
Abstract
Sandy coastlines are sensitive ecosystems where human activities can have considerable negative impacts. In particular, trampling by beach visitors is a disturbance that affects dune vegetation both at the species and community level. In this study we assess the effects of the limitation of human trampling on dune vegetation in a coastal protected area of Central Italy. We compare plant species diversity in two recently fenced sectors with that of an unfenced area (and therefore subject to human trampling) using rarefaction curves and a diversity/dominance approach during a two year study period. Our results indicate that limiting human trampling seems to be a key factor in driving changes in the plant diversity of dune systems. In 2007 the regression lines of species abundance as a function of rank showed steep slopes and high Y-intercept values in all sectors, indicating a comparable level of stress and dominance across the entire study site. On the contrary, in 2009 the regression lines of the two fenced sectors clearly diverge from that of the open sector, showing less steep slopes. This change in the slopes of the tendency lines, evidenced by the diversity/dominance diagrams and related to an increase in species diversity, suggests the recovery of plant communities in the two fences between 2007 and 2009. In general, plant communities subject to trampling tended to be poorer in species and less structured, since only dominant and tolerant plant species persisted. Furthermore, limiting trampling appears to have produced positive changes in the dune vegetation assemblage after a period of only two years. These results are encouraging for the management of coastal dune systems. They highlight how a simple and cost-effective management strategy, based on passive recovery conservation measures (i.e., fence building), can be a quick (1–2 years) and effective method for improving and safeguarding the diversity of dune plant communities.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Coastal dunes are unique transitional ecosystems, and as such they are characterized by a highly specialized fauna and flora (Carter and others 1992; Martínez and Psuty 2004; McLachlan and Brown 2006). They provide nesting, foraging and stop-over areas for many species of vertebrates and invertebrates and harbour unique biological assemblages of plants (McLachlan and Brown 2006). However, their ecological functionality is presently at risk as human pressure on coastal zones has steadily increased over the last 50 years. This phenomenon is particularly striking in the Mediterranean and is largely due to beach tourism which attracts people from all of continental Europe (Curr and others 2000). Human activities have considerable negative impacts on sandy coastlines, ranging from the complete destruction of dunes systems in the face of urban sprawl, to the introduction of alien species for ornamental purpose and the disturbance of dune vegetation through trampling (McLachlan and Brown 2006). In Europe, recent regulations have meant that the majority of the remnant coastal dune ecosystems are now protected from the most destructive human activities (European Commission 1992; Baeyens and Martínez 2004). Nonetheless, these systems host large numbers of tourists, especially in the summer months, and therefore human trampling on dune vegetation still threatens both plant communities and sensitive animal species. Dune vegetation is an essential functional component of coastal dune ecosystems. Because of the stabilizing effect of vegetation on the substrate, its loss and degradation are known to lead to erosion by water and wind (Martínez and Psuty 2004; Barbier and others 2011). In addition to mitigating the loss of top soil and the effects of wind erosion, plant assemblages of dune complexes also offer other important ecosystem services, such as protection from sea storms (Everard and others 2010; Barbier and others 2011). Moreover, dune vegetation also provides an essential habitat for several animal taxa, like insects, gastropods, reptiles and birds (Verstrael 1996; McLachlan and Brown 2006).
Although dune plants can survive extreme conditions of drought and high temperature, they are particularly vulnerable to physical disturbance such as human trampling. In fact Andersen (1995), in a comparative study investigating the effect of different levels of tourism and trampling on several types of plant communities along the coast of Denmark, found that dune plant communities were more vulnerable to human trampling than grasslands and salt marshes. These results are further backed up by several other authors who have also highlighted how certain plant communities of the coastal dune zonation are more vulnerable than others to experimental human trampling (Bowles and Maun 1982; Lemauviel and Rozé 2003). Trampling directly affects individual plants through abrasion and can cause death or severely reduced fitness, especially in sandy soils (Van der Maarel 1971; Gallet and Rozé 2001). But human trampling also has indirect negative effects on coastal dune vegetation due to characteristics of the sandy substrate, which is highly susceptible of erosion (Doody 1989).
Although several authors have found that trampling strongly affects dune vegetation (Andersen 1995; Kutiel and others 2000; Lemauviel and Rozé 2003), to our knowledge the ability of these communities to recover and the time necessary for the recovery to take place (resilience) has yet to be accurately assessed. Resilience can be defined as the time required for an ecosystem to return to an equilibrium or steady-state following disturbance. It is an essential concept in restoration ecology, which is the intentional human activity that promotes or accelerates the recovery of an ecosystem with respect to its health, integrity and sustainability (Gunderson 2000; McCann 2000). Because of the functional importance of dune vegetation, its recovery following human disturbance is an important issue for the restoration of coastal dune ecosystems (Emery and Rudgers 2010). In this context, degraded sand dune ecosystems require a well planned set of targeted strategies and decisive action plans if the ecological functionality of these disturbed environments is to be restored and enhanced (Rozé and Lemauviel 2004). Moreover, and with specific reference to the present study, a better understanding of the patterns of response of vegetation to conservation actions in sandy coastal areas would allow the development of more appropriate site-based (i.e., focused on restoration at an ecosystem level) or target-based (i.e., conservation strategies for target species or assemblages) management strategies (Schlacher and others 2006).
Based on this, the aim of this study is to assess the effects of limiting human trampling on dune vegetation in a coastal protected area. We use a diachronic approach to compare plant community diversity and composition in sectors subject to different management strategies. Specifically, we compare plant community diversity in a recently fenced sector (which strongly limits trampling) with that of an open sector subject to trampling over the course of a two year period using rarefaction curves and a diversity/dominance approach. We focus on diversity changes because previous studies found that human trampling on dune vegetation specifically affects species diversity and richness (Kutiel and others 2000).
Methods
Study Area and Experimental Design
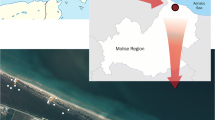
The study area (Torre Flavia wetland—Natural Monument) is located on the Tyrrhenian coast and designated as a “Special Area of Conservation” (European Commission 1979) (Fig. 1). The present wetland (about 43 ha) is a relict of a larger area that was recently drained and transformed (Battisti 2006). In the present study, habitat types were classified in accordance with the “Habitats Directive” (European Commission 1992) based on dominant and characteristic plant species (European Commission 2007; Biondi and others 2009).

Study area. The three dune sectors are indicated
Where land and sea meet, the area is characterized by a band of vegetation (Ceschin and Cancellieri 2006) that includes a number of typical habitats identified by the European Commission (2007) and by the Italian Ministry for the Environment (Biondi and others 2009). This coastal dune sector is longitudinally homogeneous. The embryo dune (Habitat 2110 of the Habitats Directive) characterized by the dominance of Elymus farctus and Anthemis maritima, is the most widespread habitat. Closer to the sea, along drift lines, there is a narrow annual vegetation strip dominated by Cakile maritima and Salsola kali (Habitat 1210). Landward of the embryo dune, the EC classification describes a true mobile dune community (Habitat 2120) dominated by Ammophila arenaria. However, this habitat is not present at Torre Flavia, although species belonging to this habitat do occur sporadically (i.e., Pancratium maritimum and Echinophora spinosa). The absence of the true mobile dune zone is probably due to the limited supply of sand from the sea, as the site is known to be undergoing erosion (Battisti and others 2008; Biondi and others 2009). The dune vegetation of the area can therefore be considered relative homogeneous and is represented mainly by Habitat 2110 (Embryonic shifting dunes) (Ceschin and Cancellieri 2006).
In the spring of 2007 the dune zone of the protected area was divided, in terms of different management schemes, into 3 sectors of equal extension (each sector measuring ~3.700 m2). To allow the nesting of two protected birds species (Charadrius dubius and Charadrius alexandrinus), and to reduce human trampling on dune vegetation, 2 of these 3 sectors were closed off by a fence (hereafter North fence and South fence). Fences were built using wooden poles separated by at least 1 meter between each pole. In this way the natural sand movement was not modified by the presence of the fence. At present they remain closed to the public. The third sector was kept open (free access to people and thus trampling) and was used as a control area (hereafter Open sector). The present study focuses on these 3 sectors (two trampled and one non-trampled) of the coastal dune system (Fig. 1).
In regards to trampling intensity during the study period, the number of visitors crossing each dune sector was reduced from 5–10 people per hour (pre-fencing) to <1/hour in the two fenced areas, while remaining constant in the open area (systematic observations by park managers). Trampling intensity rates were assessed during the beginning of the bathing season (April to June—at peak hours—10–12 a.m.).
Vegetation Sampling
In May 2007, immediately after the installation of the fencing systems, random vegetation sampling was undertaken in the three dune sectors (North fence, South fence, Open sector). The random placement of plots was achieved in a GIS environment (ArcGis 9.2). Firstly, polygons representing the position of the three dune sectors were layered onto orthophotos of the study area. Within each of the three polygons we then generated georeferenced random points and superimposed these onto our map (10 points in each sector, for a total of 30 points). Subsequently, the georeferenced points were identified in the field through the use of a Garmin GPS (GPSMAP®, 60CSx). In 2009, after two years of fencing, sampling was repeated at the same period of the year and at the same locations using the GPS unit. At each random point, we sampled all vascular plant species in a 2 × 2 m plot (4 m2), a size compatible with a visual estimation of vegetation cover (large enough for an appropriate visual estimation of cover—used here as a measure of abundance). The list of vascular plant species identified within each plot was recorded, together with the percentage of cover of each species using a 10%-interval rank scale. For each plot, the vegetation cover of all species present was visually estimated by three people. The estimation was performed by the same people in 2007 and in 2009. Taxonomy of plant species conforms to the most recent checklist of the Italian vascular flora (Conti and others 2005).
To avoid difficulties in relocating sampled plots we used an averaging procedure of position values over multiple readings (>100) to increase GPS precision, allowing us to accurately estimate plot position to the nearest 3–4 meters. The error associated with the GPS unit could not guarantee that we could return to exactly the same location when the second sampling program was carried out in 2009. However, given the sparse and homogeneous nature of the dune vegetation and the plot size used here, we are confident that possible small shifts in the patches surveyed in the two years would not critically affect results.
Data Analyses
Different approaches are available in applied ecology to study and compare the diversity of communities (Magurran 1988). Rarefaction curves are a widely used method for estimating standardized species diversity (Gotelli and Colwell 2001) and have been used effectively to describe coastal dune diversity patterns (Acosta and others 2009). The richness pattern of species was compared between the trampled sector (Open) and the two non-trampled sectors (North fence and South fence), by calculating sample based rarefaction curves for each of the three datasets (Gotelli and Colwell 2001), using the software EstimateS v. 7.5 (Colwell 2004). Expected species accumulation curves (sample-based rarefaction curves), with 95% confidence intervals, were constructed using the analytical formulas of Colwell and others (2004). The expected richness function is called Mao Tau and allows to represent the cumulative number of different species expected at each accumulation step—for details, formulas and interpretation see Colwell and others (2004). The 95% confidence intervals of the rarefaction curves (Ŝ Mao Tau) were used to determine whether species richness was significantly different among datasets (Colwell 2004).
Diversity/dominance analyses constitute a useful approach for understanding the changes that a community may undergo in response to a disturbance (Harrel and others 1967; Whittaker 1970; Battisti and others 2009). In particular, this method is commonly used to relate changes in community composition to shifts in stress levels (e.g., Whittaker 1960; Harrel and others 1967; Tokeshi 1993; Ghazoul 2002). For instance, the profile, trend, and shape of the diagram lines can shed light on anthropogenic or natural stresses affecting an assemblage (Ganis 1991; Fattorini 2005).
Diversity/dominance analyses were carried out using rank/abundance diagrams (Magurran 1988; Ganis 1991) based on relative cover (abundance) of species in each sector for both years. For each sector (10 plots in a single sector in a single year), the abundance of each plant species was calculated as its relative cover in relation to that of the total species cover in the 10 plots. To produce the rank/abundance diagrams, all species in a sector are first ranked from the most to the least relatively abundant (i.e., less frequent). Each species’ rank is then plotted on the X-axis against frequency on the Y-axis. The most abundant species is plotted first, followed by the second most common, and so on.
We fitted regression equations of abundance as a function of species rank using Monte Carlo permutations in the software EcoSim (Gotelli and Entsminger 2004) for each of the three sectors, both before fencing (2007) and then after trampling had been limited (2009). We then compared the Y-intercept and the slope of the three regressions amongst each other within each year. Assemblages strongly dominated by few species (highly sloped rank-abundance diagrams) are often present when strong disturbances have occurred, whereas low slopes may suggest a more stable and mature structure (Magurran 1988; Ganis 1991).
Results
Before the enclosures were put in place (2007), our results indicate that the plant communities of the three sectors were homogeneous. They all presented low and comparable values of species richness and the community structure was strongly skewed in favor of very few highly dominant species (see Appendix Table 2). Between 2007 and 2009, species richness increased markedly in the two enclosed sectors (North and South fence), where the total number of recorded species rose from 7–8 to 12. Furthermore, several typical dune species (Biondi and others 2009) were sampled only in the second time period: Limbarda crithmoides, Lagurus ovatus, Pancratium maritimum, Parapholis incurva, Silene canescens and Vulpia fasciculata. Conversely, in the Open sector species richness decreased slightly (7 to 6 recorded species) during the study period and two characteristic dune species (Biondi and others 2009) which had been recorded in 2007 were not found again in 2009 (Matthiola sinuata and Salsola kali) (see Appendix Table 2).
This pattern of increased species richness once fencing had been put in place is also highlighted by the rarefaction curves. In 2007, the rarefaction curves of the three sectors did not differ significantly in terms of the number of species recorded, as can be seen by the widely overlapping confidence intervals (Fig. 2a). In 2009, on the other hand, rarefaction curves show significant differences in the number of species recorded between the trampled sector (Open) and the two non-trampled areas (North fence and South fence). In terms of the cumulative species richness of all plots (Ŝ Mao Tau all species for m = 10), both fenced areas showed higher values of species richness when compared to the Open sector (Fig. 2b).

Rarefaction curves for the three sectors. In 2007 (a) the curves show no significant differences in the number of species recorded. In 2009 (b) the curves of the two non-trampled sectors show higher richness values if compared to the Open sector
As mentioned above, in 2007 the community structure of all three sectors was very similar and was characterized by the presence of a few highly dominant species (Anthemis maritima and Elymus farctus), accompanied by various less frequent and abundant species (Cutandia maritima and Chamaesyce peplis) (Appendix Table 2). As of 2009, the plant assemblage of the two fenced areas had become much more balanced, as evidenced by the more evenly distributed species frequencies. Instead, the plant assemblage of the Open sector remained very similar to that found in 2007 (Appendix Table 2).
This shift in community structure following enclosure was further supported by the diversity/dominance diagrams. Whereas the regression lines for each of three sectors in 2007 all showed high slope and Y-intercept values, indicating a high and comparable level of stress and dominance in the whole area (Fig. 3a; Table 1a), as of 2009 the picture was considerably different. The regression lines describing the two fenced areas (North fence and South fence) clearly separate from the trampled sector (Open). While the open area still shows a high slope and Y-intercept, the regression lines of the two non-trampled sectors are practically identical and both have much lower slopes and Y-intercepts (Fig. 3b; Table 1b).

Discussion
Both the results from the rarefaction curves and the diversity/dominance diagrams highlight how limiting human trampling can play a key role in reshaping diversity patterns in coastal dune plant communities. In fact, both methods indicate a recovery in plant community diversity and structure in the two fenced-off sites between 2007 and 2009, suggesting that limiting trampling can be effective in ameliorating conditions in dune habitats, even across short time periods. These results are consistent with other studies carried out in various communities subject to disturbance (Whittaker 1970; Tokeshi 1993; Ghazoul 2002; Battisti and others 2009), particularly in reference to the effects of trampling (Cole 1995; Whinam and Chilcott 1999; Hamberg and others 2010).
As mentioned above, various studies have shown dune vegetation to be particularly sensitive to human trampling (Andersen 1995; Kutiel and others 2000; Lemauviel and Rozé 2003). However, what is striking about the results of this study is that we were able to show how limiting trampling had a positive effect on dune vegetation in such a short time span. The rapid recovery of the dune plant communities could be related to the pioneer features that are typical of coastal dune plant species. In fact, several authors have highlighted that dune plant species are pre-adapted to strong natural stress and disturbances, such as wind and sea erosion, salt spray, sand burial etc. (Wilson and Sykes 1999; Perumal and Maun 2006). This pre-adaptation to natural disturbances is likely to also be an effective mechanism against human induced disturbances, allowing a fast improvement of general conditions of coastal dune communities once disturbance ceases or has been limited.
On the contrary the Open sector, which was subject not only to the factors of natural stress and disturbance which characterize the study area but also to sustained levels of human-induced disturbance, changed very little in terms of diversity and community structure during the two year study. Nonetheless, it should be noted that when comparing the results of the diversity/dominance analysis between 2007 and 2009, a slight improvement in community structure appears to have also taken place in the Open sector. Although not as clear as the changes in the fenced sectors, this raises the question of what underlying drivers are responsible for these observed patterns. In 2007 the entire dune area was suffering from particularly strong human disturbance, with visible signs of damage to both vegetation and soil (Battisti and others 2008). Since then, effective conservations measures have lead to a general improvement in the conditions of the Torre Flavia wetland. However, this increase in public awareness may be only part of the explanation of why conditions appear to have also improved in the Open sector. The variable nature of these sandy dune habitats means that inter-annual changes in community composition are common (Ward and others 2000; Elmendorf and Harrison 2009; Hassler and others 2010), as is evidenced in this specific case by the increase in cover across the entire study area of Cutandia maritima, an annual species (Appendix Table 2).
Given our finding, several important considerations regarding the management of costal dune areas should be made. Firstly, this study has shown how the use of fences or other measures that limit human trampling in dune systems can yield very encouraging results (see also Grafals-Soto and Nordstrom 2009). These measures are relatively simple to implement and maintain, and offer a cost effective solution for managing highly disturbed dune habitats. In addition to this, as the results of this study show, they can lead to a relatively quick recovery of dune vegetation communities. This simple strategy also requires little resource allocation (i.e., personnel, maintenance) over the period of the entire year, since trampling risk tends to be highly seasonal (i.e., late spring and early summer—from April to June). This season also coincides with the most delicate phenological period for most coastal dune plants, a further reason for concentrating management efforts during this time frame.
Furthermore, it is likely that in the specific case of this study part of the successful recovery of the dune vegetation can be attributed to efforts made at educating the visiting general public on some of the management and conservation issues that are important in coastal dune areas. This highlights the necessity of implementing management strategies that integrate direct conservation measures with the raising of public awareness. It also suggests that achieving goals in coastal dune conservation also requires changes in stakeholder attitudes (Nordstrom 2005).
In summary, the results of this study are encouraging for land managers working to improve the ecological functionality of coastal dune vegetation and dune ecosystems as a whole. They show that dune vegetation communities can be improved in relatively short time periods through the implementation of strategies that are effective in terms of both cost and management, and rely on the intrinsic recovery potential of these habitats.
References
Acosta A, Carranza ML, Izzi CF (2009) Are there habitats that contribute best to plant species diversity in coastal dunes? Biodiversity and Conservation 18:1087–1098
Andersen UV (1995) Resistance of Danish coastal vegetation types to human trampling. Biological Conservation 71:223–230
Baeyens G, Martínez ML (2004) Animal life on coastal dunes: from exploitation and prosecution to protection and monitoring. In: Martínez ML, Psuty NP (eds) Coastal dunes: ecology and conservation. Springer, Heidelberg, pp 279–296
Barbier EB, Hacker SD, Kennedy C, Koch EW, Stier AC, Silliman BR (2011) The value of estuarine and coastal ecosystem services. Ecological Monographs 81(2):169–193
Battisti C (ed) (2006) Biodiversità, gestione, conservazione di un’area umida del litorale tirrenico: la Palude di Torre Flavia. Gangemi Editore, Roma, p 496
Battisti C, Luiselli L, Pantano D, Teofili C (2008) On threats analysis approach applied to a Mediterranean remnant wetland: is the assessment of human-induced threats related into different level of expertise of respondents? Biodiversity and Conservation 16:1529–1542
Battisti C, Luiselli L, Frank B, Lorenzetti E (2009) Should fragment area reduction be considered a stress for forest bird assemblages? Evidence from diversity/dominance diagrams. Community Ecology 10:189–195
Biondi E, Blasi C, Burrascano S, Casavecchia S, Copiz R, Del Vico E, Galdenzi D, Gigante D, Lasen C, Spampinato G, Venanzoni R, Zivkovic L (2009) Italian Interpretation Manual of the 92/43/EEC Directive habitats. MATTM. Available from http://vnr.unipg.it/habitat/
Bowles JM, Maun MA (1982) A study of the effects of trampling on the vegetation of Lake Huron sand dunes at Pinery Provincial Park. Biological Conservation 24:273–283
Carter RWG, Curtis TGF, Skeffington MJ (eds) (1992) Coastal dunes: geomorphology, ecology and management for conservation. A.A. Balkema, Rotterdam
Ceschin S, Cancellieri L (2006) Inquadramento fitosociologico delle comunità vegetali nelle aree umide residuali del litorale nord della provincia di Roma. In: Battisti C (ed) Biodiversità, gestione, conservazione di un’area umida del litorale tirrenico: la Palude di Torre Flavia. Gangemi Editore, Roma, pp 164–168
Cole DN (1995) Experimental trampling of vegetation. I. Relationship between trampling intensity and vegetation response. Journal of Applied Ecology 32:203–214
Colwell RK (2004) EstimateS—statistical estimation of species richness and shared species from samples, Version 7.5. Available at http://viceroy.eeb.uconn.edu/estimates
Colwell RK, Mao CX, Chang J (2004) Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 85:2717–2727
Conti F, Abbate G, Alessandrini A, Blasi C (2005) An annotated checklist of the Italian vascular flora. Edizioni Palombi, Roma, p 420
Curr RHF, Koh A, Edwards E, Williams AT, Davies P (2000) Assessing anthropogenic impact on Mediterranean sand dunes from aerial digital photography. Journal of Coastal Conservation 6:15–22
Doody P (1989) Conservation and development of the coastal dunes in Great Britain. In: Van der Meulen F, Jungerius PD, Visser JH (eds) Perspectives in coastal dune management. SPB Academic Publishing, The Hague, pp 53–67
Elmendorf SC, Harrison SP (2009) Temporal variability and nestedness in California grassland species composition. Ecology 90:1492–1497
Emery SM, Rudgers JA (2010) Ecological assessment of dune restorations in the great lakes region. Restoration Ecology 18:184–194
European Commission (1979) Council Directive 79/409/EEC on the conservation of wild birds. Official Journal L 103:1–26
European Commission (1992) Council Directive 92/43/EEC on the conservation of natural habitats and of wild fauna and flora. Official Journal L 206:7–50 and its amending acts
European Commission (2007) Interpretation manual of European Union habitats, EUR 27. Available from http://ec.europa.eu/environment/nature/legislation/habitatsdirective
Everard M, Jones L, Watts B (2010) Have we neglected the societal importance of sand dunes? An ecosystem services perspective. Aquatic Conservation: Marine and Freshwater Ecosystems 20:476–487
Fattorini S (2005) A simple method to fit geometric series and broken stick models in community ecology and island biogeography. Acta Oecologica 28:199–205
Gallet S, Rozé F (2001) Resistance of Atlantic heathlands to trampling in Brittany (France): influence of vegetation type, season and weather conditions. Biological Conservation 97:189–198
Ganis P (1991) La diversità specifica nelle comunità ecologiche—Concetti, metodi e programmi di calcolo. Dipartimento di Biologia, Università di Trieste, C.E.T.A. (International Center of Theoretical and Applied Ecology), GEAD-EQ, n. 10
Ghazoul J (2002) Impact of logging on the richness and diversity of forest butterflies in a tropical dry forest in Thailand. Biodiversity and Conservation 11:521–541
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters 4:379–391
Gotelli NJ, Entsminger GL (2004) EcoSim: null models software for ecology. Version 7. Available at http://garyentsminger.com/ecosim/index.htm
Grafals-Soto R, Nordstrom K (2009) Sand fences in the coastal zone: intended and unintended effects. Environmental Management 44:420–429
Gunderson LH (2000) Ecological resilience—in theory and application. Annual Review in Ecology and Systematics 31:425–439
Hamberg L, Malmivaara-Lämsä M, Lehvävirta S, O’Hara RB, Kotze DJ (2010) Quantifying the effects of trampling and habitat edges on forest understory vegetation—a field experiment. Journal of Environmental Management. doi:10.1016/j.jenvman.2010.04.003
Harrel RC, Davies BJ, Dorris TC (1967) Stream order and species diversity of fishes in an intermittent Oklahoma stream. American Midland Naturalist 78:428–436
Hassler SK, Kreyling J, Beierkuhnlein C, Eisold J, Samimi C, Wagenseil H, Jentsch A (2010) Vegetation pattern divergence between dry and wet season in a semiarid savanna—Spatio-temporal dynamics of plant diversity in northwest Namibia. Journal of Arid Environments 74:1516–1524
Kutiel P, Eden E, Zhevelev Y (2000) Effect of experimental trampling and off-road motorcycle traffic on soil and vegetation of stabilized coastal dunes, Israel. Environmental Conservation 27:14–23
Lemauviel S, Rozé F (2003) Response of three plant communities to trampling in a sand dune system in Brittany (France). Environmental Management 31:227–235
McLachlan A, Brown AC (2006) The ecology of sandy shores. Academic Press, Burlington, p 373
Magurran A (1988) Measuring biological diversity. Blackwell, Oxford, p 256
Martínez ML, Psuty NP (2004) Coastal dunes. Ecology and conservation. Springer, Berlin, p 386
McCann KS (2000) The diversity–stability debate. Nature 405:228–233
Nordstrom KF (2005) Beach nourishment and coastal habitats: research needs to improve compatibility. Restoration Ecology 13:215–222
Perumal VJ, Maun MA (2006) Ecophysiological response of dune species to experimental burial under field and controlled conditions. Plant Ecology 184:89–104
Rozé F, Lemauviel S (2004) Sand dune restoration in North Brittany, France: a 10-year monitoring study. Restoration Ecology 12:29–35
Schlacher TA, Schoeman DS, Lastra M, Jones A, Dugan J, Scapini F, McLachlan A (2006) Neglected ecosystems bear the brunt of change. Ethology, Ecology & Evolution 18:349–351
Tokeshi M (1993) Species abundance patterns and community structure. Advances in Ecological Research 24:112–186
Van der Maarel E (1971) Plant species diversity in relation to management. In: Duffey E, Watt AS (eds) The scientific management of animal and plant communities for conservation. Blackwell Scientific Publications, Oxford, pp 45–63
Verstrael TJ (1996) Research on breeding birds in Dutch dune areas. Landscape and Urban Planning 34:301–313
Ward D, Saltz D, Olsvig-Whittaker L (2000) Distinguishing signal from noise: long-term studies of vegetation in Makhtesh Ramon erosion cirque, Negev desert, Israel. Plant Ecology 150:27–36
Whinam J, Chilcott N (1999) Impacts of trampling on alpine environments in central Tasmania. Journal of Environmental Management 57:205–220
Whittaker RH (1960) Vegetation of the Siskiyou mountains, Oregon and California. Ecological Monographs 30:279–338
Whittaker RH (1970) Communities and Ecosystems. Macmillian, New York
Wilson JB, Sykes MT (1999) Is zonation on coastal sand dunes determined primarily by sand burial or by salt spray? A test in New Zealand dunes. Ecology Letters 2:233–236
Acknowledgments
First and foremost, we would like to thank the park managers of the ‘‘Torre Flavia wetland’’ Natural Monument, funded by the Rome regional government, for allowing us to carry out the present study and for providing the systematic observations of trampling intensity. Further, we are indebted to the three anonymous referees for their extremely useful comments on the original manuscript which have allowed us to greatly improve our study. Last but not least, we thank Silvia Del Vecchio, Barbara Valentini and Elisa Bigi for their help with vegetation sampling in the field.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
See Table 2.
Rights and permissions
About this article
Cite this article
Santoro, R., Jucker, T., Prisco, I. et al. Effects of Trampling Limitation on Coastal Dune Plant Communities. Environmental Management 49, 534–542 (2012). https://doi.org/10.1007/s00267-012-9809-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-012-9809-6