessellated Darter were the key contributors to the pairwise dissimilarities in species composition between sites. The SIMPER analysis indicated that the darter and both of the dace species were responsible for the dissimilarity among sites in the later years after dam removal. Fallfish recruitment after dam removal also contributed to the dissimilarity in species composition among sites in 2010.
Discussion
The East Branch of the Eightmile River experienced significant spatiotemporal changes in fish community composition in response to the removal of the Zemko Dam. However, our results suggest that this is still a river in transition, even 4 years after dam removal. The shifts likely occurred in response to alterations in physical habitat over time including substrate composition, water temperature, flow rate, and water depth, all of which have strong influences on fish assemblage structure (Gorman and Karr 1978; Rahel and Hubert 1991; Schlosser 1982). Shifts in fish species composition occurred rapidly (within 1 year of dam breach) at both the above- and below-dam sites. The ANOSIM results demonstrated that species composition was converging over time after dam removal which is a pattern that has been observed elsewhere (Vitule et al. 2012). However, major turnover in species composition from lentic to lotic fishes failed to occur within three years of dam removal as evidenced by the NMS results. Both the rapid shift in fish relative abundance and the lack of total recovery in our study system were consistent with other recent small dam removal studies (Catalano et al. 2007; Maloney et al. 2008), but the lack of a lentic to lotic fish assemblage change is likely a reflection of the wide tolerance by these species of a range of environmental conditions. This suggests that, while barrier removal facilitates fish colonization of new sites, the generation of physical habitats that can support lotic, cold water species may take more time, or may never occur.
Although some researchers have observed more pronounced downstream effects from dam removal (Maloney et al. 2008; Bushaw-Newton et al. 2002), both the above- and below-dam sites in the Eightmile River were significantly affected by dam removal both over time and relative to the reference sites. Many researchers have identified an immediate shift from lentic to lotic species dominance after small dam removal. Fish recruitment patterns into the former Zemko Dam site were more variable. Several fluvial and transition (i.e., pool head, pool tail, or run) species (sensu Kanno and Vokoun 2008) increased in abundance following dam removal, which could indicate river recovery. However, increases in warm water and pool specialists also occurred for the three years after dam removal. While a shift toward fluvial specialist species is often expected in response to dam removal, the pattern we observed may be related to the mixed-water fish assemblages of southern New England. Connecticut Rivers contain thermal gradients that transition from warm water to cold water within watersheds. Rapid turnover in stream temperature over short geographical distances has resulted in the dominance of mixed-water fish assemblages in smaller streams of this region (Kanno et al. 2010; Kanno and Vokoun 2008).
Shifts in Fish Relative Abundance
The movement of some species into new habitats either through colonization or recruitment of small individuals into new sites suggests that certain gregarious species moved quickly into other sites, while other more site-sensitive species have yet to experience the right conditions in these newly available habitats for survival and reproduction. Fallfish, tessellated darter, brown bullhead, and redfin pickerel displayed notable shifts in fish relative abundance over the course of the study. While fallfish is a pool specialist species in many locations (Fowler and Harp 1974), it is a fluvial specialist in Connecticut (Bain and Meixler 2000), and it is considered an indicator of healthy Connecticut streams (Bellucci et al. 2011; Bain and Meixler 2000). The recruitment of small juveniles of this species into the downstream edge of the Zemko site after dam removal provides some evidence of stream recovery, although the presence of only juveniles and no larger fish may also signal that small fish are dispersing to the site, but not staying there to reproduce despite the prevalence of prey. Fallfish are mesohabitat generalists, so the recruitment of this species following dam removal may simply be the product of increased habitat availability in later years of the study. Brown bullhead relative abundance above the dam declined after an initial spike during dam drawdown, as would be expected at the dam site due to its preference of warm water, lentic river conditions (Scott and Crossman 1973). Tessellated darter increased in abundance in response to dam removal, which is consistent with its preference of transitional habitats (Finger 1982) (i.e., points of change in water flow from pools to riffles as defined by Coates et al. (2007)). Tessellated darters are found across a range of river habitats including deeper waters and larger, depositional substrates in some locations (van Snik Gray and Stauffer 2001; Henry and Grossman 2008), and both pools and riffles in other areas (van Snik and Stauffer 1999). Variation in stream conditions following dam removal likely provided an array of potential habitats for darter occupation such as the human-placed partially submerged cobble-sized rocks at the upstream edge of the above-dam site toward the end of the study period, and increases in this species’ abundance following dam removal have been observed elsewhere (Stanley and Doyle 2003). Redfin pickerel was also abundant throughout the study period in the above-dam site, and it increased in abundance following dam drawdown at the below-dam site possibly due to a reduction in lateral flooding after dam removal or as a result of increases in abundance of downstream side water habitats after dam removal. Eastern Connecticut streams host high numbers of Redfin pickerel, and dam removal may have allowed this species to expand its range as habitat availability increased over the course of the study. This highlights the importance of dam removal for fish movement, and it contrasts other prior work by Helms et al. (2011) who found that Redfin pickerel remained largely upstream after small dam removal.
American eel, white sucker, and the dace species were affected to lesser extents by dam removal. American eel relative abundance fluctuated at all of the sample sites over the course of the study, as did white sucker populations in the two dam sites. Both dace species were significant indicator species of the reference sites which would be expected based on their reliance on riffle habitat (Finger 1982; Pizzuto 2002) and their prevalence in undisturbed Connecticut streams (Bellucci et al. 2011). However, the eventual increase in dace at the dam sites in 2010 suggested that fluvial specialist species could be used as a metric of stream recovery after dam removal due to increases in their abundance over time at the Zemko site over time.
River Connectivity
The shifts in species composition over time since dam removal at the two sites demonstrate the importance of dam removal for restoring river connectivity and promoting gene flow at the catchment scale (Fagan 2002; Stanford et al. 1996). While the need for river connectivity to maintain fish metapopulation dynamics is mainly documented for salmonids (McHenry and Pess 2008; Wilson 1999), releasing restrictions on fish movement through dam removal may also influence the population dynamics of other fishes as well by providing the potential for recolonization of habitats at sites where local extinction has occurred at above-dam sites (Rieman and McIntyre 1995; Fullerton et al. 2010). Shifts in fish species composition at the Zemko Dam site were gradual, which indicates that the reassembly of fishes at this site will occur over years or even decades. Access to source populations through dam removal may enhance the population viability of rare fishes over time, especially in other similar rivers with thermal gradients that host mixed-water species assemblages. For example, Catalano et al. (2007) documented the recolonization of 10 species into upstream reaches that were previously inaccessible to downstream source populations. We similarly noted significant recruitment of Tessellated Darter and small Fallfish into the site above the dam after dam drawdown. Bill’s Pond, the only remaining downstream impoundment to the east branch of the Eightmile River, is slated to be removed in late 2014 (TNC, unpublished data), which could further enhance river connectivity and the movement of fishes to previously inaccessible upstream portions of the river.
Management Implications
Dams alter physical habitat structure and fragment once continuous river reaches, resulting in more lentic communities and truncated species distributions. Our multi-year monitoring effort corroborates other prior research on this topic, by demonstrating that dam removal can significantly influence fish abundance and distribution patterns. However, we also found that the response of fishes after dam removal can vary widely by year and by spatial location. While this multi-year study identified major short-term shifts in fish assemblage composition in response to small dam removal, the long-term response of fishes to the Zemko Dam removal remains unclear. This study represents a real-time experiment, and the ongoing effects of dam removal on fish species composition will likely continue to shift over time. The effects of dam removals can be highly site-specific, but in general, they appear to enhance river connectivity and fluvial habitat availability.
References
Agostinho AA, Gomes LC, Verissimo S, Okada EK (2004) Flood regime, dam regulation and fish in the Upper Parana River: effects on assemblage attributes, reproduction and recruitment. Rev Fish Biol Fish 14(1):11–19. doi:10.1007/s11160-004-3551-y
Akaike H (1974) A new look at the statistical model identification. IEEE Trans Autom Control 19(6):716–723
Army Corps of Engineers U (2013) National inventory of dams. Federal Emergency Management Agency, New York
Baayen RH (2007) The languageR package. http://cran.r-project.org/web/packages/languageR/index.html
Bain MB, Meixler (2000) Defining a target fish community for planning and evaluating enhancement of the Quinebaug River in Massachusetts and Connecticut. Cooperative Fish and Wildlife Research Unit, Cornell University, New York
Bednarek AT (2001) Undamming rivers: a review of the ecological impacts of dam removal. Environ Manag 27(6):803–814
Bellucci CJ, Becker M, Beauchene M (2011) Characteristics of macroinvertebrate and fish communities from 30 least disturbed small streams in Connecticut. Northeast Nat 18(4):411–444
Bliese P (2006) Multilevel modeling in R (2.2). A brief introduction to R, the multilevel package and the nlme package. Walter Reed Army Institute of Research, Washington
Bowman MB (2002) Legal perspectives on dam removal. Bioscience 52(8):739–747
Burdick SM, Hightower JE (2006) Distribution of spawning activity by anadromous fishes in an Atlantic slope drainage after removal of a low-head dam. Trans Am Fish Soc 135(5):1290–1300
Bushaw-Newton KL, Hart DD, Pizzuto JE, Thomson JR, Egan J, Ashley JT, Johnson TE, Horwitz RJ, Keeley M, Lawrence J (2002) An integrative approach towards understanding ecological responses to dam removal: the Manatawny Creek Study. J Am Water Resour Assoc 38(6):1581–1599
Catalano MJ, Bozek MA, Pellett TD (2007) Effects of dam removal on fish assemblage structure and spatial distributions in the Baraboo River, Wisconsin. N Am J Fish Manag 27(2):519–530
Clarke KR (1993) Non-parametric multivariate analyses of changes in community structure. Aust J Ecol 18(1):117–143
Coates S, Waugh A, Anwar A, Robson M (2007) Efficacy of a multi-metric fish index as an analysis tool for the transitional fish component of the water framework directive. Mar Pollut Bull 55(1):225–240
Csiki S, Rhoads BL (2010) Hydraulic and geomorphological effects of run-of-river dams. Prog Phys Geogr 34(6):755–780
De Boeck P, Bakker M, Zwitser R, Nivard M, Hofman A, Tuerlinckx F, Partchev I (2011) The estimation of item response models with the lmer function from the lme4 package in R. J Stat Softw 39(12):1–28
de Jalón DG, Sanchez P (1994) Downstream effects of a new hydropower impoundment on macrophyte, macroinvertebrate, and fish communities. Regulat Rivers Res Manag 9(4):253–261. doi:10.1002/rrr.3450090406
Doyle MW, Stanley EH, Luebke MA, Harbor JM (2000) Dam removal: physical, biological, and societal considerations. In: American society of civil engineers joint conference on water resources engineering and water resources planning and management, Minneapolis, MN
Dufrene M, Legendre P (1997) Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol Monogr 67(3):345–366
Fagan WF (2002) Connectivity, fragmentation, and extinction risk in dendritic metapopulations. Ecology 83(12):3243–3249
Finger TR (1982) Fish community-habitat relations in a central New York stream. J Freshw Ecol 1(4):343–352
Fosburgh J, Case K, Hearne D (2006) Eigtmile wild and scenic study. National Park Service, Haddam, Connecticut
Fowler C, Harp G (1974) Ichthyofaunal diversification and distribution in Jane’s Creek watershed, Randolph County, Arkansas. Proc Ark Acad Sci 28:13–18
Fullerton A, Burnett K, Steel E, Flitcroft R, Pess G, Feist B, Torgersen C, Miller D, Sanderson B (2010) Hydrological connectivity for riverine fish: measurement challenges and research opportunities. Freshw Biol 55(11):2215–2237
Gardner C, Coghlan S, Zydlewski J, Saunders R (2013) Distribution and abundance of stream fishes in relation to barriers: implications for monitoring stream recovery after barrier removal. River Res Appl 29(1):65–78
Gorman OT, Karr JR (1978) Habitat structure and stream fish communities. Ecology 59(3):507–515
Graf WL (2001) Dam age control: restoring the physical integrity of America’s rivers. Ann Assoc Am Geogr 91(1):1–27
Grant G (2001) Dam removal: panacea or Pandora for rivers? Hydrol Process 15(8):1531–1532
Gregory KJ, Gurnell AM, Hill CT, Tooth S (1994) Stability of the pool riffle sequence in changing river channels. Regulat Rivers Res Manag 9(1):35–43. doi:10.1002/rrr.3450090104
Gregory S, Li H, Li J (2002) The conceptual basis for ecological responses to dam removal: resource managers face enormous challenges in assessing the consequences of removing large dams from rivers and evaluating management options. Bioscience 52(8):713–723
Hammer Ø, Harper DA, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron 4(1):9
Hart DD, Poff NL (2002) A special section on dam removal and river restoration. Bioscience 52(8):653–655
Helms BS, Werneke DC, Gangloff MM, Hartfield EE, Feminella JW (2011) The influence of low-head dams on fish assemblages in streams across Alabama. J N Am Benthol Soc 30(4):1095–1106
Henry BE, Grossman GD (2008) Microhabitat use by blackbanded (Percina nigrofasciata), turquoise (Etheostoma inscriptum), and tessellated (E. olmstedi) darters during drought in a Georgia piedmont stream. Environ Biol Fishes 83(2):171–182
Hogg R, Coghlan SM Jr, Zydlewski J (2013) Anadromous sea lampreys recolonize a Maine coastal river tributary after dam removal. Trans Am Fish Soc 142(5):1381–1394
Humborg C, Ittekkot V, Cociasu A, VonBodungen B (1997) Effect of Danube River dam on Black Sea biogeochemistry and ecosystem structure. Nature 386(6623):385–388. doi:10.1038/386385a0
Johnson SE, Graber BE (2002) Enlisting the social sciences in decisions about dam removal: the application of social science concepts and principles to public decisionmaking about whether to keep or remove dams may help achieve outcomes leading to sustainable ecosystems and other goals in the public interest. Bioscience 52(8):731–738
Kanehl PD, Lyons J, Nelson JE (1997) Changes in the habitat and fish community of the Milwaukee River, Wisconsin, following removal of the Woolen Mills Dam. N Am J Fish Manag 17(2):387–400
Kanno Y, Vokoun JC (2008) Biogeography of stream fishes in Connecticut: defining faunal regions and assemblage types. Northeast Nat 15(4):557–576
Kanno Y, Vokoun J, Beauchene M (2010) Development of dual fish multi-metric indices of biological condition for streams with characteristic thermal gradients and low species richness. Ecol Ind 10(3):565–571
Kinsolving AD, Bain MB (1993) Fish assemblage recovery along a riverine disturbance gradient. Ecol Appl 3(3):531–544. doi:10.2307/1941921
Lytle DA, Poff NL (2004) Adaptation to natural flow regimes. Trends Ecol Evol 19(2):94–100. doi:10.1016/j.tree.2003.10.002
Magilligan FJ, Nislow KH (2005) Changes in hydrologic regime by dams. Geomorphology 71(1–2):61–78. doi:10.1016/j.geomorph.2004.08.017
Maloney KO, Dodd H, Butler SE, Wahl DH (2008) Changes in macroinvertebrate and fish assemblages in a medium-sized river following a breach of a low-head dam. Freshw Biol 53(5):1055–1068
McCune B, Mefford MJ (2011) PC-ORD: Multivariate analysis of ecological data. MJM Software Designs, Gleneden Beach
McCune B, Grace JB, Urban DL (2002) Analysis of ecological communities, vol 28. MJM software design, Gleneden Beach
McHenry ML, Pess GR (2008) An overview of monitoring options for assessing the response of salmonids and their aquatic ecosystems in the Elwha River following dam removal. Northwest Sci 82(1):29–47
Neil D, Mazari R (1993) Sediment yield mapping using small dam sedimentation surveys, Southern Tablelands, New South Wales. Catena 20(1):13–25
O’Hara RB, Kotze DJ (2010) Do not log-transform count data. Methods Ecol Evol 1(2):118–122
Olden JD, Naiman RJ (2010) Incorporating thermal regimes into environmental flows assessments: modifying dam operations to restore freshwater ecosystem integrity. Freshw Biol 55(1):86–107. doi:10.1111/j.1365-2427.2009.02179.x
Parasiewicz P, Legros J, Rogers J, Wirth M, Jackson S, Kitson H, Deblois R, Hogue J, Brwaley H, VanAcker T, Longworth B, Schmidt R, Werle S, Thomas W, Dodge J (2007) Assessment and restoration of instream habitat for the Pomperaug, Nonnewaug and Weekeepeemee Rivers of Connecticut. Northeast Instream Habitat Program, University of Massachusetts, Amherst
Pess GR, McHenry ML, Beechie TJ, Davies J (2008) Biological impacts of the Elwha River dams and potential salmonid responses to dam removal. Northwest Sci 82(sp1):72–90
Pinheiro JC, Bates DM (2000) Mixed-effects models in S and S-PLUS. Springer, New York
Pizzuto J (2002) Effects of dam removal on river form and process. Bioscience 52(8):683–691
Poff NL, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BD, Sparks RE, Stromberg JC (1997) The natural flow regime. Bioscience 47(11):769–784
Quist MC, Hubert WA, Rahel FJ (2005) Fish assemblage structure following impoundment of a Great Plains river. West N Am Nat 65(1):53–63
R Development Core Team (2012) A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rahel FJ, Hubert WA (1991) Fish assemblages and habitat gradients in a Rocky Mountain-Great Plains stream: biotic zonation and additive patterns of community change. Trans Am Fish Soc 120(3):319–332
Raymond HL (1979) Effects of dams and impoundments on migrations of juvenile Chinook Salmon and Steelhead from the Snake river, 1966 to 1975. Trans Am Fish Soc 108(6):505–529
Rieman BE, McIntyre JD (1995) Occurrence of bull trout in naturally fragmented habitat patches of varied size. Trans Am Fish Soc 124(3):285–296
Schlosser IJ (1982) Fish community structure and function along two habitat gradients in a headwater stream. Ecol Monogr 52(4):395–414
Scott WB, Crossman EJ (1973) Freshwater fishes of Canada. Fish Res Board Can Bull 184:966
Stanford JA, Ward JV, Liss WJ, Frissell CA, Williams RN, Lichatowich JA, Coutant CC (1996) A general protocol for restoration of regulated rivers. Regul River 12:391–413
Stanley EH, Doyle MW (2003) Trading off: the ecological effects of dam removal. Front Ecol Environ 1(1):15–22
Stanley EH, Luebke MA, Doyle MW, Marshall DW (2002) Short-term changes in channel form and macroinvertebrate communities following low-head dam removal. J N Am Benthol Soc 21(1):172–187
van Snik Gray E, Stauffer JR (2001) Substrate choice by three species of darters (Teleostei: Percidae) in an artificial stream: effects of a nonnative species. J Inf 2001(1):254–261
van Snik Gray E, Stauffer JR Jr (1999) Comparative microhabitat use of ecologically similar benthic fishes. Environ Biol Fishes 56(4):443–453
Velinsky DJ, Bushaw-Newton KL, Kreeger DA, Johnson TE (2006) Effects of small dam removal on stream chemistry in southeastern Pennsylvania. J North Am Benthol Soc 25(3):569–582
Vitule JRS, Skóra F, Abilhoa V (2012) Homogenization of freshwater fish faunas after the elimination of a natural barrier by a dam in Neotropics. Divers Distrib 18(2):111–120
Vorosmarty CJ, Meybeck M, Fekete B, Sharma K, Green P, Syvitski JPM (2003) Anthropogenic sediment retention: major global impact from registered river impoundments. Glob Planet Change 39(1–2):169–190
Ward JV, Stanford JA (1995) Ecological connectivity in alluvial river ecosystems and its disruption by flow regulation. Regulat Rivers, Res Manag 11(1):105–119. doi:10.1002/rrr.3450110109
Wilson R (1999) Removing dam development to recover Columbia Basin treaty protected salmon economies. Am Indian Law Rev 24(2):357–419
Wohl E, Angermeier PL, Bledsoe B, Kondolf GM, MacDonnell L, Merritt DM, Palmer MA, Poff NL, Tarboton D (2005) River restoration. Water Resour Res 41(10):W10301
Acknowledgments
This project was completed using combined funds from Menakka and Essel Bailey, the Schumann Foundation, The Nature Conservancy, and project grants from Wesleyan University to BC and the College of the Environment. Student support and internships were provided by the Mellon Foundation, the Hughes Foundation, and Schumann Foundations. The authors thank Valerie Marinelli and Susan Lastrina for administrative and logistical support and Sarah Donelan for field assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Poulos, H.M., Miller, K.E., Kraczkowski, M.L. et al. Fish Assemblage Response to a Small Dam Removal in the Eightmile River System, Connecticut, USA. Environmental Management 54, 1090–1101 (2014). https://doi.org/10.1007/s00267-014-0314-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-014-0314-y