
Abstract
Fallow vegetation within landscapes dominated by shifting cultivation represents a woody species pool of critical importance with considerable potential for biodiversity conservation. Here, through the analysis of factors that influence the early stages of fallow vegetation regrowth in two contrasting forest margin landscapes in Southern Cameroon, we assessed the impact of current trends of land use intensification and expansion of the cultivated areas, upon the conservation potential of shifting cultivation landscapes. We combined the analysis of plot and landscape scale factors and identified a complex set of variables that influence fallow regrowth processes in particular the characteristics of the agricultural matrix and the distance from forest. Overall we observed a decline in the fallow species pool, with composition becoming increasingly dominated by species adapted to recurrent disturbance. It is clear that without intervention and if present intensification trends continue, the potential of fallow vegetation to contribute to biodiversity conservation declines because of a reduced capacity, (1) to recover forest vegetation with anything like its original species composition, (2) to connect less disturbed forest patches for forest dependent organisms. Strategies to combat biodiversity loss, including promotion of agroforestry practices and the increase of old secondary forest cover, will need not only to operate at a landscape scale but also to be spatially explicit, reflecting the spatial pattern of species reservoirs and dispersal strategies and human usage across landscapes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Ranging from subsistence to fully market-oriented production, shifting cultivation systems assume a variety of forms but basically consist of the periodic spatial shift of cultivation to newly cleared forest or fallow land that is sufficiently fertile to support crop production (McGrath 1987). The continuous clearing of small patches of forests by millions of smallholder farmers (current estimates range from 35 million to almost a billion, Mertz 2009), makes shifting cultivation one of the major direct causes of tropical forest loss (FAO 2001). After a number of years of cultivation farmers rely on the regeneration of vegetation on the land left fallow to restore levels of nutrients and control pests and weeds (Burgers and others 2000). Beside fertility restoration, fallow vegetation is important because it provides food, beverage, fuel, building and medicinal products to local communities (Ambrose-Oji 2003, Toledo and others 2003). It may also mitigate habitat loss (Finegan and Nasi 2004, Kupfer and others 2006) and enhance carbon storage on deforested land (Kotto-Samè and others 1997, Palm and others 2005, Swallow and others 2007). Traditional shifting cultivation landscapes are considered less degraded than where forest conversion is to other more intensive and permanent agricultural land uses (Padoch and others 2005).
The general trend towards more intensive cultivation observed within shifting cultivation landscapes across the tropics (Mertz 2009) is expected to reduce their capacity to provide environmental services, including forest conservation. Studies of the effects of shifting cultivation on biodiversity, soil and water resources, suggest that this form of land use is sustainable where a sufficient fallow length is maintained.
Previous studies have analyzed the factors that influence secondary vegetation regrowth in early post-cultivation succession. Beside the age of the succession, studies point to field preparation and management practices: the last crop (Mitja and Hladik 1989, Ferguson and others 2003, Dalle and De Blois 2006), farming duration (Uhl 1987, Mitja and Hladik 1989, Chazdon 2003), fallow sequence and length of fallow (Lawrence 2004, Dalle and De Blois 2006). Others consider the vegetation type before last cropping, the occurrence of weeds such as Chromolaena odorata (De Rouw 1994, Ngobo and others 2004) and the occurrence of standing trees retained at the moment of field preparation (Carrière and others 2002). A few studies point to lateral flows from adjacent land cover units and seed dispersal processes (van Noordwijk 2002, Chazdon 2003, Lawrence 2004) highlighting the importance of the spatial arrangement of units within the landscape, particularly proximity of mature secondary vegetation (Lawrence 2004) and forest (Kupfer and others 2004, Hooper and others 2004, Makana and Thomas 2006). Very little knowledge exists about the combination of these factors and their interaction at a landscape scale.
The present study sought to assess the impact of combined agricultural expansion/deforestation and intensification processes on the potential for biodiversity conservation in shifting cultivation landscapes. The study was conducted in Southern Cameroon at the western margin of the 162 million ha of dense forest in Central Africa (de Wasseige and others 2009), the second largest remaining tract of humid tropical forest after the Amazon that is threatened by the continuous expansion of smallholder farming (Achard and others 2002).
Southern Cameroon is one of the “deforestation hot spots” (TREES 1998) in the region where continuous population growth, in particular in the urban areas has been combined with greater market access for small scale farmers at the forest margin, associated with improved transport infrastructure (Mertens and others 2000). This in turn has led to rapid conversion of forest into agricultural land, accompanied by a shift from the traditional long forest fallow to short rotational bush or tree fallows (Nounamo and Yemefack 2002). Short fallows (<5 years) are found in particular in areas with high population density as an adaptation to the scarcity of land and. However, even in land abundant areas, fields may be cultivated in short period fallow, because of the rapidly increasing labor requirement for clearing as fallow period lengthens (Gockowski and others 2004). Fallow shortening concerns in particular crops that are the traditional basis of household food-security such as groundnut and cassava. The increasingly market-oriented production of cucumber (Cucumeropsis m.), banana-plantain and oil palm affect almost exclusively old growth forest.
Our main hypothesis was that agricultural expansion and intensification based on fallow shortening, by modifying the spatial and temporal arrangement of forest and agricultural units within an area, affect fallow vegetation regrowth. We investigated the factors controlling the first 15 years of succession in fallow plots in two shifting cultivation landscapes with contrasting agricultural intensification processes in Southern Cameroon, through the analysis of woody species composition and abundance. We combined plot level and landscape level analysis to assess the relative importance of landscape scale processes, compared to the more widely studied plot level factors. In the present paper we discuss the implications of these results for landscape management to maintain forest conservation value in shifting cultivation landscapes.
Material and Methods
Sample Sites
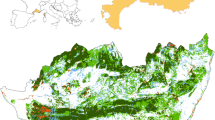
The present study was carried out in two contrasting sites within the Central African Forest Benchmark of the Alternatives to Slash and Burn program (ASB, www.asb.cgiar.org): Akok and Nkometou (Fig. 1). The benchmark encompassed a North-South gradient of population pressure, agricultural intensification and agriculturally driven deforestation (Gockowski and others 2004).

ASB forest margin benchmark (FMBA) in its regional context and showing detail of the sampled sites: Nkometou and Akok. Land cover maps for Central Africa and South Cameroon were modified from GLC2000. The vegetation map of Africa employs the following key: dark grey dense high forest, grey secondary forest and rural complex, light grey forest/savannah mosaic. The 1 km2 maps of the sampled sites (key shown) were produced by visual interpretation of Ikonos satellite images (Robiglio 2008)
The northernmost village, Nkometou (N 4.09944, E 11.53722), was close to Yaounde, and had an overall population density of 72 people km−2 and very good market access (Gockowski and others 2004). The agriculture/forest pattern was “diffuse” (TREES 1995) with the land cover types finely interspersed. The dominant forest type was the “extremely degraded semi-deciduous forest” (Letouzey 1985).
At the opposite extreme Akok (N 2.74167, E 11.23611) in the South, was relatively inaccessible and far from major markets, population density was 7 people km−2. The agriculture/forest pattern was “linear”, dominated by forest with interspersed cultivated fields and fallows lying along a forest track. The dominant forest type was the “degraded semi-deciduous mixed forest with prevailing elements of the evergreen forest” (Letouzey1985).
In the study area farmers combined food production from crops grown in shifting cultivation cycles with cocoa agroforests and more recently small oil-palm plantations.
Depending on its subsistence requirements each household cropped a minimum of two new food crop fields a year (Gockowski and others 2004) and maintained a set of fallow parcels under various successional stages. Fields and fallows were scattered throughout the village territory according to the spatial configuration of local land tenure (Robiglio 2008). Fallows were the dominant component of the rural mosaic.
Site selection for cropping was the result of a series of trade-offs that households faced at the beginning of each cropping season (Brown 2006). Basically, the fallow or forest land to be cleared was selected on the basis of the requirements of the crops that were going to be planted. This results at landscape scale in a complex rotational system that combines short and long fallows (Nounamo and Yemefack 2002). Fallow sequences were not replicated in a fixed manner (Robiglio 2008) and short and long fallows sometimes alternated on the same plot. Field shapes were irregular and their perimeters were not necessarily maintained across fallow cycles.
Farmers preferentially cropped fields close to those that they, or their close relatives, had cropped in previous seasons resulting in spatial clustering of their activities that made efficient use of their time (Robiglio 2008). Therefore, despite their small size (mean size of a mixed food crop field was 0.2 ha, std dev 1.7) the spatial aggregation of contiguous fields and fallows results in extensive areas of agricultural matrix.
On village land cover maps for 2001, derived from very high resolution Ikonos images (Fig. 1), we identified two dominant types of agricultural matrix reflecting a different degree of cultivation intensity. The short fallow matrix (herbaceous matrix) was dominated by young herbaceous fallows interspersed with clustered fields and scattered shrub/tree fallows. The long fallow matrix (shrubland matrix) was dominated by shrub/tree fallows with scattered cultivated fields and herbaceous fallows. In Nkometou a cultivated grassland matrix was also identified, with a mix of grassland and herbaceous fallows interspersed with clustered cultivated fields.
Vegetation Data
We sampled fallow vegetation in 180 plots located in two 1 km2 square windows per site (4 in total) characterized by a different landscape fragmentation pattern (Fig. 1d, e, f, g). In the agricultural matrix of each window, 30 fixed area sample plots (25 m × 25 m) were selected in a stratified sequence of fallow age and distance from the nearest forest margin. To reflect the effects of agricultural expansion and fallow shortening we focused only on the early stages of fallow vegetation regrowth, excluding traditional long forest fallow rotations not representative of such conditions.
Four classes of ‘age since plots were last cultivated’ were defined on the basis of existing literature on the ecology of fallow in similar zones of the country (Ngobo and others 2004, Ndam 1998): (1) very young fallow (0–3 years), (2) young fallow (4–7 years), (3) medium fallow (8–12 years) and (4) old fallow (13–16 years). For each age class a minimum of six plots at various distances from the forest were sampled. A further 15 control plots per window were selected within forest stands at a minimum distance of 50 m from the forest edge.
In each plot all trees and shrubs with dbh (stem diameter at breast height of 1.3 m) (5 cm, were identified to species level and counted. A general vegetation description was recorded and herbaceous and shrub vegetation cover was assessed by species according to the Braun-Blanquet Scale (1928, in Kent and Coker 1992).
The basic assumption for our study was that at each site, fallow samples in the two different windows, were derived from the same original forest types. We tested this applying a Multi-Response Permutation Procedure (MRPP) in PC-ORD 4 (McCune and Grace 2002). MRPP using the forest controls was done to test for any statistically significant separability amongst forest samples within the two 1 km2 windows at each site. Similarly we assessed the validity of the original separation of the samples in age classes by running an MRPP on age. We did the calculation using semi-metric Bray–Curtis distance on species abundance (McCune and Grace 2002). MRPP runs confirmed that the samples within each site could be treated as having been derived from the same forest types. The four age classes, however, were overlapping so we aggregated them in two groups: young fallow (yf) and old fallow (of) with plots from 1 to 7 and from 8 to 15 years in age, respectively, that were then separable.
Plot and Landscape Factors
Drawing on existing literature on post cultivation succession and fragmented landscapes (see the review in the “Introduction” section) we distinguished factors directly acting on plot characteristics from landscape level factors. Six plot level variables were identified and some grouped where they were associated with one another:
-
1.
Fallow age: the number of years since last cultivation;
-
2.
Land use: (a) last field type, (b) number of previous fallows and (c) vegetation type before last cropping;
-
3.
Weed layer: percentage cover of Chromolaena odorata, an exotic invasive species that tends to dominate fallows;
-
4.
Un-felled trees: density of trees retained during field preparation.
Information for 1 and 2 was established by interviewing the farmers directly on the spot. Farmers were asked to describe the recent evolution of fallow vegetation and to trace back the sequence of fallows and cropping cycles. To facilitate the task in each village we used local fallow classification categories previously identified through focus group work in the two villages (Robiglio 2008).
In addition, five landscape scale variables were measured and again grouped where related to one another:
-
1.
Fragmentation pattern: dominance of forest or agricultural classes in the 1 km2 sample window;
-
2.
Land cover: class of the polygon containing the sample plot on the 2001 land cover maps;
-
3.
Forest effect: (a) minimum distance of the plot centre to the nearest forest, (b) percentage of forest in an area delimited by a circle of radius 100 m from the plot centre;
-
4.
Frequency of vegetation with trees in reproductive status along plot boundaries (hereafter mature vegetation).
Mature vegetation included fallow vegetation at advanced successional stage, cocoa agroforests and remnants of un-slashed primary or secondary vegetation.
Landscape variables 1, 2, and 3 (a and b) were assessed on land cover maps derived from high resolution satellite images (Robiglio 2008) using Arc GIS 9.2 (ESRI 1999).
To assess if there was any influence of past land cover and landscape structure we included two historic variables describing past habitat conditions (corresponding to landscape scale variables 2 and 3b) that were assessed on land cover maps for 1984 and 1951 (Robiglio 2008).
Association between variables was explored using two ways contingency tables and Chi-squared test were calculated. Spearman Rank correlation Coefficient was used to assess the correlation between numeric variables. C. Odorata, remnant trees density and occurrence of surrounding vegetation were subjected to ANOVA to test for the association with land cover.
Species Diversity
To compare species richness we used individual based species accumulation curves using rarefaction with 999 iterations (Gotelli and Colwell 2001, Kindt and Coe 2005), whereas for overall species diversity properties we used Rényi’s diversity profile plots in Biodiversity R (Kindt and Coe 2005).
Factors Affecting the Pattern of Abundance of Woody Species
Relationships between species distribution in fallow communities and the three explanatory sets of factors (plot, present landscape and historical landscape) were explored using redundancy analysis (RDA; Legendre and Legendre 1998). We first tested the hypotheses of an influence of each of the three sets of variables. For each set we sought to assess the variation in the species matrix explained by the overall combination of the variables in the set and then identify the variables within the set that were most important in structuring the ordination. Therefore RDA ordinations were carried out for each age class in two steps. In a preliminary RDA we related species data to the explanatory sets. We then performed partial RDA, imposing the strongest variables in the previous step as co-variables in order to differentiate more clearly any species associations to weaker explanatory variables in each set (Legendre and Legendre 1998, Oksanen and others 2007).
We applied the Montecarlo permutation test and rejected the null hypothesis if the overall explanatory set was not significant. The significance of single axes was considered in the general interpretation of species-explanatory set relationships. To assess how strong the relationship was between the species matrix and variable sets we calculated the species-environment correlation for each canonical axis (Legendre and Legendre 1998).
To assess the significance of individual constraints we performed an ANOVA-like test (anova.cca test; Oksanen and others 2007) and assessed the effects of the correlation amongst the variables of each set, comparing the importance and significance of conditional (tested in a joint context) and marginal (tested separately) effects of the variables.
To increase the reliability of each explanatory set, we selected a subset of variables based on their bi-plot scores (>0.3 corresponding to 9% of axis variance; Tabachnick and Fidell 2007) in the rda.output file and the outputs of the anova.cca test. We retained the variables presenting significant conditional and marginal effects (Legendre and Legendre 1998, Oksanen and others 2007).
To understand which factors explained the most of the observed variation we combined them and performed a variance partitioning procedure according to the model presented in Fig. 2. (Legendre and Legendre 1998).

Partition of the variation of woody species responses in fallow vegetation hypothesized in this study. The length of the horizontal line corresponds to 100% of the variation in the response matrix. Adapted from Legendre and Legendre (1998)
We considered only the axes accounting for more than 5% of the overall variability in the first series of RDA and identified the environmental variables that were most responsible for structuring the ordination by selecting those with bi-plot scores for constraining variables in the Biodiveristy R rda. ouput file >0.3 corresponding to 9% of axis variance (Tabachnick and Fidell 2007).
The proportion of variance explained by individual sets of variables were estimated by reiterating conditioned RDA with plot level and landscape level variables used in turn as the constrained and conditioning variables. We then calculated the size of overlap amongst sets of variables by subtracting the sum of plot and landscape scale effects from the amount of variability explained by an ordination model of the combined variables.
A second series of RDA ordinations was performed for the significant sub-sets.
Evidence of association between single species and the explanatory pattern in the RDA were derived from species scores on the canonical axes. Using the equilibrium circle of descriptors (Legendre and Legendre 1998; Kindt and others 2006) we identified the species whose abundance pattern was most clearly associated with the explanatory structure displayed by the bi-plots (Legendre and Legendre 1998).
All the analyses were performed using Biodiversity R (Kindt and Coe 2005) based on functions of the Vegan package of R (Oksanen and others 2007, R Development Core Team 2007).
Results
Floristic Description
The total number of species and families inventoried in young and old fallows in Akok were similar, with 108 species in 37 families and 109 species in 35 families, respectively. In both fallow age classes the most important families in terms of numbers of species were Euphorbiaceae with 16.5 and 13.7% of the species in young and old fallows respectively and Leguminosae-Cesalpinoideae with 7.5% of the species in young and 7.2% in old fallow age classes. In Nkometou, the total species and families inventoried were higher in young than old fallows with 92 species in 34 families and 83 species in 32 families, respectively. As in Akok, Euphorbiaceae was the family with most species, accounting for 18.0% of the species in the young fallows and 14.7% in the older class. In both sites most of the species inventoried were antropophilous of semi-deciduous forest (Letouzey 1985). Besides pioneer tree species like Musanga cecropioides and Macaranga sp., longer-lived, persistent species were found, like Terminalia superba, Ceiba pentandra, Canarium schweinfurthii and Piptadeniastrum africanum (Kahn 1982; Letouzey 1985), as well as species typical of mature secondary forest among which Pycnanthus angolensis, Pentachletra sp. and Petersianthus macrocarpum (Letouzey 1985). As one could expect from the characteristics of the original vegetation described in the map of vegetation by Letouzey (1985) some typical evergreen forest species like Coelocaryon preussi and Caloncoba welwischii, were found in Akok but were absent in Nkometou. There we found, instead, species typical of pioneer forest on savannah-grassland, like Albizia ferruginea, and Allophyllus africanum (Letouzey 1985; Youta 1998).
Species Richness and Diversity
Species accumulation curves were similar at both sites and there was no significant difference amongst curves for young and old fallows at either site (not shown). The curves for the old fallows were not asymptotic indicating that a larger sample would have resulted in a more sound assessment.
In Akok, Rényi diversity profiles for young and old fallows (Fig. 3a), overlapped at the origin reflecting the similar total number of species but for α > 0 old fallow plots were consistently more diverse than the young fallows. In Nkometou the Rényi curves for old and young fallow age classes (Fig. 3b) were very close together but intersected where α was between 0.5 and 1 indicating that diversity of these communities were not comparable.

Comparison of diversity in young (yf) and old (of) fallows in a Akok and b Nkomeotu using Rényi’s diversity profile. Results are based on 999 randomizations
Factors Controlling Woody Species Abundance Patterns
In Nkometou, only the landscape set exhibited a significant effect (P < 0.05) and only in the young fallows, explaining 39% of the variation, with, the 2001 land cover class (type of matrix), bordering mature vegetation and forest distance most important in structuring the ordination. Old fallow samples in Nkometou were excluded from further analysis because overall variation was remarkably low (0.23 compared to 0.67, 0.71 and 0.69 for young fallows in Nkometou and young and old fallows in Akok, respectively), and none of the explanatory sets had a significant effect. This is consistent with the high number of species shared by the fallow samples and the narrow abundance range of those species that were observed in the analysis of stem density and ecological distance between samples presented elsewhere (Robiglio 2008). In Akok, plot and landscape scale sets had significant effects explaining 30.4 and 23.0 % of the variation in young and 47.1 and 51.4 % of variation in old fallows, respectively. In both young and old fallows the plot level ordination was structured in order of importance by C. odorata cover, previous vegetation type (before last cropping), number of fallows and fallow age, with all except the number of previous fallow cycles in old fallows and previous vegetation type in young fallows having significant conditional effects. Tree density also had a bi-plot score of > 0.3 for old fallows but did not have significant marginal and conditional effects. Landscape scale variables important for structuring the ordination differed amongst young and old fallows with, in order of importance, fragmentation pattern, 2001 land cover class, and % of forest (conditional effect not significant) influencing young fallows, whereas, bordering mature vegetation, 2001 land cover class and distance from the forest influencing old fallows.
The set of historical landscape factors had no overall effect on either young or old fallows at either of the sites.
Breakdown of the Total Variability
Results of variation partitioning among the plot and landscape sets are presented in Venn diagrams in Fig. 2. Old fallow samples in Nkometou were excluded from this analysis because of their low variation.
Landscape factors explained more of the variation in woody species abundance than plot factors for old fallows in Akok and young fallows in Nkometou while plot factors explained more variation than landscape factors in young fallows in Akok (Fig. 4).

Break-down of the total variability of plot and landscape sets at Akok (a, b) and young fallows at Nkometou (c). P is the partial effect of the plot variables, L the partial effect of the landscape. The joint Plot-Landscape effect, O was obtained by subtraction. Codes for significance ***P < 0.001, **P < 0.01, *P < 0.05
In Akok, 78% of the species had a at least 30% of their variation in abundance explained by the landscape set and for 53% of the species the landscape sub-set explained more of the variation than the combined plot variables (Fig. 5).

Proportion of variation of the species matrix (x axis) attributable to different sets of variables in the old fallows at Akok. Species codes are given in Appendix 1. Landscape, is the variation explained by the landscape sub-set of variables with the effects of the plot sub-set removed; plot, is the variation explained by the plot sub-set with the effects of the landscape subset removed; overlap is the joint variation that may be equally attributed to the plot or to the landscape sub-set that is obtained by subtraction (Legendre and Legendre 1998). A negative value of the overlap indicates that due to interactions between plot and landscape variables, the two sub-sets combined explain the species matrix better than the sum of the individual effects of plot and landscape (Legendre and Legendre 1998). The sum of the proportions in each bar may be greater than one because categories are not mutually exclusive
Species-Factor Relationships for Significant Landscape Variable Sub-Sets
The landscape sub-set ordination of the woody species matrix in Akok (Fig. 6) was divided between factors characterizing samples in the short fallow matrix (LC01[T.yf]-short fallow) such as scarce mature vegetation around the plot (a_vF_0; a_vF_1), and longer distance from the forest (dist), from factors associated with the long fallow/shrubland matrix (LC01[T.lnf]—long fallow) or forest matrix (LC01F_frg). The most important factor that structured the ordination space was the occurrence of mature vegetation along the fallow plot borders. Musanga cecropioides, Macaranga Barteri and Macaranga spinosa (all forest pioneers) were associated with mature vegetation along about half the plot borders, long fallow matrix and closeness to forest.

RDA ordination bi-plot and equilibrium circle, illustrating the effect of landscape constraints on the woody species abundance matrix for the old fallows in Akok. Species and environment codes are given in Appendices 1 and 2, respectively. Species in red had the largest portion of variation explained by the landscape constraints. Species with vector longer than the circle radius are the species whose variability significantly contributed to the discrimination of canonical axes. RDA axes 2 and 3, RDA2 and RDA3, were significant at P < 0.05 and accounted for 8.3 and 7.7% of the overall variation with species-environment scores of 0.89 and 0.85, respectively. The first canonical axis (RDA1) accounted for 11.2% of the total variation and had a strong species-environment correlation but was not significant (P = 0.06) and is not shown
In contrast, species typical of degraded and exposed conditions such as Alchornea cordifolia and Margaritaria discoidea were abundant in the short fallow/ herbaceous matrix at far distances from forest and with no surrounding mature vegetation. The abundance of forest pioneer species in sites distant from the forest in the short fallow matrix was associated with the presence of surrounding mature vegetation, as in the case of Antochleista schweinfurthii and A. boonei.
In Nkometou (not shown) typical forest pioneer species, M. cecropioides, Bridelia micrantha, and A. schweinfurtii were more abundant in the long fallow matrix in plots with mature vegetation along at least half of their borders. Abundance of M. cecropioides and Trema orientalis were strongly controlled by distance from forest.
Species typical of more degraded habitats like Alchornea cordifolia, Albizia adianthifolia and Harungana madascariensis were abundant in the grassland-herbaceous matrix, far from forest and with little mature vegetation around. In the same type of matrix Voacanga africana and Albizia zygia were abundant where there was some mature vegetation around at least half of the borders.
Discussion
The present study revealed that initial stages of secondary succession in contemporary fallows at different locations in Cameroon present highly different patterns. Such variation is common to other sites the Central African region as the study on two contemporary fallow communities of the Makokou area in Gabon by Mitja and Hladik (1989) confirms.
The effect and significance of the factors controlling the early stages of successions in fallows differed between young and old fallow classes and by site. The preferential association with plot or landscape factors differed among species and for some species differed among sites. Our most important result was that landscape structure and composition may have an independent and statistically significant effect on the pattern of woody species abundance in fallow regrowth and may exceed that for plot characteristics and management.
Historical landscape variables did not show any significant effect on woody species regeneration. The early successional stages considered here were probably inappropriate to assess the long term effects of disturbances and, in fact, reflected the conditions enabling the rapid colonization of the fallow area once cultivation ceases.
In shifting cultivation systems repeated vegetation removal by slash and burn (De Rouw 1995) and weeding during the cropping phase, have a negative, when not dramatic impact on soil seed bank and seedling survival (Norgrove and others 2001) and on resprouting from both stems and roots (De Rouw 1995, Dalle and De Blois 2006). Under such conditions seedlings recruited in early fallow stages derive mostly from newly dispersed seeds that come into the area (Chazdon 2003).
The model we presented for plot level variables reflected the importance of repeated cropping cycles and short fallow rotations: the number of fallow cycles and the type of vegetation before the last cropping were amongst the variables with the stronger effect upon fallow vegetation, after C. Odorata. Forest species were associated with a low number of fallow cycles and/or old fallow or forest vegetation before last clearing, while aggressive and generalist small pioneer trees were more clearly associated with degraded conditions determined by an intense disturbance history. Lawrence (2004), for long fallows in Borneo, found a decline in early succession species richness with increasing number of disturbances and Dalle and De Blois (2006) presented similar results for short fallow areas in Mexico. It is likely that in the short rotational system under study the two variables reflect plot suitability for tree recruitment and growth, for example soil fertility (not considered by this study), presence of microhabitats, and the coverage of C.odorata, than long term species erosion processes.
Chromolaena odorata spread and persistency is favored by intensive farming practices on the plot (de Rouw 1995, Norgrove and others 2001), and in the surrounding areas (Ngobo 2002). Its effects are known: the thick layer reduces tree establishment and growth (Norgrove and others 2001). In our study it affected above all the gregarious light-demanding forest pioneers such as Musanga cecropioides, Trema orientalis and Macaranga barteri. The effect declines with time, once trees become tall enough to prevent new infestation cycles (De Rouw 1995, Norgrove and others 2001), as we observed for the older fallows where the abundance of secondary forest species was strongly associated with an irregular and senescent cover of the weed.
At the landscape scale, agricultural intensification entails the conversion of long fallow agricultural matrices into short fallow matrices. While expanding, the agricultural patches merge, increasing the isolation of the forest fragments (Robiglio 2008). Stronger margin and isolation effects exacerbate degradation processes in the forest remnants (Ewers and Didham 2006), that are also used by local communities to harvest timber and non timber forest products.
The type of agricultural matrix determines the conditions that facilitate or hamper grain dispersal processes and tree establishment, providing (or not) suitable microhabitats and dispersal conditions.
Tests conducted on the association between the variables of plot and landscape sets showed that there are recurrent patterns determined by the way in which fallow shortening and forest conversion processes progress. Young and old fallow matrices were respectively associated (P < 0.01) with: (1) a higher coverage of C. odorata, (2) a higher density of remnant trees. The type of matrix and the amount of mature vegetation surrounding the plot were not significantly associated however, plots belonging to the short fallow matrix had less mature vegetation around than plots of the long fallow matrix.
The implication is that under current trends fallow woody communities turn into distinct communities, dominated by pioneer species typical of degraded conditions that gradually offset pioneer forest species leading to a further decline of the conservation value of shifting cultivation landscapes. The low variation in the old fallow samples in Nkometou already reflected such conditions. In Akok, where both deforestation and intensification process were less pronounced (Robiglio 2008), forest pioneer species were still dominant in both fallow classes. Nonetheless, fallow species richness and diversity were low and non pioneer forest dependent species were rare. Such species are strongly influenced by the proximity to the forest.
Finegan and Nasi (2004) warned about the limited conservation value of fallow communities, under fallow shortening trends. The present study reinforces that view, and highlights the additional risk of repeated short fallow cycles and the expansion of agricultural patches.
The range and complexity of dispersal mechanisms of the species surveyed (Appendix 1) makes any generalization difficult, however we note that in our study the species that were more responsive to the landscape factors were dispersed by vertebrates. Most of the forest species inventoried were dispersed by birds (see Appendix 1). These results support the prediction by Finegan and Nasi (2004) and are crucial with respect to biodiversity conservation. We expect that the composition and extension of the short fallow matrix hamper the presence of dispersal organisms across it.
With the loss of habitat and conduit functions of the agricultural matrix and at an increasing distance from the forest, it is possible that a considerable portion of the matrix no longer experiences forest seed dispersal and may become a potential barrier for re-colonization of land by forest species including pioneers (Nepstad and others 1996, Zimmermann and others 2000). The spatial analysis of landscape change for biodiversity conservation and restoration in agricultural mosaics at the forest margin, should define spatial parameters for the agricultural classes that, based on literature about tree dispersal capacity and seed shower, asses the number and extent of areas beyond a defined distance from the forest edge (analogous to the “core area index” for fragmented forest landscapes; McGarigal and Marks 1995). In terms of seed availability and habitat conditions, fallow secondary regrowth in these internal areas is not subject to any effect of the adjoining forest.
Recommendations for Management at the Landscape Scale
Intensification trends and the turning towards more market integrated systems are part of the inevitable future of smallholder agriculture at the tropical forest margin. Our results suggest some options to intervene in order to maintain some conservation potential at least in identified crucial forest margins (e.g., buffer zones around protected areas or corridors between areas).
From this study it is clear that the landscape is the scale for management designs to conserve/restore biodiversity conservation potential in shifting cultivation landscapes. Taking advantage of existing traditional practices, management schemes should seek to diversify the agricultural matrix, designing the arrangement of the composing patches. We propose three types of complementary designs: (1) improve the fine grained connectivity of short fallow matrices without intervening on fallow length; (2) increase the proportion of old fallow vegetation in the matrix (see Finegan and Nasi 2004); (3) maintain or increase the presence of remnant old growth forests and of forest-like functioning patches such as traditional cocoa agro-forest (avoid deforestation).
With the loss of fallow functions, standing trees, hedges, multi-strata agroforests as well as small degraded forest spots that farmers maintain as a convenient source of forest products (Robiglio 2008), become the foci of conservation and dispersal of forest species across the matrix and improve landscape connectivity. Fine grained interspersion of mature vegetation across the matrix is preserved protecting the mature vegetation spots, included the remnant trees, from fire. In very degraded and poorly interspersed matrices some planting is required. There is very little knowledge about possible techniques to be applied and species to be used, but we know that some of the pioneer long-lived species in secondary forests are of relevant local and international market value (e.g., Terminalia superba, Milicia excelsa, Triplochiton scleroxylon) and are traditionally managed by farmers in the cocoa agroforest. Combining on farm timber production with the landscape scale management of the fallow matrix is an option to explore, in particular for the economic interest that it could have for farmers.
The small effect of remnant tree density on species abundance pattern in the present study suggest that it is important to understand how to ensure/optimize the synergy among factors. Our results lessen the conclusions of studies by Carrière and others (2002), Duncan and Chapman (1999) about the role of scattered trees in facilitating successional dynamics. Once combined in a set the positive effect determined by one single factor may get obscured. In the case of remnant trees further analysis is needed to identify the parameters such as density, species, guild, etc., that may have a more pronounced independent effect and their interactions/association with the other factors.
In the emerging mechanisms for enhancing carbon storage in forest landscapes through reduced emissions from deforestation and degradation, conservation and carbon enhancement (REDD+; Angelsen 2008), there may be new opportunities for communities within shifting cultivation landscapes to obtain value from the second and third designs presented above. However, as the current study shows, beside the carbon enhancement function, where this biomass is retained or increased will be critical for the potential for maintaining and enhancing biodiversity conservation. Mechanisms for valuing mature tree cover more where it matters most for sustaining forest functions across the landscape could have potential to influence individual farmer decisions of which land unit to preserve.
Incorporating a landscape scale perspective where spatial location of interventions matters, is required beyond simply promoting practices that reduce the fraction of cultivated land at the forest margin. This requires a sound understanding of the cultural, institutional and legal framework of current land use systems (local knowledge, farmers resources management strategies, tenure) and of the drivers of change, to explore future agricultural intensification scenarios and assess the feasibility of adapted landscape designs to adopt.
Conclusion
The potential for shifting cultivation landscapes to conserve biodiversity is determined to a considerable extent by the spatial arrangement of individual plot crop/fallow sequences, decided upon by individual farmers. Market forces acting through agricultural intensification, if left unchecked, are likely to continue to exhaust forest functions maintained within shifting cultivation landscapes and seriously jeopardize their potential for biodiversity conservation, checking secondary succession development and favoring the presence of low value species.
There may be opportunities, especially where resources are available for carbon storage, to create incentives for farmers to mitigate degradation trends by retaining more mature vegetation in the landscape. But, if such schemes are to be effective for forest conservation, then valuing retention of mature tree cover where it matters most to sustain woody species regeneration across the landscape, will be required so that farmers’ decisions are influenced in a spatially explicit way.
References
Achard F, Eva HD et al (2002) Determination of deforestation rates of the worlds humid tropical forest. Science 297 no. 5583. Special features on sustainable development pp 999–1002
Ambrose-Oji B (2003) The contribution of NTFPs to the livelihoods of the ‘forest poor’: evidence from the tropical forest zone of south-west Cameroon. International Forestry Review 5:2
Angelsen A (ed) (2008) Moving ahead with REDD: issues, options and implications. CIFOR, Bogor, Indonesia, p 156
ASB, Alternative to Slash and Burn program. http://www.asb.cgiar.org
Brown D (2006) Personal preferences and intensification of land use: their impact on southern Cameroonian slash-and-burn agroforestry systems. Agroforestry Systems 68(1):53–67
Burgers P, Hairiah K, Cairns M (2000) Indigenous fallow management. Lecture Note 4. International Center for Research in Agroforestry, South East Asian Research Programme, Bogor, Indonesia
Carrière SM, Letourmy P, McKey DB (2002) Effects of remnant trees in fallows on diversity and structure of forest regrowth in a slash-and-burn agricultural system in southern Cameroon. Journal of Tropical Ecology 18:375–396
Chazdon RL (2003) Tropical forest recovery: legacies of human impact and natural disturbances. Perspectives in Plant Ecology, evolution and systematics. 6:51–71
Dalle SP, De Blois S (2006) Shorter fallow cycles affect the availability of noncrop plant resources in a shifting cultivation system. Ecology and Society 11(2):2. http://www.ecologyandsociety.org/vol11/iss2/art2/
De Rouw A (1994) Effect of fire on soil, rice, weeds and forest regrowth in a rain forest zone (Cote d’Ivoire). Catena 22:133–152
De Rouw A (1995) The fallow period as a weed break in shifting cultivation (tropical wet forests). Agriculture Ecosystem and Environment 54:31–43
de Wasseige CDD, de Marcken P, Eba’a Atyi R, Nasi R, Mayaux Ph (2009) Les Forêts du Bassin du Congo—Etat des Forêts 2008. Office des publications de l’Union européenne 2009:426
Duncan SR, Chapman CA (1999) Seed dispersal and potential forest succession in abandoned agriculture in tropical Africa. Ecological Applications, Ecological Society of America 9(3):998–1008
Ewers RM, Didham RK (2006) Confounding factors in the detection of species responses to habitat fragmentation. Biological Reviews 81(1):117–142
FAO (2001) Global forest resource assessment 2000. Main report. FAO Forestry Paper No. 140. FAO, Rome, Italy
Ferguson BG, Vandermeer J, Morales H, Griffith MD (2003) Post-Agricultural Succession in El Petén, Guatemala. Conservation Biology 17(3):818–828
Finegan B, Nasi R (2004) The biodiversity and conservation potential of shifting cultivation landscapes. In: Schroth G, da Fonseca GAB, Harvey CA, Gascon C, Vasconcelos HL, Izac AMN (eds) Agroforestry and biodiversity conservations in tropical landscapes. Island Press, Washington, DC, USA
Gockowski J, Tonye J, Baker D, Legg C, Weise S, Ndoumbé M, Tiki-Manga T, Fouaguégué A (2004) Characterization and diagnosis of farming systems in the forest margins benchmark of Southern Cameroon. IITA Social Sciences Working Paper No 1. Ibadan, Nigeria
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters 4:379–391. http://www.blackwell-synergy.com/doi/abs/10.1046/j.1461-248.2001.00230.x
Hardesty BD, Parker T (2002) Community seed rain patterns and a comparison to adult community structure in a West African tropical forest. Plant Eoclogy 164:49–64
Hawthorne WD (1996) Holes and the sums of parts in Ghanaian forest: regeneration, scale and sustainable use. In Swaine MD, Alexander IJ, Watling R (eds) Studies in Guinea-Congo rain forest. Proceedings of the Royal Society, Edinburgh 104b:75–176
Kahn F (1982) La reconstitution de la forêt tropicale apres culture traditionnelle (sud-ouest de la Côte-d’Ivoire). Memoire ORSTOM, Paris, France
Kent M, Coker P (1992) Vegetation description and analysis: a practical approach. CRC Press, Boca Raton; Belhaven Press, London, pp 363
Kindt R, Coe R (2005) Tree diversity analysis. A manual and software for common statistical methods for ecological and biodiversity studies. World Agroforestry Centre (ICRAF), Nairobi
Kindt R, Van Damme P, Simons AJ, Beeckman H (2006) Planning tree species diversification in Kenya based on difference in tree species composition between farms. I. Analysis of tree uses. Agroforestry Systems 67:215–228
Kotto-Samè J, Woomer PL, Moukam A, Zapfack L (1997) Carbon dynamics in slash-and-burn agriculture and land use alternatives of the humid forest zone in Cameroon. Agriculture Ecosystems and Environ. 65(3):245–256
Kupfer JA, Webbeking AL, Franklin SB (2004) Forest fragmentation affects early successional patterns on shifting cultivation fields near Indian Church, Belize. Agriculture Ecosystems and Environment 103:509–518
Kupfer JA, Malanson GP, Franklin SB (2006) Not seeing the ocean fort he islands: the mediating influence of matrix based processes on forest fragmentation effects. Globale Ecology and Biogeography 15:8–20
Lawrence D (2004) Erosion of tree diversity during 200 years of shifting cultivation in Bornean rain forest. Ecological Applications, the Ecological Society of America. 14:1855–1869
Legendre P, Legendre L (1998) Numerical ecology. 2nd English edn. Elsevier, Amsterdam
Letouzey R (1985) Notice de la carte phytogeographique du Cameroun au 1:500.000. Institut de la Carte Internationale de la Végétation, Toulouse, France
Makana JR, Thomas SC (2006) Impacts of selective logging and agricultural clearing on forest structure, Floristic composition and diversity, and timber tree regeneration in the Ituri Forest, Democratic Republic of Congo. Biodiversity and Conservation 15:1375–1397
McCune B, Grace JB (2002) Analysis of ecological communities. MjM Software, Oregon, Gleneden Beach, USA
McGarigal K, Marks BJ (1995) FRAGSTATS: spatial pattern analysis program for quantifying landscape structure. General Technical Report PNW-351, USDA Forest Service
McGrath DG (1987) The role of biomass in shifting cultivation. Human Ecology 15:221–242
Mertens B, Sunderlin W, Ndoye O, Lambin EF (2000) Impact of macro economic change on deforestation in South Cameroon: integration of household survey and remotely-sensed data. World Development 28(6):983–999
Mertz O (2009) Trends in shifting cultivation, and the REDD mechanism. Current Opinion in Environmental Sustainability 1(2):156–160
Mitja D, Hladik A (1989) Aspects de la reconstitution de la végétation dans deux jachères en zone forestière humide (Makokou, Gabon). Acta Oecologica/Oecologia Generalis 10:75–94
Ndam N (1998) Tree regeneration, vegetation dynamics and the maintenance of biodiversity on Mount Cameroon: the relative impact of natural and human disturbance. PhD Thesis, University of Wales, Bangor, UK
Nepstad DC, Uhl C, Pereira CA, Cardoso da Silva JM (1996) A comparative study of tree establishment in abandoned pastures and mature forest of Eastern Amazonia. Oikos 76(1):25–39
Ngobo MP (2002) Ecology and socio-economic importance of short fallows in the humid forest zone of Southern Cameroon. PhD Thesis, University of Wales, Bangor, UK
Ngobo M, McDonald MA, Weise S (2004) Impacts of type of fallow and invasion by Chromolaena odorata on weed communities in crop fields in Cameroon. Ecology and Society 9(2):1
Norgrove L, Hauser S, Weise S (2001) Response of Chromolaena odorata to timber tree densities in an agrisilvicultural system in Cameroon: aboveground biomass, residue decomposition and nutrient release. Agriculture Ecosystems and Environment 81(3):191–207
Nounamo L, Yemefack M (2002) Farming systems in the evergreen forest of southern Cameroon: shifting cultivation and soil degradation. Tropenbos Cameroon Documents 8, Kribi, Cameroon
Oksanen J, Kindt R, Legendre P, O’Hara RB (2007) Vegan: community ecology package Version 1.8–6. http://cran.r-project.org/
Padoch C, Coffey K, Mertz O, Leisz SJ, Fox J, Wadley RL (2005) The Demise of Swidden in Southeast Asia? Local Realities and Regional Ambiguities. Danish Journal of Geography 107(1):29–41
Palm CA, van Nordwijk M, Woomer PL, Alegre JC, Arevalo L, Castilla CE, Cordeiro DG, Hairiah K, Kotto-Samè J, Moukam A, Parton WJ, Ricse A, Rodrigue V, Sitompul SM (2005) Carbon losses and sequestration following land use change in the humid tropics. In: Palm CA, Vosti SA, Sanchez PA, Ericksen PJ, Juo ASR (eds) Slash and burn: the search for alternatives. Columbia University Press, New York, NY, USA
PC-ORD. Multivariate Analysis of Ecological Data. Version 4.34. MjM Software, Gleneden Beach, Oregon, USA
Poorter L, Bongers F, Kouame FN, Hawthorne WD (eds) (2004) Biodiversity of West African forests. An ecological atlas of woody plant species. CABI Publishing, Oxford, UK, p 528
R Development Core Team (2007) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org
RI ES (1999) ArcGIS v9.2. Redlands CA, USA
Robiglio V (2008) Beyond slash and burn: landscape ecology of shifitng cultivation systems in Southern Cameroon. PhD Thesis, University of Wales, Bangor, UK
Swallow B, van Noordwijk M, Dewi S, Murdiyarso D, White D, Gockowski J, Hyman G, Suseno B, Robiglio V, Meadu V, Ekadinata A, Agus F, Hairiah K, Mbile P, Sonwa DJ, Weise S (2007) Opportunities for avoided deforestation with sustainable benefits. An Interim Report by the ASB Partnership for the Tropical Forest Margins. ASB Partnership for the Tropical Forest Margins, Nairobi, Kenya, 42 p
Tabachnick BG, Fidell LS (2007) Using multivariate Statistics. 5th edn, Pearson International edn. Boston, USA
Tchouto MGP (2004) Plant diversity in a Central African rain forest: implications for biodiversity conservation in Cameroon. PhD Thesis, Wageningen University
Toledo VM, Ortiz-Espejel B, Cortés L, Moguel P, Ordoñez MDJ (2003) The multiple use of tropical forests by indigenous peoples in Mexico: a case of adaptive management. Conservation Ecology 7(3):9. http://www.consecol.org/vol7/iss3/art9/
TREES (1995) Study of forest/non-forest interface. Typology of fragmentation of tropical forest. CEC/ISPRA Joint Research Centre Trees Series B: Research Report No. 2. Joint Research Center, European Space Agency, Luxemburg
TREES (1998) Identification of deforestation hot spot areas in the humid tropics. CEC/ISPRA Joint Research Centre Trees Series B: Research Report No. 4. Joint Research Center, European Space Agency, Luxemburg
Uhl C (1987) Factors controlling succession following slash-and-burn agriculture in Amazonia. The Journal of Ecology 75(2):377–407
van Noordwijk M (2002) Scaling trade-offs between crop productivity, carbon stocks and biodiversity in shifting cultivation landscape mosaics: the FALLOW model. Ecological Modelling 149:113–126
Vivien J, Faure JJ (1989) Arbres des Forets Denses d'Afrique Centrale. Especes du Cameroun. Republique francaise; Ministere des Relations Exterieures, de la Cooperation et du Developpement. Agence de Cooperation Culturelle et Technique
Youta H (1998) Arbres contre graminee: la lente invasion de la savane par la foret au Centre-Cameroun. Thesis. Universite de Paris-Sorbonne, Pparis IV, u.f.r. de geographie et amenagement. Paris, France
Zimmermann JK, Pascarella JB, Aide TM (2000) Barriers to forest regeneration in and abandoned pasture in Puerto Rico. Restoration Ecology 8(4):350–360
Acknowledgments
The authors acknowledge ASB, the Alternative to Slash and Burn program at the World Agroforestry Centre (ICRAF) Nairobi for assisting in the accomplishment of this research project. We also acknowledge funding from the Italian Foreign Ministry that covered the costs of work in Cameroon at the International Institute of Tropical Agriculture (IITA) Regional Station in Yaoundé. The study was part of a PhD degree within the School of the Environment and Natural Resources, Bangor University, UK. Thanks to the two anonymous reviewers and the editors for their comments and guiding.
Author information
Authors and Affiliations
Corresponding author
Appendices
Rights and permissions
About this article
Cite this article
Robiglio, V., Sinclair, F. Maintaining the Conservation Value of Shifting Cultivation Landscapes Requires Spatially Explicit Interventions. Environmental Management 48, 289–306 (2011). https://doi.org/10.1007/s00267-010-9611-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-010-9611-2