
Abstract
Public rangelands in North America are typically managed under a multiple use policy that includes livestock grazing and wildlife management. In this article we report on the landscape level extent of grassland loss to shrub encroachment in a portion of the Rocky Mountain Forest Reserve in southwestern Alberta, Canada, and review the associated implications for simultaneously supporting livestock and wildlife populations while maintaining range health on this diminishing vegetation type. Digitized aerial photographs of 12 km of valley bottom from 1958 and 1974 were co-registered to ortho-rectified digital imagery taken in 1998, and an un-supervised classification used to determine areas associated with grassland and shrubland in each year. Field data from 2002 were over-layed using GPS coordinates to refine the classification using a calibration-validation procedure. Over the 40-year study period, open grasslands declined from 1,111 ha in 1958 to 465 ha in 1998, representing a 58% decrease. Using mean production data for grass and shrub dominated areas we then quantified aggregate changes in grazing capacity of both primary (grassland) and secondary (shrubland) habitats for livestock and wildlife. Total declines in grazing capacity from 1958 to 1998 totaled 2,744 Animal Unit Months (AUMs) of forage (−39%), including a 58% decrease in primary (i.e., open grassland) range, which was only partly offset by the availability of 1,357 AUMs within less productive and less accessible shrubland habitats. Our results indicate shrub encroachment has been extensive and significantly reduced forage availability to domestic livestock and wildlife, and will increase the difficulty of conserving remaining grasslands. Although current grazing capacities remain marginally above those specified by regulated grazing policies, it is clear that continued habitat change and decreases in forage availability are likely to threaten the condition of remaining grasslands. Unless shrub encroachment is arrested or grassland restoration initiated, reductions in aggregate ungulate numbers may be necessary.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Considerable interest has focused on changes within grasslands of North America, including the threat of encroachment by woody species (Archer and others 1988; Schlesinger and others 1990; Briggs and others 2005) leading to major landscape-scale changes in natural vegetation (Baker 1992; Turner and Romme 1994). Many studies have documented rates and patterns of shrub and forest expansion into prairie ecosystems (Bailey and Wroe 1974; Bragg and Hulbert 1976; Mast and others 1997; Briggs and others 2002a; Bai and others 2004). Causes of this expansion include fire suppression, leading to selection for tall-statured species that accumulate aboveground biomass (Knight and others 1994; Cook and others 1994; Briggs and others 2005), changes in climate that render shrubs more competitive than understory vegetation relative to the amount and timing of available moisture (MacDonald 1989; Sturm and others 2001), nitrogen deposition (Köchy and Wilson 2001) and grazing (Dunwiddie 1977; Briggs and others 2002b). Preferential selection of herbs during defoliation can facilitate woody plant invasion by initiating an interspecific competitive shift within the community toward the shrub component. Although the phenomenon of shrub encroachment has been well documented, the implications of grassland conversion to woody species are not well known (Wessman and others 2004).
Within the Rocky Mountain Forest Reserve (RMFR) of west central Alberta, floodplain meadow grasslands are small in area but are important sources of wildlife habitat, native biodiversity, and summer range for livestock (Willoughby 2001). Similar to the western US (Cook and others 1994), fire suppression in this portion of the Rocky Mountains appears to have favored the establishment of mountain-shrub communities, particularly those dominated by bog birch (Betula glandulosa Michx.).Footnote 1 Although this species is known to tolerate and even increase with occasional burning (Bork and others 1996; DeGroot and Wein 1999), presumably due to its long evolutionary exposure to periodic fire, more frequent wildfires likely played an historical role in keeping the abundance of this species in check (DeGroot and others 1997). Native Americans historically burned valley bottoms to establish camps and improve habitat to attract game (Lewis 1980), with frequent, low intensity burns likely maintaining valley bottoms as herbaceous meadows (Willoughby 2001). Long-term rangeland reference areas in the RMFR indicate shrub abundance has increased 15% regardless of livestock presence (Willoughby 2000). Additionally, shrub cover inside enclosures protected from cattle is greater than outside (Willoughby 2000), suggesting cattle grazing has played a role in impeding shrub encroachment.
Where they remain, the maintenance of native grasslands is important not only from the perspective of conserving biodiversity (Watkinson and Ormed 2001) but also as a source of forage for simultaneously supporting cattle ranching in summer and resident elk herds on this public land-base (Adams and others 1992; Bork and others 1996; Willoughby 2001). As the encroachment of woody species into grasslands is widely known to reduce forage availability (e.g., Bailey and Wroe 1974; Briggs and others 2005), grassland loss may increase land use conflict, including the risk of reducing the health associated with remaining grasslands through overgrazing. This threat, in turn, may necessitate either a reduction in livestock grazing or the increased harvest of wildlife. Consequently, public land managers are increasingly under pressure to maintain native grasslands as well as restore those areas lost to shrub encroachment. Prescribed burning has been used sporadically in an attempt to control shrubs in the region (Bork and others 1996) but remains difficult to undertake for several reasons, including the high cost of burning, the brief burn window in these mesic environments, and the high potential impacts of escaped fires on adjacent economic activities (forestry and energy extraction).
Effective grassland restoration within floodplain meadows depends on accurate knowledge about the historical abundance of this type of habitat and any vegetation changes since fire suppression began. Reliable information on the landscape area affected by shrub encroachment can then be used by managers to modify existing management plans where land use conflicts (e.g., over the allocation of forage) may exist. Moreover, this information can establish the urgency of grassland restoration and identify target abundances of habitats such as grassland and shrubland, which can then be met through specific actions such as prescribed burning or other restoration procedures.
The detection of shrub encroachment and quantification of landscape changes from grassland to shrubland is crucial for ensuring progress toward long-term land management objectives. This process can be accomplished using remotely sensed data and GIS applications (Tueller 1989; Mast and others 1997; Bai and others 2004). The widespread availability of repeat aerial photography for much of the RMFR, dating back to as early as the 1950s, provides the unique opportunity to assess landscape-level changes in habitat over time.
As the implications of shrub encroachment for land management are often poorly understood (Wessman and others 2004), this study was developed to further our understanding of the linkage between encroachment, habitat change and potential land use conflict. Previous efforts to integrate spatial information on vegetation change with production data to address forage allocation conflicts and threats to range health are uncommon. The specific objectives of this study were to (1) quantify the spatial extent of grassland loss (and therefore, shrub encroachment) over a 40-year period within grassland meadows in a portion of the Clearwater River drainage in the Upper Foothills of the RMFR, (2) document changes in grazing capacity associated with observed vegetation changes, and (3) examine the impact of encroachment on potential land use conflicts between livestock grazers and wildlife managers, as well as the risk to range health.
Methods
Study Area
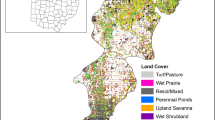
We evaluated floodplain meadows in a portion of the Clearwater Grazing Allotment 80 km southwest of Rocky Mountain House, Alberta, Canada (52° 2′ 37′′ N; 115° 32′ 56′′ W). We focused on 12 km of valley bottom encompassing 2,392 ha along Elk Creek and Radiant Creek (Fig. 1), close to where they drain into the Clearwater River, at 1,425–1,530 m elevation. Grasslands in the region are on outwash plains, eskers, or glaciofluvial terraces, on areas with 1–9% slope (Rayner 1984). The study area receives an average of 538 mm of precipitation annually, with over half (340 mm) in the summer. Temperatures average 11.5°C in summer and −6.0°C in the winter, with only 75 frost free days (Hanson 1973).

Overview of the Clearwater Grazing Allotment (top) and the five sub-regions within which shrub encroachment was assessed between 1958 and 1998
The study area is located within the Upper Foothills Natural Subregion (Strong and Leggat 1992), with widespread abundance of lodgepole pine (Pinus contorta Loudon) on adjacent slopes (Cormack 1953). Valley bottoms contain a mosaic of shrubland and mesic grassland. Dominant grasses are foothills rough fescue (Festuca campestris Rydb.) and tufted hairgrass [Deschampsia cespitosa (L.) Beauv.], with the predominant shrubs bog birch (Betula glandulosa Michx.) and Barclay’s willow (Salix barclayi Anderss). Valley bottoms within the study area, including the associated shrubland communities, are representative of those found in the broader Clearwater River drainage and much of the RMFR. Soils are largely Regosols or Gleysols on medium to course textured alluvial deposits, with small pockets of medium to fine textured alluvium and mesic Organics (Rayner 1984).
Land Use History
The study area is public land managed by the Rangeland Management Branch (RMB) within the department of Alberta Sustainable Resource Development (ASRD), and is managed for multiple uses under the Forest Reserves Act (Government of Alberta 2000), including the maintenance of biodiversity. While forest harvesting and energy extraction are common on uplands, valley bottoms are used extensively by livestock and as wildlife habitat by ungulates, particularly elk. Cattle grazing started in the area during the 1930s, with little regulation of use during the first 23 years. In 1953, the area became part of the Clearwater Grazing Allotment, with the first management plan in 1958 establishing a livestock stocking rate of 3,080 Animal Unit Months (AUMs), where one AUM is the amount of forage necessary to support a single 454 kg cow with or without a calf up to 6 months of age for 1 month duration (Society for Range Management 1989). This livestock stocking rate was thought to represent 50% of the maximum carrying capacity and accommodated a large elk population that used the valley bottoms during winter and spring calving (Weerstra 1990). A second management plan in 1972 had a revised livestock stocking rate of 2,250 AUMs, and in 1976, a rotational grazing system was implemented with grazing throughout the summer and early fall (Weerstra 1990). As part of the provincial policy for resource management within the eastern slopes of the Rocky Mountains, allowable livestock stocking rates for the Clearwater Allotment were to be maintained at the 1977 level of 2,803 AUMs. The current study examines a portion of the Clearwater Allotment, and includes Elk Creek and Radiant Creek, with the most recent management plan (unpublished ASRD report) identifying a specific livestock grazing capacity for this area of 1,102 AUMs.
Scientific Approach
A GIS framework was used to conduct all spatial analyses. Digital ortho-rectified photography of the study area from 1998 was initially converted to a grid, and a natural breaks classification used to find classes in the data. The initial classification was then refined using a calibration–validation procedure employing field transects to more accurately identify the range of pixel values associated with grassland and shrubland. The same procedure for establishing habitat classes was repeated using digitized imagery from 1958 and 1974, although validation of the latter data sets was not possible due to the lack of field data for those dates other than photographs and limited data collected from a single grazing enclosure (i.e., Elk Creek enclosure). Finally, calculated areas of grassland in 1958 and 1974 were compared against the area of grassland remaining in 1998.
Image Preparation
Historical aerial photos were obtained of the study area west of Rocky Mountain House and selected based on their quality and scale. The 1958, 1974 and 1998 photography were taken 15 August at a scale of 1:15840, 1 September at a scale of 1:21120, and 15 September at a scale of 1:40000, respectively. The 1998 photography was rectified using provincial base map features in digital format. Scanning of 1958 photos led to a ground resolution of 0.16 m2, which contrasted with a resolution of 0.56 m2 for 1974 and 1998. The two earlier air photos were co-registered to the 1998 ortho-photography in Arcinfo using a minimum of 30 ground control points concentrated in valley bottoms, including roads, trails and creek crossings. The RMSE of registering the 1958 and 1974 data to the 1998 data were 11 and 16 m, respectively.
Digital images were initially clipped to contain all meadows in the study area; geographic coordinates were retained in the process. Images were further divided into five sub-regions of valley bottom to facilitate analysis (Elk, South Elk, Radiant, West Radiant, and Elk-Radiant Transition), which ranged from 132 to 797 ha in size. The same shape file was used to clip images of the study area from each time period, ensuring analysis of the same spatial extent in all 3 years.
The geographic coordinates of 33, 30-m linear field transects were overlayed on the digital images, and used to refine the initial classification. Transects were used to collect vegetation data on shrub and understory forage abundance. Coordinates and elevation (m) were obtained using a GPS (Global Positioning System). Transects overlayed on the digital imagery were spatially corrected for each year.
Field Sampling
Transects (n = 33) used in the calibration–validation exercise were sampled in July of 2002. We assumed that portions of the study area dominated by grassland in 2002 were grassland in 1998, a reasonable assumption given the time delay of only 4 years and lack of shrub clearing (and fire) in the interim. Transects were established in pairs throughout the study area, with one in grassland and another in adjacent encroached shrubland (i.e., areas relatively free of shrubs in 1958). Pairs were maintained on the same ecological site (Society for Range Management 1989) to minimize variability in growing conditions (i.e., moisture, nutrient, drainage, and soils). Transect distribution was throughout the study area and designed to capture a broad range of variation in shrub abundance (i.e., open through closed communities), although it later became evident that few communities had shrub cover between 20 and 50%. We attributed this to rapid rates of shrub canopy closure following the initial establishment of shrubs near the leading edge of encroachment, which in turn, appeared to limit opportunities to sample stands with intermediate amounts of shrub cover.
Measurements of vegetation on each transect included the canopy cover of shrubs using the line intercept method (Bonham 1989), in which all linear portions of each transect overtopped by shrubs was recorded, and aboveground understory herbaceous production. The latter was harvested to ground level within 2, 0.5 m2 quadrats randomly placed along each transect in July of 2002, with samples dried to constant mass and weighed. Minimum sample areas needed to obtain reliable estimates for each transect were determined prior to sampling (Burkinshaw 2005). All biomass quadrats were protected from large animal herbivory during the year of sampling using portable range cages.
Imagery Analysis
Prior to classification, all digital images were converted to grids to facilitate processing, after which an initial unsupervised classification was done using the natural breaks method. This procedure established breakpoints between classes using a statistical formula (Jenk’s optimization) that minimizes the sum of the variance within each class (ESRI 2002). During classification the number of classes was set to five, which was considered to comprise all key habitats. These classes included water as the darkest pixels with low digital values, roads and trails with bright pixels and high digital values, and three vegetation classes delineating conifer forest, shrubland and grassland.
While this procedure was effective in initially separating the various cover classes, we used a calibration–validation exercise to refine the range of pixel values associated with the two key cover classes of interest, shrubland and grassland. This process determined the digital values of all pixels in the 1998 imagery along seven randomly selected transects (training sites), including three in grassland and four in shrub dominated areas. A sensitivity analysis of the digital pixel values was conducted to evaluate variation in the sampling “width” of field transects. This was done because although pixel rows were 0.75 m wide, similar to the width of field transects, the ability of the spatial analysis to accurately correlate field transects with the digital data may have been limited by the error of GPS determination of transects, as well as the registration error of digital data. Three randomly selected transects were chosen and digital value ranges examined across consecutive increases in pixel sampling “band” width, including one row of pixels (n = 41 pixels: 0.75 m × 30.75 m in length), and increments of one additional row, up to 6 rows in total (n = 246 pixels: 4.5 m by 30.75 m). While increasing the width of digital sampling from one to six rows led to an increase in the range of digital values for grass and shrub cover classes, mean pixel values for each transect remained stable regardless of the number of pixel rows examined, indicating a narrow row width of only 1 pixel was adequate in all subsequent analysis.
During calibration with the field data, pixel values associated with shrub cover were obvious along most transects. Although portions of transects dominated by grassland were highly visible before and after the reclassification process during calibration, distinguishing whether individual pixel values were indicative of the presence of grassland in transects containing between 12 and 60% shrub cover was more difficult. Fortunately, all but 5 of the field transects examined consisted of shrub cover over 60%, making identification of the majority relatively easy. Conversely, intermediate communities containing moderate amounts of shrub were the most difficult to assess, but are arguably the most important as they represent those areas undergoing active succession from open grassland to shrubland.
Calibration ultimately involved determining the number of pixels (out of 41) in each of the seven individual training transects occupied by shrubland or grassland. Once digital pixel value ranges associated with each type of habitat were delineated within the training transects, the remainder of the digital grid for the study area was reclassified using the established digital cutoff between classes. Accuracy of the initial calibration was assessed (i.e., validated) by testing the empirical relationship (R 2) between predicted (i.e., from digital values) and actual shrub cover data from 2002 within the remaining transects (n = 26) not used in the calibration.
Finally, the same procedure for classifying grassland and shrubland was applied to the 1958 and 1974 imagery. As images from different years varied in baseline intensity, pixel value cutoffs varied: for example, grassland was represented by digital values greater than 121, 156 and 165, in 1958, 1974 and 1998, respectively. Although it was not possible to validate earlier classifications, information from the Elk Creek enclosure was used to supplement the classification of earlier imagery. This enclosure was established in 1965 by ASRD to monitor the long-term trend in range health under livestock grazing (Weerstra 1990). We also assumed areas free of shrubland in 2002 around the enclosure were free of shrubland in 1958 and 1974, a conclusion supported by historical field data and ground photos (Weerstra 1990). As a result, the area immediately around the enclosure was used to help determine pixel values associated with grassland for the earlier sampling times.
Assessing Grassland Change
To quantify changes in grassland between 1958 and 1998, the area classified as grassland for each sub-region was compared between time periods. Total areas of grassland were calculated by multiplying pixel sizes from a given imagery date by the number of pixels identified as grassland. Given that this procedure concentrated on classifying grassland, and all of the study area was either grassland or shrubland following the removal of other vegetation types in the initial classification, only the area of grassland is presented here.
Changes in grassland were also corrected using the empirical relationship between predicted and actual shrub abundance from 2002 (Fig. 2): predicted shrub cover over-estimated actual cover by an average of 15.4%. As a result, predicted grassland areas were adjusted by multiplying the area of calculated grassland in each sub-region by a coefficient of 1.154. Finally, adjusted grassland areas from 1958 to 1998 were combined with mean forage production from each type of habitat as identified by Bork and Burkinshaw (2009) to quantify changes in primary (i.e., grassland) and secondary (i.e., shrubland) grazing capacity, standardized to AUMs. Grazing capacity was calculated by multiplying the area of affected grassland or shrubland by the corresponding production levels, and included a standard 50% adjustment for allowable utilization on public land to maintain range health. Grasslands (<12% shrub cover; n = 16 transects) produced a mean of 4,396 kg ha−1 herbage production, while shrublands (>34% shrub cover; n = 17 transects) produced 1,454 kg ha−1.

Relationship between actual and predicted shrub canopy cover on transects used for validation (SE = 3.5%, RMSE = 0.12, P < 0.0001; n = 26). Average difference between predicted and actual shrub cover = 15%. The dotted line represents the 1:1 slope
Results
Grassland Abundance
Vegetation field data from 2002 revealed a minor shrub abundance associated with a number of transects that were predominantly grassland. As a result, a threshold shrub cover level was used to discern shrubland from grassland habitat. Sixteen transects had under 12% shrub (willow and bog birch) cover, and 17 transects had between 34% and 92% cover, indicating a distinct split between grassland (<12%) and shrubland (>34%) habitats. Moreover, declines in production with progressive increases in shrub cover were non-linear, with more pronounced decreases within communities containing shrub canopies that were less than 30% (Bork and Burkinshaw 2009).
Results of the calibration–validation exercise using field data from 2002 demonstrated the accuracy of determining those pixels associated with grassland and shrubland in the ortho-photography of 1998. The standard error of predicted shrub cover across validation transects was 3.5% with a mean difference between actual and predicted shrub cover of 15.4% (Fig. 2). Although the relationship between actual and predicted shrub abundance had a high Goodness-of-Fit (r 2 = 0.89), the error arose from a tendency to systematically over-estimate shrub cover (%) for all transects. Additionally, absolute predicted shrub cover was more accurate when fewer shrubs were on the site. This finding may be attributed to the range of pixel values for shrubland being set too low in the calibration exercise, or more likely, from pixel values associated with the shadows of shrubs being incorporated into the shrubland class, resulting in their over-estimation. As a result, adjusted grassland areas were emphasized in all reporting to provide a more conservative estimate of grassland loss.
Temporal Changes in Grassland Area and Carrying Capacity
There was minor variation among all sub-regions of the study area (Elk, South Elk, Radiant, West Radiant, and Elk-Radiant Transition) in the extent of decrease in grassland throughout the 40-year study period (Table 1; Fig. 3). Data from Elk Creek and West Radiant demonstrated the greatest decrease in grassland, but also had the most area of grassland in 1958 at the start of the monitoring period. Relative losses in grassland among sub-regions varied from a low of 53% at Radiant to a high of 67% at South Elk (Table 1). Across the study area, grassland loss was 413 ha (−37.2%) between 1958 and 1974, and another 233 ha (−21%) between 1974 and 1998. The total area of grassland remaining in 1998 was 647 ha, indicating 58% of grasslands had been lost since 1958 (Table 1).

Comparison of a portion of the Radiant Creek subregion as photographed in 1958 (top) and 1998 (bottom). Light areas represent grassland, gray areas shrubland, and the darkest areas conifer forest. Points represent the start and end point of transects sampled in 1998, and the same areas in 1958. Note the marked difference in shrubland between photos
While the shift from grassland to shrubland across the study area was marked, this change may not have captured the entire loss of grassland over the time period examined. Observed vegetation changes may also be attributed in part to conifer encroachment, though it is unknown whether conifers encroached into shrub or grassland habitat. Although not analyzed in our study, visual comparisons of the 1958 and 1998 photos indicated there appeared to be a small increase in conifer trees that we estimated to be less than 1% of these floodplain meadows (e.g., see Fig. 3).
Grazing capacity changed sharply from 1958 to 1998 due to a shift toward less productive and less accessible forage in a landscape with increasing shrubland (Table 2). In 1958, the estimated grazing capacity of open grasslands, known as primary range, across the study area was approximately 7,047 Animal Unit Months (AUMs). In 1974, total grazing capacity dropped to 5,293 AUMs, of which only 4,426 AUMs was primary range, with the balance contributed by forage in areas dominated by shrubs at that time. From 1974 to 1998, total grassland grazing capacity decreased to 4,303 AUMs, of which only 2,946 AUMs was primary grasslands (Table 2). Over the complete 40-year time period the decrease in primary grazing capacity was 4,101 AUMs. Even when contributions from secondary ranges were included in the assessment of grazing capacity, the decline in AUMs remained 39%.
Discussion
Changes in Grassland
A key objective of this study was to assess landscape level changes in vegetation, namely grassland and shrubland, during a 40 year period. By examining a time series of aerial photos, we successfully quantified rates of grassland loss within this portion of Alberta’s RMFR, which remained relatively consistent across sub-regions. Moreover, the vegetation changes we documented remained apparent even after adjustments were made for potential misclassification error between vegetation types (i.e., error associated with classifying grassland pixels as shrubland). The magnitude of the 58% decrease in grassland area from aerial photos between 1958 and 1998 clearly highlight the role of shrub encroachment (Table 1, Fig. 3). While we acknowledge that our study examined only a small portion of the RMFR (i.e., 2,492 ha of river floodplain), the area examined does constitute a large fraction (38%) of the primary and secondary range available for grazing within the 53,295 ha Clearwater Grazing allotment. Additionally, our findings are supported by localized data from other regions of the RMFR (e.g., Adams and others 1992; Willoughby 2000) reinforcing that grassland loss is a widespread phenomenon in the region. Finally, although we were not able to identify the specific mechanisms responsible for facilitating shrub encroachment, which include climate change, grazing and fire suppression, the current study is unique in that it provides quantitative information on the spatial extent of grassland loss at the landscape level.
The majority of shrubs within this ecosystem (~70%) were bog birch, a species that that can exhibit high seed production (Weis and Hermanutz 1988), which in turn, may account for much of the observed increase in shrub over time. In addition, both bog birch and the associated willows within these habitats reproduce extensively through vegetative layering, allowing species like bog birch to spread up to 5 m from the parent plant (DeGroot and others 1997) even when conditions for seedling recruitment are not favorable. Bog birch distribution and abundance are known to be temporally dynamic in other regions (Edwards and others 2008), with this species strongly influenced by climatic variation (DeGroot and others 1997). Warmer conditions have been shown to lead to increases in this species (DeGroot and Wein 1999). Thus, climate change may be a key factor positively influencing shrub encroachment in this ecosystem, similar to other shrublands in North America (Archer and others 1995).
Together with shrubs, there was a small visible increase in conifers within the air photos examined (Fig. 3), and small pine and spruce trees were noted in 4 of the 33 field transects sampled. This is not surprising given that shrubs can facilitate forest invasion along ecotonal boundaries (Petranka and McPherson 1979). Increases in the density, cover and height of tall shrubs such as willow and bog birch also coincided with the disappearance of key native bunchgrasses from these communities (Bork and Burkinshaw 2009), highlighting the potential degree to which the range health of these communities may be affected. Collectively, these observations support the previously publicized notion of widespread successional advancement across the study area (Willoughby 2001), and indicate that should these advancements continue, remaining grasslands may eventually be restricted to those ecological sites that are too dry or nutrient poor for woody species establishment, the extent of which remains unknown. Although annual rates of recent grassland loss between 1974 and 1998 (9.7 ha year−1) are lower than those observed during the first portion of the study period from 1958 to 1974 (25.8 ha year−1), there is little reason to believe that shrub abundance had peaked within the study area in 1998.
The decline in primary range grazing capacity of 4,101 AUMs, including overall grazing capacity of 2,744 AUMs, across the study area reflects the serious impact that grassland loss has on the availability of forage within alluvial floodplains. In recent management plans developed by ASRD, managers have made a commitment to maintain the 1977 stocking level of 1,102 AUMs for the study area examined here. According to our current estimates, the Elk and Radiant Creek current primary grazing capacity (2,946 AUM) remains well above that mandated under the current provincial grazing guidelines. Despite the apparent abundance of forage supply, however, it should be noted that the government approved grazing capacity does not account for forage use by elk in the region. While limited in accuracy, the most recent estimates of elk populations from 2005 in the immediate region indicate there are at least 59 elk in the Elk-Radiant drainage. Factoring in the 496 AUMs that elk require for their year-round forage needs, this brings the total current forage demand to 1,598 AUMs. Although this number remains 2,706 AUMs below the area’s total grazing capacity based on primary and secondary range (4,304), public land managers need to be vigilant of the magnitude of the extent of grazing capacity changes during the preceding 40-year period, and be aware that further declines in forage may result in continued convergence of actual and mandated grazing capacities.
Of the 58% decrease in primary grazing capacity, 37% occurred prior to 1974, and another 21% after 1974, suggesting further reductions in grazing capacity are probable with continued shrub encroachment. Moreover, because cattle rarely extend use into secondary (i.e. shrub dominated) ranges, the potential for degradation of remaining grassland habitats is high, particularly with these bunchgrass communities being sensitive to overuse (Willms and others 1985). Additionally, it must be noted that grazing can accelerate shrub encroachment (Brown and Archer 1999; Van Auken 2000; Briggs and others 2002b), a process linked to interspecific competitive shifts in favor of shrub rather than grassland plant species. Although elk will utilize browse in their diets, particularly during winter, this is less likely in the case of bog birch due to the chemical defenses of this plant species (Bryant and others 1989; DeGroot and others 1997) leading to its relatively poor forage value. Accordingly, elk may be reluctant to alter their foraging patterns to include shrublands until after significant use has occurred of open grasslands, particularly as elk preferences during foraging have been linked to habitats with maximum grass biomass (Hebblewhite and others 2008). More intensive use of a declining grassland area, in turn, is more likely to compromise other land use values such as the maintenance of range health and associated biodiversity. This is particularly likely given that adjacent forests, while widespread in area, are coniferous and offer very little available forage in the understory (Willoughby 2001) and have also been associated with increased predation risk on elk from wolves (Hebblewhite and others 2005).
Worldwide, grasslands are considered one of the most threatened natural regions on earth (Wessman and others 2004). Although this particular study area is not in the grassland natural region of Alberta, it does have an important grassland component (Bork and others 1996; Willoughby 2001) that contributes significantly to local and regional biodiversity, as well as wildlife habitat within the province (e.g., Pengelly and Hamer 2006). Loss or degradation of unique grasslands is widely recognized as a significant threat to biodiversity and their conservation is an important objective in natural resource management (West 1993).
Current management in the RMFR is striving to develop and adopt a strategy for conserving Alberta’s biodiversity, which in turn, will set the course for minimizing the loss of biodiversity and associated economic, environmental, social and political risks. Unlike private land, where a single user may restrict land use in the interest of achieving resource conservation, addressing biodiversity issues on public land requires the effective integration of a wide variety of land uses and associated stakeholder interests, and thus reflects a shared vision consistent with maintaining multiple uses. In the face of ongoing shrub encroachment within the RMFR, it appears public land managers will have to contribute to the effort of conserving remaining grasslands by making difficult decisions in the future, including working to control further encroachment by shrubs, as well as restoring those grasslands already invaded.
Potential methods to control shrubs include the use of mowing, herbicides or prescribed burning. While herbicides are an unattractive option in areas where the retention of plant diversity is important, and mowing is likely to be difficult and expensive to undertake, prescribed burning of valley bottoms in the RMFR is perhaps the most feasible way to reduce shrubs and restore grassland (Bork and others 1996). The historical use of fire for the control of bog birch has been documented elsewhere (Edwards and others 2008). In addition to damaging shrubs, burning can promote seed set of key forage species and initiate a flush of early successional forbs (Bork and others 2002). Despite the known historical importance of fire in the region, prescribed burning is not widely implemented at this time, presumably due to the risk associated with its use. Moreover, repeated fires are necessary on a frequent interval to achieve control of bog birch, which is well-adapted to and highly tolerant of fire (Bork and others 1996; DeGroot and Wein 2004).
Perhaps most important, once woody plants dominate these grasslands as in the current situation the fire regime is likely to change. For example, the low intensity, frequent fires created from abundant fine fuels that would have historically kept these valley bottoms dominated by grasslands, many of which were introduced by native peoples for vegetation management (Lewis 1980), have now been replaced by a different (woody) fuel type that burns less often but with greater intensity (Wallace and others 2003). The additive effects of grazing by livestock and wildlife may reduce fine fuels and decrease fire effectiveness in controlling woody species (Briggs and others 2002a), and in some situations, fire may increase encroachment by evoking vigorous resprouting (Briggs and others 2005). Although fire events are predicted to increase with future climate change in western Canada (Flannigan and Van Wagner 1991), the ability of bog birch to survive a wide range of fire frequencies (DeGroot and Wein 1999), as well as tolerate warm post-fire conditions (DeGroot and Wein 2004), indicates this species will likely become more problematic in the future, and will be difficult to combat even with tools such as prescribed fire.
Decisions on when and whether to conduct a prescribed burn are currently based on the potential economic loss of timber allocated to forest companies, as well as the risk to energy extraction infrastructure on uplands adjacent to floodplain meadows. In contrast, it is difficult to attach a monetary value to grasslands that are contributing to livestock grazing, much less regional biodiversity and wildlife habitat. Cost considerations are a key determinant of the ability of public land administrations to undertake controlled burns, particularly when faced with the very real likelihood that restoration may initially require a more frequent fire regime than the historical one to restore the original grassland condition (see Briggs and others 2005). Heisler and others (2004) concluded that while fire restoration may prevent further encroachment of grasslands, the abundance of existing shrubs is unlikely to change. This may be the case in the current study as bog birch exhibits ecological inertia, with repeated fires unable to remove this species once established (Bork and others 1996). Despite the fact that fire may be the most natural tool for shrub control, the potential cost of dealing with an escaped prescribed burn often precludes its use, particularly at the spatial and temporal scale needed to effectively reverse shrub encroachment throughout the RMFR. However, should the conservation of grasslands in this region of Alberta be considered a priority, more resources would need to be made available and greater efforts directed into restoration.
Evaluation of the GIS/Field Validation Approach
The quality of photos used in this study influenced the observed pixel digital ranges associated with shrubland and grassland habitats. Furthermore, the aerial photographs from 1958 and 1998 were of superior quality than the 1974 photos. Despite this, the 1974 imagery resulted in quantified areas of grassland consistent with (i.e., intermediate between) those from 1958 and 1998. Nevertheless, the exact quantitative decrease in grassland associated with the 1974 data relative to 1958 should be used cautiously and is better interpreted as an overall trend. More specifically, the exact rate of grassland loss associated with the three dates may partly be a result of limited photo quality.
When determining the values of pixels on digital images assembled from a mosaic of aerial photos, there is risk of having different digital values for the same habitat feature across all photos. This was evident even with the high quality 1998 imagery. For example, where two photos were spliced together, two different grassland values were evident on either side of the splice. Although this situation led to a range of values for individual habitats (e.g., 166–194 for grasslands in the 1998 reclassification), the digital value cutoffs used to separate vegetation types appeared effective in retaining the identity of each type of habitat.
This investigation utilized a comparison of the area of grassland obtained separately from each of the 3 years of imagery. This was necessary because of the high spatial resolution of the scanned photographs (e.g., around 0.5 m2), and the error associated with image co-registration. As true change detection can only be done if images are accurately registered to one another (Singh 1989), which was obviously not the case here given the small size of pixels examined (i.e. sub-meter resolution) relative to registration error, this would preclude individual changes in pixel value (and interpreted vegetation type) to be determined. Moreover, because the study area had very few anthropogenic features in 1974, and even fewer in 1958, accurately registering the imagery across years was not possible. Although a true change detection analysis would have had the advantage of mapping discrete localized areas of shrub expansion across the study area, the procedure used here remained robust to the objective of assessing overall grassland loss across the study area, a process made possible by the fact that these grasslands and shrublands were surrounded by a matrix of conifer forest with differential reflectance.
Conclusion
Despite challenges in determining pixel values associated with shrubland and grassland, the results of this study indicate significant areas of grassland have been lost in this portion of the Clearwater Grazing Allotment within the RMFR of Alberta between 1958 and 1998. We found a marked 58% decrease in grassland across all sub-regions over the 40 years. This change also led to the loss of 38% of the area’s grazing capacity, including 4,101 AUMs of primary range for cattle and elk. Left unchecked, this change may intensify competition between herbivores, increase utilization within remaining grasslands, and subsequently reduce the future range health of native grasslands if ungulate numbers are not adjusted. Moreover, this trend toward a shrub dominated valley bottom is likely to continue with modern fire suppression efforts.
This study provides regional public land managers with important information regarding ongoing habitat change and associated ungulate foraging opportunities. In addition, this study establishes the degree of urgency for implementing various land management strategies consistent with conserving grassland habitats. Continued grassland loss will decrease the availability of unique vegetation types that contribute to regional diversity, and reduce wildlife habitat and commercial livestock grazing opportunities. In order to maintain existing grasslands as well as reverse the observed reduction in grassland, public land managers will need to proactively make difficult decisions regarding the use of prescribed burning or other shrub control practices, and ensure they effectively match ongoing forage demand from livestock and wildlife with forage availability in affected areas.
Notes
Nomenclature follows that of Moss (1983)
References
Adams BW, Castelli O, Gardner FG, Cartwright G, Clark J (1992) Fire and grazing to manage willow shrubland on foothill range. Range Notes 15:1–6. Pub. No. T/207. Lands and Forest Services, Edmonton, Alberta, Canada
Archer S, Scifres C, Bassham CR, Maggio R (1988) Autogenic succession in a subtropical savanna: conversion of grassland to thorn woodland. Ecological Monographs 58(2):111–127
Archer SR, Schimel DS, Holland EH (1995) Mechanisms of shrubland expansion: land use, climate or CO2? Climate Change 29:91–99
Bai Y, Broersma K, Thompson D, Ross TJ (2004) Landscape-level dynamics of grassland-forest transitions in British Columbia. Journal of Range Management 57:66–75
Bailey AW, Wroe AL (1974) Aspen invasion in a portion of the Alberta parklands. Journal of Range Management 33:37–40
Baker WL (1992) Effects of settlement and fire suppression on landscape structure. Ecology 73:1879–1887
Bonham CD (1989) Measurements for terrestrial vegetation. Wiley, New York, NY, 338 pp
Bork EW, Burkinshaw AM (2009) Cool-season floodplain meadow responses to shrub encroachment in Alberta. Rangeland Ecology and Management 62:44–52
Bork E, Smith D, Willoughby M (1996) Prescribed burning of bog birch. Rangelands 18(1):4–7
Bork EW, Adams BW, Willms WD (2002) Resilience of foothills rough fescue, Festuca campestris, rangeland to wildfire. Canadian Field Naturalist 116:51–59
Bragg TB, Hulbert LC (1976) Woody plant invasion of unburned Kansas bluestem prairie. Journal of Range Management 29:19–24
Briggs JM, Hoch JA, Johnson LC (2002a) Assessing the rate, mechanisms, and consequences of the conversion of tallgrass prairie to Juniperus virginiana forest. Ecosystems 5:578–586
Briggs JM, Knapp AK, Brock BL (2002b) Expansion of woody plants in tallgrass prairie: a fifteen year study of fire and fire-grazing interactions. American Midland Naturalist 147:287–294
Briggs JM, Knapp AK, Blair JM, Heisler JL, Hoch GA, Lett MS, McCarron JK (2005) An ecosystem in transition: causes and consequences of the conversion of mesic grassland to shrubland. BioScience 55:243–254
Brown JR, Archer S (1999) Shrub invasion of grassland: recruitment is continuous and not regulated by herbaceous biomass or density. Ecology 80:2385–2396
Bryant JP, Tahvanainen J, Sulkinoja M, Julkunen-Tiitto R, Reichardt P, Green T (1989) Biogeographic evidence for the evolution of chemical defense by boreal birch and willow against mammalian browsing. The American Naturalist 134:20–34
Burkinshaw AM (2005) Shrub encroachment onto floodplain meadows within the Rocky Mountain Forest Reserve, Alberta. MSc Thesis, University of Alberta, Edmonton, Alberta, Canada, 123 pp
Cook JG, Hershey TJ, Irwin LL (1994) Vegetative response to burning on Wyoming Mountain-shrub big game ranges. Journal of Range Management 47:296–302
Cormack RGH (1953) A survey of coniferous forest succession in the eastern Rockies. Forestry Chronicles 29(3):218–232
DeGroot WJ, Wein RA (1999) Betula glandulosa Michx. response to burning and postfire growth temperature and implications of climate change. International Journal of Wildland Fire 9:51–64
DeGroot WJ, Wein RA (2004) Effects of fire severity and season of burn on Betula glandulosa growth dynamics. International Journal of Wildland Fire 13:287–295
DeGroot WJ, Thomas PA, Wein RW (1997) Betula nana L. and Betula glandulosa Michx. Journal of Ecology 85:241–264
Dunwiddie PW (1977) Recent tree invasion of subalpine meadows in the Wind River Mountains, Wyoming. Arctic and Alpine Research 9(4):393–399
Edwards KJ, Schofield JE, Mauquoy D (2008) High resolution paleoenvironmental and chronological investigations of Norse landnám at Tasiusaq, Eastern Settlement, Greenland. Quaternary Research 69:1–15
Environmental Systems Research Institute (ESRI) (2002) Arcview version 3.X for windows. Redlands, CA, USA
Flannigan MD, Van Wagner CE (1991) Climate change and wildfire in Canada. Canadian Journal of Forest Research 21:66–72
Government of Alberta (2000) Forest reserves act. Chapter/regulation F-20 RSA 2000. ISBN#0779728203
Hanson WR (1973) History of the eastern rockies forest conservation board 1947–1973. Eastern Rockies Forest Conservation Board, unpublished Alberta Forest Service report. Edmonton, Alberta, 54 pp
Hebblewhite M, Merrill EH, McDonald TL (2005) Spatial decomposition of predation risk using resource selection functions: an example in a wolf-elk predator-prey system. Oikos 111:101–111
Hebblewhite M, Merrill EH, McDermid G (2008) A multi-scale test of the forage maturation hypothesis in a partially migratory ungulate population. Ecological Monographs 78:141–166
Heisler JL, Briggs JM, Knapp AK, Blair JM, Seery A (2004) Direct and indirect effects of fire on shrub density and aboveground productivity in a mesic grassland. Ecology 85:2245–2257
Knight CL, Briggs JM, Nellis MD (1994) Expansion of gallery forest on Konza Prairie Research Natural Area, Kansas, USA. Landscape Ecology 9(2):117–125
Köchy M, Wilson SD (2001) Nitrogen deposition and forest expansion in the northern Great Plains. Journal of Ecology 89:807–817
Lewis HT (1980) Indian fires of spring. Natural History Journal 89:76–81
MacDonald GM (1989) Postglacial palaeoecology of the subalpine forest-grassland ecotone of southwestern Alberta: new insights on vegetation and climate change in the Canadian Rocky Mountains and adjacent foothills. Palaeography, Palaeoclimatology, Palaeoecology 73:155–173
Mast NJ, Veblen TT, Hodgson ME (1997) Tree invasion within a pine/grassland ecotone: an approach with historical aerial photography and GIS modeling. Forest Ecology and Management 93:181–194
Moss EH (1983) Flora of Alberta. University of Toronto Press, Toronto, ON, 687 pp
Pengelly I, Hamer D (2006) Grizzly bear use of pink hedysarum roots following shrubland fire in Banff National Park, Alberta. Ursus 17(2):124–131
Petranka JW, McPherson JK (1979) The role of Rhus copallina in the dynamics of the forest-prairie ecotone in North Central Oklahoma. Ecology 60:956–965
Rayner MR (1984) Ecological land classification and evaluation: Ram–Clearwater. Resource evaluation branch, resources evaluation and planning division, Alberta energy and natural resources, Edmonton, Alberta, Canada
Schlesinger WH, Reynolds JF, Cunningham GL, Huennecke LF, Jarrell WH, Virginia RA, Whitford WG (1990) Biological feedbacks in global desertification. Science 247:1043–1048
Singh A (1989) Digital change detection techniques using remotely sensed data. International Journal of Remote Sensing 10:989–1003
Society for Range Management (1989) A glossary of terms used in range management, 3rd edn. Society for Range Management, Denver, CO, USA
Strong WL, Leggat KR (1992) Ecoregions of Alberta. Alberta Forestry, Lands and Wildlife, Resource Information Branch, Edmonton, Alta. T/245, 77 pp
Sturm M, Racine C, Tape K (2001) Increasing shrub abundance in the Arctic. Nature 411:546–547
Tueller PT (1989) Remote sensing technology for rangeland management applications. Journal of Range Management 42:442–453
Turner MG, Romme WH (1994) Landscape dynamics in crown fire ecosystems. Landscape Ecology 9:59–77
Van Auken OW (2000) Shrub invasions of North American semiarid grasslands. Annual Review of Ecology and Systematics 31:197–215
Wallace OC, Qi J, Heilma P, Marsett RC (2003) Remote sensing for cover change assessment in southeast Arizona. Journal of Range Management 56:402–409
Watkinson AR, Ormed SJ (2001) Grasslands grazing and biodiversity: editor’s introduction. Journal of Applied Ecology 38:233–237
Weerstra BG (1990) Elk creek rangeland reference area range use, history, condition and trend. Biota Consultants, Cochrane, Alberta. Forest Land Use Branch, Range Management Section, Edmonton Alberta, Canada
Weis IM, Hermanutz LA (1988) The population biology of the arctic dwarf birch, Betula glandulosa: seed rain and the germinable seedbank. Canadian Journal of Botany 66:2055–2061
Wessman CA, Archer S, Johnson LC, Asner GP (2004) Woodland expansion in U.S. grasslands: assessing land cover change and biogeochemistry impacts. In: Guttman G, Janetos A, Skole D (eds) Land change science: observing, monitoring and understanding trajectories of change on the Earth’s surface. Kluwer Academic Publishers, Dordrecht, Netherlands, pp 185–208
West NE (1993) Biodiversity of rangelands. Journal of Range Management 46:2–13
Willms WD, Smoliak S, Dormaar JF (1985) Effects of stocking rate on a rough fescue grassland vegetation. Journal of Range Management 38:220–225
Willoughby MG (2000) Rangeland reference areas: species composition changes in the presence and absence of grazing and fire on the rough fescue-hairy wildrye dominated community types of the Upper Foothills Subregion. Lands and forest services division, Edmonton, Alberta
Willoughby MG (2001) Range plant community types and carrying capacity guides for the Upper Foothills Subregion. Alberta Sustainable Development, Public Lands. Pub No. T/356. Edmonton, Alberta, Canada
Acknowledgments
This research was funded by Alberta Sustainable Resource Development, the Alberta Conservation Association, and the University of Alberta. We thank Drs. Peter Crown, Evelyn Merrill and Bob Hudson for their helpful comments on an earlier version of this manuscript, and thank Brent Finnestead for field assistance, as well as Rick Pelletier for assistance with the GIS analysis. We also thank 4 anonymous reviewers for helpful advice on improving the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Burkinshaw, A.M., Bork, E.W. Shrub Encroachment Impacts the Potential for Multiple Use Conflicts on Public Land. Environmental Management 44, 493–504 (2009). https://doi.org/10.1007/s00267-009-9328-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-009-9328-2