
Abstract
The estimation of optimum temperature of vegetation growth is very useful for a wide range of applications such as agriculture and climate change studies. Thermal conditions substantially affect vegetation growth. In this study, the normalized difference vegetation index (NDVI) and daily temperature data set from 1982 to 2006 for China were used to examine optimum temperature of vegetation growth. Based on a simple analysis of ecological amplitude and Shelford’s law of tolerance, a scientific framework for calculating the optimum temperature was constructed. The optimum temperature range and referenced optimum temperature (ROT) of terrestrial vegetation were obtained and explored over different eco-geographical regions of China. The results showed that the relationship between NDVI and air temperature was significant over almost all of China, indicating that terrestrial vegetation growth was closely related to thermal conditions. ROTs were different in various regions. The lowest ROT, about 7.0 °C, occurred in the Qinghai-Tibet Plateau, while the highest ROT, more than 22.0 °C, occurred in the middle and lower reaches of the Yangtze River and the Southern China region.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The optimum temperature for photosynthesis of terrestrial vegetation is the same as the optimum environmental temperature for vegetation growth. The estimation of optimum temperature of vegetation growth is very important for scientific research on both plant physiology and global changes, especially for some terrestrial ecological process models.
As a basic physiological process of vegetation growth, photosynthetic intensity depends on the difference between the actual temperature and the optimum temperature (Zhu 2005). Temperature, as a major factor determining the duration of the vegetation growing season, has been confirmed by many studies (Tucker and others 2001; Zhou and others 2003; Häninnen 1994). Appropriate temperature will increase the activity of enzymes and accelerate plant growth, until it reaches a maximum. By controlling the activity of enzymes, temperature effects the entire life cycle of vegetation (Berry and Bjorkman 1980; Chapin 1983). Vegetation net primary productivity is determined jointly by two processes, photosynthesis and respiration, both of which are affected by temperature. Generally, photosynthetic start and end time (onset and offset) are closely linked to temperature. Photosynthesis decreases and vegetative growth trends to decline when below or above this optimum temperature. On the other hand, respiration trends to respond to temperature exponentially. High air temperature probably reduced the values of net ecosystem exchange by increased respiration (Hunt and others 2004). Accumulated temperatures during the growing season can also influence the choice of cropping systems and their yield (Dong and others 2009). However, current research on estimating the optimum temperature of vegetation growth is usually undertaken in laboratories, such as controlled environment trials for single plant species. However, the heterogeneity of optimum temperature for vegetation growth is very large, due to scaling differences among single species, plant communities or ecosystems. Thus results from controlled environment studies for single species often not reflect the actual situation of vegetation growth in the field or at larger geographical scales.
The optimum temperature for vegetative growth is important in global change research. Several factors can profoundly alter vegetation growth status, such as water, soil, light, fire, and disease. Among these factors, temperature has been considered to be the major determinant of the length and growth of the growing season (Piao and others 2006; Wang and others 2005). As an input parameter of many phenological models and global carbon cycle models, temperature, especially the optimum temperature has influenced simulation accuracy and the final results to a large extent (Häninnen 1994; Rea and Eccel 2006; Liu 2008). Liu (2008) analyzed the influences of light use efficiency, photosynthetically active radiation, enhanced vegetation index, land surface water index, and temperature on gross primary productivity, to find that optimum temperature was the largest influencing factor. Accordingly, Wang and others (2005) found the NPP in some temperate zones was very sensitive to temperature. Also, a study result showed that the fraction of photosynthetically active radiation severely constrained by temperature, especially during the winter (Alcaraz-Segura and others 2009). However, current ecological models often simplify the mechanism of temperature requirements for growth and give simplified definitions of the optimum temperature. Consequently, parameter values used in studying the same ecosystem in the same region differ in the construction of different models. For example, Zhu (2005) defined the optimum temperature as the corresponding temperature when the normalized difference vegetation index (NDVI) reaches a maximum in a soil-vegetation process model. The optimum temperature in global production efficiency model (GLO-PEM) is defined as the long-term mean temperature for the growing season, based on the concept that plants grow efficiently at the prevailing temperature (Cao and others 2004). Besides, some researchers define the optimum temperature as the air temperature in the month when the NDVI reaches its maximum for the year (Los and others 1994). These different definitions will lead to a lack of comparability among the simulated results of different models and influence their reliability.
Vegetation growth of a plant community is the integrated result of the environment responses of plant species forming the community and various facets. Thus the distribution and growth of plant communities differ with geographic location and the growth conditions of those locations. Different vegetation types may require different temperature thresholds to trigger the green-up that marks the beginning of the growth season (Piao and others 2006). Researchers have found that the optimum temperature of some species, e.g., Notodanthonia penicillata, Poa caespitosa, could reach 27.0 °C, while the optimum temperature of other species, e.g., C. rigida might be as low as 9.0 °C. Even within species temperature responses of populations may differ according to the temperature conditions of the environments to which they are adapted. For example, Festuca novae-zelandiae populations from low latitudes had maximum temperatures for growth at 18.0 °C, but this dropped to 12.0 °C for populations from high latitudes (Scott 1970).
In order to characterize the large-scale macro characteristics of the optimum temperature for growth of communities and ecosystems, remote sensing provides a practicable approach. In this regard, the long time-series NDVI data set produced by Global Inventory Modeling and Mapping Studies (GIMMS) has been widely used to monitor vegetation growth and land surface characteristics dynamics (Myneni and others 1997; Julien and Sobrino 2009). For example, Schwartz and Chen (2002) combined plant phenology with NDVI to study the vegetation growing season in northern China, and found that NDVI is relatively stable as an indicator of the onset and offset of growing season. Chen and others (2005) and Piao and others (2006) studied the vegetation phenology characteristics and change trends in Chinese vegetation based on the NDVI data set.
Based on the above consideration, we used the GIMMS NDVI data set, the eco-geographical regionalization map of China, and temperature data to preliminarily estimate the optimum temperature for vegetation growth in different regions of China. The main objectives of this study were to: (1) examine the relationship between NDVI and temperature for verifying the feasibility of acquiring the optimum temperature for vegetation growth with NDVI, (2) investigate the internal relation of different definitions of the optimum temperature and phenological characteristics of vegetation, and (3) quantitatively provide and analyze the optimum temperature.
Data and Methods
Study Data
The GIMMS NDVI data set product is available for a 25-year period from 1982 to 2006 (http://glcf.umiacs.umd.edu/data/gimms/). It is derived from the imagery of the Advanced Very High Resolution Radiometer instrument on board the NOAA satellite series 7, 9, 11, 14, 16, and 17. The NDVI data set has a spatial resolution of 8 km × 8 km and a 15-day time interval. It has been corrected for calibration, view geometry, volcanic aerosols, and other effects not related to vegetation change.
The observational climatic data were provided by China meteorological data sharing service system. The daily climate data set was provided by 752 meteorological stations over China. This study selected the daily average temperature data, and made its time range correspond with the NDVI data set.
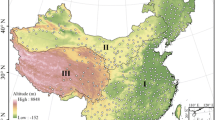
The Chinese eco-geographical regions data were obtained from a digitized 1:1,000,000 eco-geographical regions map, provided by the Institute of Geographical Sciences and Natural Resources Research, Chinese Academy of Sciences (Fig. 1) (Zheng and Li 2005).

Eco-geographical regions in China
Study Methods
In consideration of some unusual circumstances, e.g., NDVI is vulnerable to soil background information in regions with sparse vegetation cover (e.g., desert, arid lands, and frozen landscapes), this study defined that regions with multi-annual NDVI values less than 0.1 were desert regions, and this was confirmed by consulting the China land use/land cover (LULC) data set of 2000 (Liu and others 2005). We processed these desert regions with a special step (step 4). All steps are detailed below (Fig. 2):

Flowchart of procedures for defining temperature influences on the seasonal pattern of vegetation growth. G mni respects the 25-year average half months NDVI data set. C mti respects the 25-year average half months temperature data set. T(maxNDVI) is the time when NDVI value reaches maximum. T(maxRAT) is the time when maximum NDVI growth rate appear. T(GSSET) describes the duration time of growing season, from the start time to the end time [T(onset) / T(offset)]. GT(maxNDVI), GT(maxRAT), and GT(GSSET) are the time data after a threshold process (0.1). Tem(maxNDVI), Tem(maxRAT), and Tem(GSSET) [onset(tem) / offset(tem)] represent the temperature data corresponding with time data above
-
Step 1 the Savitsky–Golay smoothing filter was used to obtain smooth NDVI curves (Jönsson and Eklundh 2004). After this step, the 25-year half month data set (G n) was averaged to 24 half monthly NDVI time-series (G mni);
-
Step 2 for regions of max(G mni) ≥0.1, the corresponding time of maximum NDVI appearance [T(maxNDVI)] was directly calculated;
-
Step 3 processed the regions with max(G mni) ≥0.1. For these regions, the time when maximum NDVI growth rate (maxRAT) appears was calculated by T(maxRAT) = max[d(NDVI) / d(time)]; the vegetation growing season was calculated by Normalized NDVI ratio = (NDVI − NDVImin)/(NDVImax − NDVImin) (White and others 1997), and the ranges of Normalized NDVI ratio are 0–1. Here we made reference to the results of related studies and set the NDVI ratio threshold to 0.5 to acquire the onset and offset of vegetation growing seasons (Zhang and others 2004; Piao and others 2006; Nagai and others 2010; Lacaze and others 2009). Also, this method effectively eliminated the disturbance of the cropping index. After the third step, we obtained the corresponding time of the maximal NDVI rate and growing season length [T(maxRAT) and T(GSSET)].
-
Step 4 we extracted NODATA regions with max(Gmni) <0.1, and then used the majority values (the value that appeared most often) of the T(maxNDVI) / T(maxRAT) / T(GSSET) in the same eco-geographic regions to replace the NODATA respectively. After this step, we acquired the corresponding times of maximum NDVI [GT(maxNDVI)], the maximum NDVI rate [GT(maxRAT)], and the growing seasons [GT(GSSET)] for different eco-geographic regions.
-
Step 5 the Anuspline interpolation method was used to interpolate the temperature data (Hutchinson 1989; Dong and others 2010). We obtained the same temporal resolution and spatial resolution with the NDVI data, C mti (i = 1, 2, 3, … 22, 23, 24).
-
Step 6 used the results of steps 2–5 to acquire the corresponding temperature data [Tem(maxNDVI), Tem(maxRAT), and Tem(GSSET)]. Based on these temperature data, we investigated the referenced optimum temperature (ROT) of vegetation growth.
In order to verify if the relationship between temperatures and NDVI meet the need for further analysis, correlation between C mti and C mni was calculated. Moreover, for the convenience of statistics and analysis (Liu and others 2003), we combined with eco-geographical regions and divided into seven vegetation-climate regions, and each region had specific climate characteristics (Table 1).
Results and Analysis
The Relationship Between NDVI and Temperature
This study estimated the influence of temperature on vegetation growth based on the relationship between the intra-annual change of NDVI and corresponding temperature. The correlation analysis and significance test were done using the half-month average NDVI and temperature data. A statistically positive relationship was found in most vegetation-climate regions (Fig. 3).

The correlation coefficients R and R adj between 24 half-month NDVI and 24 half-month temperatures for areas of China. a Without elimination of the influence of no vegetation coverage (R). b Adjusted correlation coefficient (R adj) between temperature and NDVI that had been processed in step 4
In order to restore the true relationship of vegetation and temperature in no-vegetation or sparsely vegetated areas, we assumed that the adaptability of vegetation to temperature conditions was similar within the same eco-geographic region, i.e., particular eco-geographic regions were consistent with a particular climate and ecosystems. Here, we calculated the correlation coefficient (R) between mean NDVI and temperature in all eco-geographic regions, and then calculated the correlation coefficient (R adj) between temperature and adjusted NDVI after the processing of step 4.
The correlation coefficients in no-vegetation regions improved significantly after the processes of step 4. The regions of negative correlation or no correlation mainly occurred in arid regions such as Northwest China (NWC), in mountain humid areas such as several western parts of Eastern China and Southern China (EC and SC), and in alpine regions such as Qinghai-Tibet Plateau (TP) (Fig. 3). For the arid regions (e.g., Fig. 1, TP, HID1, and IID2 of NWC) an explanation was that their low-vegetation cover limits the spatial resolution of NDVI and it could not fully reflect the vegetation information. The mountain humid areas were mainly covered by evergreen forests in the Yunnan-Guizhou Plateau, evergreen rainforest in the southeastern area the Himalaya Mountains, and seasonal rainforest in Southern Yunnan medium altitude mountainous region (VA5, VA6, and VIIA3 regions). There were two main explanations for the smaller R values for these regions. The first was that the NDVI data set experiences more disturbance from terrain in mountainous regions (Fontana and others 2009) and this also applied to the alpine regions. The second explanation was that there are no clear and distinct seasons in the forested regions, so the vegetation change characteristics were not obvious with the temperature change. Further, these rainforest regions had the problem of NDVI super-saturation. However, the area percentage of no correlation or negative correlation decreased from 20.5 to 8.9 % after the adjustment of step 4, while the areas with statistically positive relationship between NDVI and temperature were over 90.0 % (Table 2). The above analysis illustrated that NDVI (or adjusted NDVI) can reflect the vegetation information. So, we can use the NDVI time series and the corresponding temperature data to express the relationships between vegetation growth and temperature.
Specific Growth Stages and Corresponding Temperature
T(maxNDVI) and Tem(maxNDVI) for Different Areas of China
The time and corresponding temperature when the NDVI value reached its maximum were shown in Fig. 4a, b, respectively. The times of maximum NDVI were mainly in summer (from June to August) and autumn (from September to November), covering 81.4 and 16.3 % of the area of China, respectively. The maximum NDVI in spring (from March to May) covered a small area (1.3 %). This phenomenon could be explained by the ephemeral and ephemerid plants characteristic of deserts. For this desert, there was a small period in spring when temperature and water availability allow growth. In winter (from December to February), it would be too cold for growth, but water would be accumulated in the soil (or snow), enough to allow growth in spring before it becomes too hot and dry for plants to survive in summer (Zhang and Chen 2002; Cui and others 2010). The small area (1 %) with T(maxNDVI) in winter was mainly located in tropical mountainous regions.

a The time of maximum NDVI [T(maxNDVI)] and b the temperature when this occurred [Tem(maxNDVI)] for different areas of China
Tem(maxNDVI) was higher in the southeast than the northwest of China, excluding the TP. The four humid and semi-humid regions, NEC, CC, EC, and SC, had mean Tem(maxNDVI) values of 20.6, 23.4, 23.0, and 22.7 °C, respectively. For the Inner Mongolia (IM), the mean Tem(maxNDVI) was close to 20.0 °C, reaching 19.4 °C. The mean Tem(maxNDVI) in the arid climate region (NWC) was 16.9 °C, which might reflect the effects of drought while the mean Tem(maxNDVI) in the TP region was 7.9 °C, which might relate to cold temperature stress on vegetation growth (White and others 1997; Jobbágy and others 2002).
The T(maxRAT) and Tem(maxRAT)
The times when the NDVI development rate was at maximum [T(maxRAT)] were mainly in spring and summer (Fig. 5a). Regions with T(maxRAT) in spring were the most extensive, covering 55.3 % of the total area of China. Regions with T(maxRAT) in summer covered 43.5 % of the area of China and were mainly located in the southwestern TP and the Taklimakan Desert regions (IIID1), the northern grasslands of IM (IID1 and IIC3) and southern Yunnan-Guizhou Plateau (VA5 and VIIA3).

a The time of maximum growth rate derived from NDVI [T(maxRAT)] and b the temperature when this occurred [Tem(maxRAT)] for different regions of China
When the NDVI increase rate reached a maximum, the photosynthesis rate of vegetation and growth rate of vegetation were also at their maximum (Fig. 5a). At this time (Fig. 5b), the mean Tem(maxRAT) of the SC region was the highest, reaching 22.0 °C; the mean Tem(maxRAT) of CC, EC, IM, and NWC regions ranged from 15.0 to 17.0 °C; the mean Tem(maxRAT) of NEC was 11.5 °C; and the mean Tem(maxRAT) of TP was 6.2 °C. By comparing the maxRAT of different climatic regions, we could find that it is directly related to the initial value of NDVI.
The Times and Temperatures for Vegetative Growth Season
The start of the growing season [Fig. 6a, T(onset)] is the time in a year when vegetation begins to grow after a period of dormancy, most usually after annual periods of winter cold or aridity in areas with seasonal rainfall. It is related to the time when the plants sprout from seeds (for annual plants) or the time when new leaves start or renew their development (for perennial plants). The end of the growing season [Fig. 6b, T(offset)] is defined as the date where the curve of the biophysical parameter of NDVI decreases below the value defined by step 3. T(onset) and T(offset) delineate the main period of the annual growth of vegetation with phases of accumulation of biomass by photosynthesis, use of synthesis for respiration, and loss of biomass by decomposition. The onset of the growing season in China was mainly in spring and summer (Fig. 6a). In most of the northern and southern regions of China, including NEC, CC, EC, and SC, growth began in spring. Although in some other regions, e.g., NWC, IM, and TP, vegetation growth was delayed due to low temperatures or shortage of water. T(offset) occurred in autumn for 81.8 % of the area of China (Fig. 6b). While the end of the growing season was in winter covering 13.9 % of the area, characteristically in tropical regions such as SC (VIA2, VIIA2, and VIIA3).

The times and temperatures of the onset and offset of the vegetation growing season (GS) in regions of China. a Onset of GS [T(onset)]. b Offset of GS [T(offset)]. c Temperature at onset of GS [Tem(onset)]. d The temperature at offset of GS [Tem(offset)]. e Mean temperature of GS [Tem(GSSET)]
The temperatures at onset [Tem(onset)) and offset (Tem(offset)] of the growing season were shown in Fig. 6c, d, respectively, and Fig. 6e shows the mean temperature of the growing season [mean Tem(GSSET)]. Tem(GSSET) (Fig. 6c) of the CC, EC, and SC regions was approximately 20.0 °C, for NEC, IM, and NWC is 15.0–17.0 °C, and for the TP region, it was the lowest at 6.1 °C. We could conclude from above analysis that the corresponding temperature of the vegetation growing season mainly fall into the range of temperature vegetation requires to complete its annual life cycle. Therefore, the growth season temperature could not be considered as the optimum temperature for vegetation growth.
The Optimum Temperature for Vegetation Growth
Although the optimum temperature for vegetation growth was generally described as the environmental temperature that corresponds with the most suitable temperature conditions for photosynthesis, some researchers had defined it differently when optimum temperature was a parameter in large scale research. There was consistency between the situation of vegetation growth and the intra-annual change of NDVI. Therefore, this study attempted to find the optimum temperature by analyzing the corresponding relationship between NDVI and temperature trends (Table 2).
The T(onset) and T(offset) of the vegetation growing season basically corresponded with the annual life cycle period of vegetation growth. It could reflect the process of plants sprouting, leaves growing, and biomass accumulation. That is, vegetation growth was within the range of the lower and upper limits of temperature at which growth could occur. Thus the temperature range and accumulation of degrees of temperature accumulation of the growing season needed to meet the temperature conditions that vegetation requires to complete its annual life cycle. We believed that the best situation of vegetation growth must occur in the growing season, and its optimum temperature also occurred during the growing season. So, optimum temperature could be estimated through processing the temperatures that occurred during the vegetation growing season (Fig. 7).

The relationship between the intra-annual change of NDVI and corresponding temperature
Vegetation growth rate accelerates from when the growing season began with the onset temperatures and temperatures continued to increase from this time. When the NDVI growth rate reached the maximum (maxRAT), photosynthesis attains its maximum rate. At the stage of T(maxRAT), there was rapid increase of leaf area, and vegetation changed from the reviving stage into the continually increasing phase (Gu and others 2005). Therefore, in theory, maxRAT and its corresponding time and temperature [T(maxRAT) and Tem(maxRAT)] were good indicators of vegetation growth. However, there were practical problems in estimating maxRAT, whose value was very vulnerable to the initial values. When the NDVI absolute value increased slightly, the NDVI growth rate would abruptly increase since the initial NDVI values were small. The T(maxRAT) determined by the maxRAT usually appeared in spring, and the corresponding temperature value of maxRAT [Tem(maxRAT)] was usually at a low level.
Along with the vegetation growth, NDVI growth rate might become slower, but NDVI would continue to increase until it reached its maximum value (maxNDVI). At this point, the vegetation reached its maturity phase, although the corresponding temperature may not be at the maximum. After that, branches and leaves turned yellow, accumulated biomass began to senesce and vegetation entered into the decreasing phase. It was assumed that T(maxNDVI) determined by maxNDVI was the time when gain of vegetation biomass through photosynthesis matched loss of biomass through respiration and decomposition, and the corresponding mean temperature of Tem(maxNDVI) always was chosen as the optimum temperature for vegetation growth. However, the optimum temperature was actually the temperature when the rate of vegetation biomass accumulation was the highest and not when the amount of standing biomass was the highest. T(maxNDVI) usually occurred in the summer or autumn, and its corresponding temperature Tem(maxNDVI) was usually at a higher level than the optimum temperature level. Then the vegetation stays in a zone of physiological stress was still away from the optimum temperature. Similarly, the midday depression of photosynthesis also indicated that the maximum temperature was not necessarily the optimum temperature for vegetation growth, though here we not referred to more plant physiology because of scale difference.
In summary, this study suggested that the upper limit of the optimum temperature should be the Tem(maxNDVI) and the lower limit should be determined by Tem(maxRAT). In order to quantitatively analyze optimum temperature, the upper and lower limit values were simply averaged to provide a ROT (Fig. 8).

ROT for vegetative growth in the climatic and eco-geographical regions of China
Discussion
The Applicability and Uncertainty of the ROT
Traditional records of the vegetation growth process were usually natural phenological observations for single species or communities, e.g., the Japanese and European phenology networks. The controlled environment experiments which considered the effects of light, temperature, water availability, and soil fertility factors on vegetation growth only can be done in a laboratory (Fitter and Fitter 2002). In recent decades, remote sensing had provided an appropriate and low-cost method to detect the large-scale vegetation growth process (Schwartz and Chen 2002; Piao and others 2006). However, the applicability of ROT in the analysis of vegetation growth process or modeling at large scale should be taken into account. Remote sensing was appropriately used to explore the vegetation change characteristics at ecosystem or regional scales.
One of presuppositions of the study was that the characteristics of vegetation types had not changed over the past 25 years, i.e., the vegetation composition was stable or the changes were just natural succession processes. Actually, vegetation composition was very complex. It would be influenced by environmental factors, human activities, and global climate change. The effects of this myriad of factors on inter-annual change were beyond the scope of consideration by the study. Further, the relatively sparse distribution of meteorological stations in some regions made the sample representativeness and effectiveness inadequate to some extent. Another presupposition was that the study period from 1982 to 2006 included almost all climate conditions, so it could represent the stable realistic status of vegetation. Precipitation and NDVI showed frequent fluctuations during the study period (Zhu and others 2011; Chen and others 2005). Precipitation had a very important influence on vegetation growth and composition (Li and others 2011), and water use efficiency and adaptive strategy of vegetation in arid and humid regions were very different. Even plants that grew in the same eco-geographical regions still had different responses to rain pulse events (Xu and Li 2006). This study focused on the characteristic temperature of vegetation growth. So synthesis analysis of long-term data was needed to explore complex realistic situations. Moreover, some studies had verified that, for particular humidity condition, the suitability of an environment for vegetation growth depended on the temperature situation (Li 2000; Pianka 1978). Therefore, the multi-annual NDVI and temperature data were used to explore ROT.
Although various environmental factors could influence NDVI values and the correlation, the results obtained were of practically useful and effective in current large-scale ecological modeling, and ROT could relate to actual vegetation growth condition well. Therefore, when simulation studies were done on a regional or global scale, the optimum temperature could meet the needs of mainstream ecological models.
The Geographical Distribution of the ROT
There was no obvious correlation between the ROT distribution and vegetation species distribution. On the contrary, the heterogeneity distribution of ROT reflected more the spatial variation of topography and environmental factors such as light, temperature, water, etc. The temperature conditions required by the same species in different habitats might be different, because the temperature requirements of species determined which environments they occurred in (White and others 1997; Jobbágy and others 2002). The temperature difference at the ecosystem scale was more obvious. Even same ecosystem at the same latitude, could have different optimum temperatures (Fig. 5).
Chapin (1983) summed up the optimum temperatures of different species occurring in different regions. The optimum temperature of vegetation in the Arctic Zone or cold temperate zone was approximately 15.0 °C, for the temperate zone it was about 25.0 °C, and for desert zones it could reach 35.0–45.0 °C. The ROTs based on the China eco-geographic regions (Table 1) were basically coincident with Chapin’s results, but some values were a little low, especially in some desert regions (IIID1, IID2, and IID3 of NWC, etc.). A possible reason was that the optimum temperature in desert regions was the optimum temperature of oasis species in the same eco-geographic region and not desert species after process of step 4. Also, the ROT of the TP was very low, which might reflect the influence of the low temperatures of high elevation. According to the result based on Shelford’s law of tolerance, the optimum temperature of vegetation usually occurred at 30–35 °C when the relative humidity (RH) was more than 85 %. Optimum temperature values decrease as RH would decrease, and RH was usually less than 60 % in most regions of China. Otherwise, Wang and others (2005) found that the temperature of tropic zone in China was outside of the optimum range for NPP since vegetations must provide more energy to support respiration at high temperatures. All those were theoretical basis of this study. Therefore, the lower ROT values might be appropriate.
Conclusions
In order to assess the potential of estimation optimum temperature, the spatial relationship between NDVI and the corresponding temperature in eco-geographical regions of China was analyzed using GIMMS NDVI data and climatic data for 25 years (from January 1982 to December 2006). Several phases of vegetation growth and corresponding temperatures were extracted. The study finally verified significant positive relationships between NDVI and temperature for 90 % of the area of China. The study also provided a ROT based on the characteristic temperatures of vegetation growth and the theoretical fluctuation range of the optimum temperature. ROT was obtained by systemic process of “NDVI fluctuate curve → some key growth phases of vegetation phenology → corresponding temperature → upper and lower limit temperature → ROT”. It was emphasized that ROT was the realistic ROT for actual plant communities. Comparing with studies of vegetation growth under controlled environment and experimental conditions, ROT might have more practical significance for model simulation at larger regional scale.
The order of regional ROTs from high to low was SC, EC, CC, IM, NWC, NEC, and TP (Table 1). The distribution also was in accordance with the spatial distribution of air temperature, indicating the adaptation of vegetation growth in these regions was an evolution result over a long period. In some regions of southern China, without the precipitation limitation, vegetation grew well under a high temperature conditions above 25.0 °C, whereas in the Qinghai-Tibet Plateau, vegetation also survived and grew well under temperature less than 10.0 °C. Although the hypothesis for calculating ROT was consistent with theoretical analysis of defining optimum temperature, measurement of the photosynthetic optimum temperatures of plant communities or dominant species, or validation to models with ROT will be required to verify the study results in future.
References
Alcaraz-Segura D, Cabello J, Paruelo JM, Delibes M (2009) Use of descriptors of ecosystem functioning for monitoring a national park network: a remote sensing approach. Environ Manage 43(38):48
Berry J, Bjorkman O (1980) Photosynthetic response and adaptation to temperature in higher plants. Annu Rev Plant Phys 31(1):491–543
Cao MK, Prince SD, Small J, Goetz S (2004) Remotely sensed interannual variations and trends in terrestrial net primary productivity 1981–2000. Ecosystems 7:233–242
Chapin FSI (1983) Direct and indirect effects of temperature on Arctic plants. Polar Biol 2:47–52
Chen XQ, Hu B, Yu R (2005) Spatial and temporal variation of phenological growing season and climate change impacts in temperate eastern China. Global Change Biol 11:1118–1130
Cui YP, Wang RH, Liu T, Zhang HF (2010) Extraction of vegetation information in arid desert ares based on spectral mixture analysis: a case in the western Gurbantunggut desert. J Desert Res 30(2):334–341
Dong JW, Liu JY, Tao FL, Xu XL, Wang JB (2009) Spatio-temporal changes in annual accumulated temperature in China and the effects on cropping systems, 1980s to 2000. Clim Res 40(1):37–48
Dong J, Liu J, Yan H, Tao F, Kuang W (2010) Spatio-temporal pattern and rationality of land reclamation and cropland abandonment in mid-eastern Inner Mongolia of China in 1990–2005. Environ Monit Assess 179:137–153. doi:10.1007/s10661-010-1724-9
Fitter A, Fitter R (2002) Rapid changes in flowering time in British plants. Science 296(5573):1689
Fontana F, Trishchenko AP, Khlopenkov KV, Luo Y, Wunderle S (2009) Impact of orthorectification and spatial sampling on maximum NDVI composite data in mountain regions. Remote Sens Environ 113(12):2701–2712
Gu Z, Chen J, Shi P, Xu M (2005) Correlation analysis of NDVI difference series and climate variables in Xinlingole steppe from 1983 to 1999. Acta Ecologica Sinica 29(5):753–765
Häninnen H (1994) Effects of climatic change on trees from cool and temperate regions: an ecophysiological approach to modeling of budburst phenology. Can J Bot 73:183–199
Hunt ER, Kelly RD, Smith WK, Fahnestock JT, Welker JM, Reiners WA (2004) Estimation of carbon sequestration by combining remote sensing and net ecosystem exchange data for northern mixed-grass prairie and sagebrush-steppe ecosystems. Environ Manage 33:432–441
Hutchinson M (1989) New objective method for spatial interpolation of meteorological variables from irregular networks applied to the estimation of monthly mean solar radiation, temperature, precipitation and windrun. Techn Memo 89(5):95–104
Jobbágy EG, Sala OE, Paruelo JM (2002) Patterns and controls of primary production in the Patagonian steppe: a remote sensing approach. Ecology 83(2):307–319
Jönsson P, Eklundh L (2004) TIMESAT–a program for analyzing time-series of satellite sensor data. Comput Geosci 30(8):833–845
Julien Y, Sobrino JA (2009) Global land surface phenology trends form GIMMS database. Int J Remote Sens 30(13):3495–3513
Lacaze R, Balsamob G, Baretc F, Bradleyd A, et al (2009) Geoland2-Towards an operational GMES land monitoring core service. First results of the biogeophysical parameter core mapping service. http://www.isprs.org/proceedings/XXXVIII/part7/b/pdf/354_XXXVIII-part7B.pdf
Li B (2000) Ecology. Higher Education Press, Beijing
Li L, Lu H, Campbell D, Ren H (2011) Methods for estimating the uncertainty in energy table-form models. Ecol Model 222:2615–2622
Liu M (2008) Study on estimation and uncertainty of terrestrial ecosystem productivity based on RS and GIS—take the grassland transect in Tibetan Plateau for example. Nanjing Normal University, Nanjing
Liu JY, Zhang ZX, Luo D, Xiao X (2003) Land-cover classification of China: integrated analysis of AVHRR imagery and geophysical data. Int J Remote Sens 24(12):2485–2500
Liu J, Liu M, Tian H, Zhuang D, Zhang Z, Zhang W, Tang X, Deng X (2005) Spatial and temporal patterns of China’s cropland during 1990–2000: an analysis based on Landsat TM data. Remote Sens Environ 98(4):442–456
Los SO, O JC, Tucker CJ (1994) A global 11 by 11 NDVI data set for climate studies derived from the GIMMS continental NDVI data. Int J Remote Sens 15:3493–3518
Myneni RB, Keeling CD, Tucker CJ et al (1997) Increased plant growth in the northern high latitudes from 1981 to 1991. Nature 386:698–702
Nagai S, Nasahara KN, Muraoka H, Akiyama T, Tsuchida S (2010) Field experiments to test the use of the normalized-difference vegetation index for phenology detection. Agric Forest Meteorol 150(2):152–160
Pianka E (1978) Evolutionary ecology. Harper and Row, New York
Piao SL, Fang JY, Zhou LM et al (2006) Variations in satellite-derived phenology in China’s temperate vegetation. Global Change Biol 12(4):672–685
Rea R, Eccel E (2006) Phenological models for blooming of apple in a mountainous region. Int J Biometeorol 51:1–16
Schwartz MD, Chen XQ (2002) Examining the onset of spring in China. Clim Res 12:157–164
Scott D (1970) Relative growth rates under controlled temperatures of some New Zealand indigenous and introduced grasses. N Z J Bot 8:76–81
Tucker CJ, Slayback DA, Pinzon JE et al (2001) Higher northern latitude normalized difference vegetation index and growing season trends from 1982 to 1999. Int J Biometeorol 45:184–190
Wang Q, Ni J, Tenhunen J (2005) Application of geographically-weighted regression analysis to estimate net primary production of Chinese forest ecosystem. Global Ecol Biogeogr 14:379–393
White MA, Thornton PE, Running SW (1997) A continental phenology model for monitoring vegetation responses to interannual climatic variability. Global Biogeochem Cycles 11(2):217–234
Xu H, Li Y (2006) Water-use strategy of three central Asian desert shrubs and their responses to rain pulse events. Plant Soil 285(1):5–17
Zhang LY, Chen CD (2002) On the general characteristics of plant diversity of Gurbantunggut sandy Desert. Acta Ecologica Sinica 22(11):1923–1932
Zhang X, Friedl MA, Schaaf CB, Strahler AH (2004) Climate controls on vegetation phenological patterns in northern mid-and high latitudes inferred from MODIS data. Global Change Biol 10(7):1133–1145
Zheng D, Li BY (2005) Chinese eco-geographic map. Beijing Commercial Press, Beijing
Zhou L, Kaufmann RK, Tian Y et al (2003) Relation between interannual variations in satellite measures of vegetation greenness and climate between 1982 and 1999. J Geophys Res 108(D1):4004. doi:10.1029/2002JD002510
Zhu W (2005) Estimation of net primary productivity of Chinese terrestrial vegetation based on remote sensing and its relationship with global climate change. Beijing Normal University, Beijing
Zhu WQ, Tian HQ, Xu XF, Pan YZ, Chen GS, Lin WP (2011) Extension of the growing season due to delayed autumn over mid and high latitudes in North America during 1982–2006. Global Ecol Biogeogr 21(2):260–271
Acknowledgements
This study was supported by the National Basic Research Program (2010CB950901 and 2012CB955800), National Natural Science Foundation of China (41171438), and Major Program of Humanities and Social Sciences Key Research Base of Ministry of Education of China (10JJDZONGHE015).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Cui, Y. Preliminary Estimation of the Realistic Optimum Temperature for Vegetation Growth in China. Environmental Management 52, 151–162 (2013). https://doi.org/10.1007/s00267-013-0065-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-013-0065-1