
Abstract
Reed beds of Phragmites australis in the River Amudarya delta near the Aral Sea constitute permanent breeding areas of the Asian Migratory locust, Locusta migratoria migratoria. Every year, thousands of hectares are treated with broad-spectrum insecticides to prevent locust swarms from damaging crops in adjacent areas. To devise efficient locust monitoring and management plans, accurate and updated information about the spatial distribution of reeds is necessary. Given the vast geographic extent of the delta, traditional, ground survey methods are inadequate. Remotely sensed data collected by the MODIS sensor aboard the TERRA satellite provide a useful tool to characterize the spatial distribution of reeds. Multi-temporal MODIS data, collected at different times of the growing season, were used to generate spectral–temporal signatures for reeds and other land cover classes. These spectral–temporal signatures were matched with reed phenology. MODIS information was digitally classified to generate a land cover map with an overall accuracy of 74%. MODIS data captured 87% of the ground-verified reed locations. Estimates derived from MODIS data indicate that 18% of the study area was covered by reeds. However, high commission error resulted from misclassification of reeds mixed with shrubs class and shrubs class as reeds. This could have resulted in overprediction of the area covered by reeds. Additional research is needed to minimize the overlap between reeds and other vegetation classes (shrubs, and reed and shrub mix). Nevertheless, despite its relatively low spatial resolution (250 m), multi-temporal MODIS data were able to adequately capture the distribution of reeds. Instead of blanketing the fragile wetland ecosystem of the Amudarya delta with chemical anti-locust treatments, plant protection specialists can use this information to devise ecologically sound pest management plans aimed at reducing the adverse environmental impact in the zone of the Aral Sea ecological catastrophe. MODIS methodology to identify reed stands can be applicable to the Migratory locust habitats in other geographic areas.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
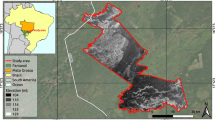
The stands of the common reed Phragmites australis (Cav.) Trin. ex Steud. in the Amudarya (Oxus) River delta near the Aral Sea in southwestern Uzbekistan (Fig. 1) harbor a permanent breeding area of the Asian Migratory locust, Locusta migratoria migratoria L. (Novitsky 1963). The dynamic delta ecosystem provides favorable conditions for locust survival and reproduction. The sandy river banks serve as oviposition sites, whereas the reed stands furnish food and shelter for nymphs and adults. Locust outbreaks occur there at irregular intervals (3–15 years), last for several years, and devastate the irrigated crops nearby (Latchininsky and Gapparov 1996). Locust swarms disperse in different directions covering distances over 1000 km (Tsyplenkov 1970). Usually, the outbreaks follow abnormally dry years when larger areas emerge from flooding, expanding the habitat suitable for oviposition. Every year, thousands of hectares are treated with broad-spectrum insecticides to prevent locust swarms from damaging crops in adjacent areas (Fig. 2).

Location of the study area (highlighted) south of the Aral Sea, Uzbekistan. The image backdrop is a Moderate Resolution Imaging Spectroradiometer (MODIS) scene acquired on July 19, 2004 and displayed in 2 (infrared), 1 (red), and 1 combination

Areas infested by and treated annually against the Asian Migratory locust in the Amudarya delta, per decade (1930–2000). Source: Archives of the Uzbek Plant Protection Service
Spatial distribution of reeds within the Amudarya delta is influenced by its hydrologic regime. Since the 1950s, the amount of water tapped for agricultural use from the Amudarya River has increased greatly. Diminished water flow in the mouth of the river has led to the rapid desiccation of the Aral Sea. By the end of the 20th century, it had lost more than half of its area and volume and moved down from the fourth to the eighth place in the list of the world′s largest lakes (Stone 1999). With the contraction of the Aral Sea, the area covered by reeds has also shrunk by two thirds (Gaparov and Latchininsky 2000). Numerous lakes and bogs have dried up, resulting in drastic changes in the delta′s ecology. Beside insights for locust monitoring, characterizing the spatial distribution of reeds could also provide valuable information about the hydrologic regime of the Amudarya River and better understanding of the interconnectivity of the components within this complex wetland ecosystem.
Satellite data collected at different times of the year, referred to as multi-temporal data, have been used for understanding vegetation phenology (Reed and others 1994) and mapping land cover features using vegetation seasonal change information (Lo and others 1986, Hirosawa and others 1996, Friedl and others 2002, Sedano and others 2005). Studies have demonstrated the utility of multi-temporal data sets to map agricultural crops (Odenweller and Johnson 1984, Mkhabela and others 2005), distinguish evergreen trees from deciduous trees (Reed and others 1994, Senay and Elliott 2000, Fairbanks and McGwire 2004) and measure productivity and carbon exchange (Veroustraete and others 1996, Ricotta and others 1999, Posse and Cingolani 2004). Vegetation response to the electromagnetic spectrum in the red and infrared regions is influenced by factors such as differences in chlorophyll content, nutrient levels, water content, and underlying soil characteristics. Reflectance values measured over time are converted to vegetation index values, such as ratios. These values are used to construct “spectral-temporal profiles” to identify the phenological events in vegetation or map vegetation types based on their phenological changes (Reed and others 1994).
Data obtained from Landsat Multi Spectral Scanner and Thematic Mapper sensors have also been used for such studies (e.g., Lo and others 1986). However, spectral–temporal profiles generated from Landsat data were found to omit certain phenological changes because of the relatively long revisit time (16 days) or presence of clouds. Advanced Very High Resolution Radiometer (AVHRR) data collected by the National Oceanic & Atmospheric Administration weather satellites enabled researchers to obtain daily information and conduct phenological change studies on land cover features (Justice and others 1985, Hirosawa and others 1996, Fairbanks and McGwire 2004). However, the coarse spatial resolution (1 km × 1 km) of AVHRR data limits their use for obtaining detailed spatial information about specific features on the ground.
With the availability of Moderate Resolution Imaging Spectroradiometer (MODIS) data, possibilities for land cover mapping and monitoring have increased because of their improved spectral, spatial, geometric, and radiometric attributes relative to AVHRR data (Friedl and others 2002, Townshend and Justice 2002). MODIS data are collected daily onboard the TERRA and AQUA satellites in 36 spectral bands at 250-, 500-, and 1000-m spatial resolution. Because of their wide swath width of 2330 km (Lillesand and others 2004), these data are suitable for monitoring and assessing land cover conditions over relatively large geographic scales, allowing for generating regional (Zhan and others 2002, Fensholt 2004) and global (Friedl and others 2002) land cover products. MODIS data collected in the red (620–670 nm) and infrared (841–876 nm) regions of the electromagnetic spectrum (bands 1 and 2, at 250-m spatial resolution) are useful for vegetation monitoring and mapping, because reflectance in these regions is influenced by numerous vegetation conditions (Campbell 2002). High temporal resolution provided by MODIS data is crucial to effectively capture changes in vegetation phenology (Borak and others 2000). Zhan and others (2002) concluded that 250-m MODIS data were useful in identifying changes in vegetative cover in Africa, America, and Asia. Fensholt (2004) found that the MODIS-based enhanced vegetation index was better correlated with leaf area index than the AVHRR-based vegetation index.
The objective of this study was to characterize the spatial distribution of reeds, which serve as Asian Migratory locust habitat in the Amudarya River delta. Timely and precise information on reed distribution would enable the plant protection specialists to devise efficient pest management plans and minimize the amount of chemicals introduced into the environment. Given the vast, more than 1,000,000-ha, geographic extent of the delta, generating up-to-date reed distribution information is impossible by conventional, ground-based survey methods. Remotely sensed data could be a viable source to provide the information on reed distribution on a periodic basis.
Study Area
The Aral Sea and Its Desiccation
The Aral Sea is a saltwater lake with no outlet, located in the arid interior of Central Asia, 450 km east of the Caspian Sea. In the early 1960s, the Aral Sea was the fourth largest lake on Earth, with a surface area greater than 6,950,000 ha and a volume of 1064 km3 (Precoda 1991). It was a shallow reservoir, with a maximum depth of 68 m and an average depth of only 15.5 m (Glazovsky 1990).
The Aral Sea was fed by two tributaries, the Amudarya in the south and the Syrdarya in the northeast. The Amudarya River formed a vast wetland delta (1,400,000 ha), comprising an intricate system of channels, ponds, temporary lakes, large and small rivulets, and islands. The hydrological regime of the Aral Sea was more or less stable until the end of the 1950s, with rains (3–9 km3 per year) and inflow from its two tributaries (55–56 km3 per year) compensating for the high evaporation losses (58–65 km3) (Mainguet and others 1995). Since the late 1950s, the water from the tributaries has been increasingly diverted for agricultural irrigation. The rapid expansion of the irrigated area from 5,600,000 to 8,500,000 ha led to an almost total extraction of the average annual inflow by the end of the 1980s (Perera 1993). This resulted in the rapid and irreversible desiccation of the Aral Sea, which lost >60% of its area and >80% of its volume by the year 2000. The shoreline in the most shallow, southeastern part had receded 80–120 km from its original littoral limit and exposed 3,600,000 ha of the former sea bottom (Stone 1999).
Native Vegetation and Its Changes
Before the desiccation started, the native vegetation of the Amudarya delta included two principal plant communities (Latchininsky and Gapparov 1996). A particular type of moist grassland, a virtual monoculture of the common reed (Ph. australis), covered 1,200,000 ha, or about 85% of the delta. The reeds formed dense stands of different heights (up to 6 m) depending on the soil moisture. Other plant species in the reed stands included sedges and rushes, and represented a minor proportion of the vegetation cover. The second vegetative community occupied the immediate vicinity of the main water bodies with a total area of 200,000 ha, or 15% of the delta. It was a fluvial forest–shrub mix composed of several native poplars (Populus spp.), elaeagnus (Elaeagnus spp.), salt cedar (Tamarix spp.), willows (Salix spp.), and saxaul species (Halimodendron halodendron, Haloxilon sp.) (Gapparov and Latchininsky 2000). Other vegetative communities occupied a tiny proportion of the delta. The zone of croplands was located outside of the delta, along the main course of the Amudarya. Rice, cotton, melon, and potatoes were the principal crops; near villages, small grains, vegetables, vineyards, and orchards were also cultivated.
The native wetlands of the Aral Sea basin have undergone dramatic changes resulting from the sea contraction. The area of reed beds shrunk by 70%. More than 30,000 ha of lakes and bogs have dried up completely (Gapparov and Latchininsky 2000). The fluvial forests along the rivers have disappeared almost entirely. Besides reeds, the vegetation is represented currently by sparse shrubs or reed–shrub mixture. A significant proportion of emerged soil remained devoid of any vegetation and became a salinized desert. Frequent dust storms aggravated the severe soil erosion.
Reeds, Their Phenology, and Role as Locust Habitat
The common reed is a perennial grass propagating mainly by rhizomes. In the Amudarya delta, it overwinters at a rhizome state. In late April or early May, the above-ground growth resumes. Within 6 to 8 weeks, the reed stands reach their maximum height. Flowering begins in July, and senescence starts in late August or early September. By the end of October, the above-ground reed vegetation dries out entirely, but the dry stems often remain standing through the winter. Reed productivity and density depend on the level of ground water: if the water is 1 m below the soil level, the reeds form a relatively dense stand with 80% soil cover and 1.9 t/ha of above-ground vegetation. In the drier areas where the ground water is 2 m below the soil level, the stands are sparse (60% cover) and the productivity is lower (1.4 t/ha). The reed stands that are submerged may form a 100% cover with very high productivity of more than 27 t/ha (Plisak 1981).
The univoltine life cycle of the Asian Migratory locust is synchronized with the development of its primary food plant, the common reed. Hatching occurs in late May. The nymphal development continues through early July, followed by adult emergence, sexual maturation, and mating. The females lay eggs from late July until September. The eggs overwinter in the soil and produce hatchlings the next May, when there are plenty of young reed plants for food. The primarily graminivorous locust is well adapted to the reed monoculture and plays a major role in the cycling of nutrients (Latchininsky and others 2002). However, it becomes a serious agricultural pest when the reed resources are depleted and the swarms fly out of the reed beds into the adjacent crop areas.
Locust Control Measures
The principal goal of locust control in the Amudarya delta is to prevent the development of the dense swarms capable of emigration flights toward crop areas. Monitoring and management of the locust populations are carried out by a special plant protection team of the Ministry of Agriculture and Water Resources of Uzbekistan. The team conducts locust surveys, identifies the infested areas, and implements control operations by spraying the infestations, which are confined primarily to reed stands. Locust infested areas averaged 300,000 ha per year, and during outbreaks, the areas of reeds treated with insecticides exceeded 500,000 ha annually (Fig. 2). The desiccation of the Aral Sea caused a severe reduction in the reed areas in the Amudarya delta, which decreased threefold by the end of the 20th century (Gapparov and Latchininsky 2000). Despite this contraction of the locust habitats, the ground surveys remain an extremely difficult task because of the lack of roads in the wetlands and the shortage of economic means. Furthermore, recent changes in the hydrological regime of the Amudarya resulted in the increase of water flow into the Large Aral. The subsequent expansion of the reed beds in the Amudarya delta and the former Aral Sea bottom enlarged the areas of locust habitats and increased the risk of locust infestations (Latchininsky and others 2002). Accurate and timely surveys became a necessity for effective locust management under tight financial constraints. Using satellite imagery for mapping potential locust-infested areas could provide a useful tool for local pest managers to ensure a better targeted and more efficient locust control.
According to the guidelines for the ground locust survey, the average daily area inspected by a professional scout comprises between 100 and 150 ha (Tsyplenkov 1970). This means that for a team of 20 scouts (the number of locust personnel working in the Amudarya delta), it will take 11 months to survey the entire 1,000,000-ha delta. In the real world, the nymphal survey period during which the infested areas are identified for subsequent treatments is limited to 3 to 4 weeks (Latchininsky and others 2002). Taking into account the paucity of resources combined with difficult accessibility and poor road network in the delta, the traditional, ground survey methods are obviously inadequate for efficient locust management.
Materials and Methodology
Satellite Data
MODIS single day, surface reflectance products (MOD09GQK) at 250-m spatial resolution acquired by Terra satellite in 2004 were downloaded from the U.S. Geological Survey–Land Processes Distributed Active Archive Center (LPDAAC) in HDF-EOS format. Bands 1 (620–670 nm) and 2 (841–876 nm) reflectance values were corrected for the effect of atmospheric gases, aerosols, and thin clouds (Jin and Sader 2006). Quality control data provided by the LPDAAC were used in combination with visual analyses to screen for clouds and other anomalies such as missing rows and stripes. From this, six single-day MODIS scenes obtained on April 30 (Julian day – 121), May 24 (145), June 17 (169), July 19 (201), August 18 (231), and September 21 (265) were selected. These dates correspond to the following phenological events of reeds: above-ground growth (late April or early May), maximum growth (mid- to late June), flowering (July), and senescence (late August or early September) (Plisak 1981).
Reference and Verification Data
Reference and verification data required for image classification and accuracy assessment were collected through interviews with local experts, two aerial reconnaissance missions, and three ground surveys in 2004.
Landsat data were digitally displayed using ViewFinder® software (ERDAS, Atlanta, GA), and the plant protection specialists were interviewed to obtain reference data (n = 65) and gain insights about reeds and other vegetation. Through this exercise, specific information about reeds and their spatiotemporal distribution within the delta was obtained. The plant protection personnel also provided valuable information about the relationship between water flow in the Amudarya River and its influence on the reed distribution. These data were used to aid image classification.
An ANTONOV-2 aircraft was used for the aerial reconnaissance missions to survey remote locations from altitudes ranging between 800 and 1000 m. Location information about reference points was collected with a Garmin (Garmin Ltd., Olathe, KS) Global Positioning System, along with digital photographs and description of the land cover type. Personnel from the Uzbek Plant Protection Institute assisted in conducting field surveys and collecting ground reference data. Landsat data acquired in 2002 were printed at 1:500,000 scale and used for navigation and site selection. Overall, 203 verification data were collected through these methods. This information was used for validating the classified images.
Classification Scheme
For this study, we developed a land cover classification scheme consisting of six classes to assign to pixels in the satellite imagery (Table 1). Based on the predominance of reeds in the vegetation cover, these six thematic classes have also been assigned different levels of risk as potential locust habitats.
Data Processing
Each MODIS scene was reprojected to Universal Transverse Mercator (UTM) projection (zone 40 North, WGS84 datum) in ERDAS Imagine 8.7 software and a subset corresponding to the study area, geographic center (59° 39′′ E; 44° 07′′ N), was generated. A vegetation index ratio (MODIS band 2 / MODIS band 1) for each scene was computed (Huete and others 2002). The vegetation index or simple ratio can provide valuable information about vegetation biomass as described by Schlerf and others (2005). From these six ratio bands, a composite image consisting of six vegetation index bands was generated. Using Iterative, Self-Organizing Data Analysis (ISODATA) classification technique, pixels in the composite image were grouped to generate classified images containing between 25 and 100 clusters. The ISODATA classification technique is a widely used method for generating pixel clusters based on spectral or temporal similarity (Lillesand and others 2004). Also classes 1 through 3 (Table 1) had some overlap in comparison to the rest of the classes; hence, identifying the cut-off on a consistent basis posed a challenge. Spectral distances between clusters in each classified image were analyzed, and the classified image with 50 clusters was found to be adequate (Lo and others 1986).
Using the mean vegetation index ratio value for each time step, spectral–temporal profiles were constructed for the 50 clusters (Lo and others 1986). Each cluster represented certain land cover class and methodologies outlined by Thenkabail and others (2000) and Sivanpillai and others (2005) were adopted to assign each cluster to a thematic class. Clusters were displayed sequentially and sample ground reference data were compared to the shape of the spectral–temporal profile at those locations. If the vegetation index values remained low (or were negative) throughout the year, it is more than likely that they corresponded to thematic classes other than vegetation (e.g., water or bare ground). For vegetation classes, the index values increased early in the growing season and might either remain steady or decline based on the plant growth and chlorophyll content. Using field data, clusters corresponding to vegetation classes were assigned to one of the thematic classes described in Table 1.
Accuracy Assessment
Ground verification data collected through ground visits and aerial surveys were used for assessing the accuracy of the classified multi-date MODIS image. A total of 203 verification sites were used for this analysis and to generate an error matrix (Congalton 1991, Jensen 1996). From this error matrix, the overall classification accuracy, kappa agreement index, and producer and user accuracies for each thematic class were computed. Overall accuracy and the kappa agreement index provide a measure of agreement between the thematic classes represented in the image and corresponding ground observed data. Producer accuracy for each thematic class represents the mapmaker′s accuracy and conveys the utility of the data and processing method. User accuracy for each thematic class provides a measure of map utility from a user′s perspective (Campbell 2002). With these values, map users can gain insights about the likelihood of finding the same class on the ground as represented in the map. To assess the utility of MODIS data for mapping reeds and other land cover classes, these values must be evaluated together.
Results
Multi-temporal MODIS Classification
Temporal signatures of reeds, sandy soil, and water were distinctly different in comparison to the temporal signatures of other thematic classes (Fig. 3). However, temporal signatures of shrubs, reed and shrub mix, and sparse vegetation had overlap. Reeds had higher ratio values among all vegetative classes throughout the study period (Fig. 3). Pixels corresponding to reed monoculture (class 1) had relatively low values in April and increased rapidly until July. This increase corresponds to the increase in biomass and plant vigor. Values declined after July, which corresponds to the senescence and decreased photosynthetic activity in reeds.

Temporal profiles for thematic classes in the study area (Reeds, Reed and shrub mix, Shrubs, Sparse vegetation, Sandy soil, Water). MODIS, Moderate Resolution Imaging Spectroradiometer
Pixels corresponding to mixtures of reeds and shrubs (class 2) had a similar temporal profile but lower in magnitude in comparison to reed monoculture (class 1). However, the temporal profiles for other vegetative classes overlapped and were not distinctly different from each other. Reference data corresponding to these classes (2: reeds and shrubs, 3: shrubs, and 4: sparse vegetation) were used to assign the clusters to one of these classes through the spatial overlay method.
Temporal profiles for sandy soil (class 5) were lower than for vegetation classes. However, the ratio values remained more or less constant throughout the study period (Fig. 3). Temporal signatures for water (class 6) had low values throughout the study period, because there is more absorption in the infrared region of the spectrum than in the red region.
Accuracy Assessment
The overall accuracy of the classified image was 74%, and most of the classification error could be attributed to the overlap between reeds and shrubs, and shrub classes (Table 2). The kappa agreement index was 0.68 (maximum value = 1.0 indicates complete agreement). Producer accuracy values for rest of the classes ranged between 77% (sandy soil) and 87% (reeds) (Table 3). The producer accuracy value for reeds indicates that the classification method used in this study was able to correctly identify 87% of the ground reference points. Multi-temporal MODIS data were successful in identifying sparse vegetation (84%), sandy soil (77%), and water (80%) in the study area. However, the producer accuracies for shrubs, reed, and shrub mix were 51% and 58%, respectively, indicating lower accuracy because of the overlap between these classes.
On the other hand, the user accuracy for reeds was only 63% (Table 3). Misclassification of shrubs, reeds, and the shrubs mixed class as reeds resulted in a lower user accuracy value. If 100 locations identified as reeds were to be selected from this map, only on 63 of those locations would the users find pure stands of reeds (class 1). In 25 locations, they would find shrubs, and reeds and shrubs mixed class. Using this classification, users could identify potential locust habitats of high and medium risk 72% (51 of 71) of the time and of medium and low risk 16% (12 of 71) of the time (Table 1). User accuracy values for shrubs, sparse vegetation, sandy soil, and water were higher than 75%, indicating that users of this map have higher probability of finding the class represented in the map on the ground.
Area Estimates and Spatial Distribution of Reeds
Based on the results obtained from classifying multi-temporal MODIS data, 18% (399,262 ha) of the total study area had homogeneous reed stands (class 1 in Table 3). These estimates included reeds that were growing on soil and in water bodies. The majority of the reed beds were found in wet soils, water bodies, and some in abandoned agricultural fields (Fig. 4).

Vegetation map depicting potential locust habitats, derived from multi-temporal Moderate Resolution Imaging Spectroradiometer (MODIS) data
Reeds mixed with shrubs (class 2) were found on 137,700 ha (or 6%) of the total area. The majority of the study area (40% or 897,131 ha) was represented by sandy soil (class 5), followed by sparse vegetation (22% or 490,681 ha; class 4).
The spatial resolution of MODIS bands 1 and 2 (250 m) was not adequate to identify the main course of the Amudarya River or other small channels and water bodies within the delta. At the same time, relatively large water bodies, which occupied several contiguous pixels, were identifiable from MODIS data. Most of the sandy soils were found along the edges (except the southern extremity) of the study area.
Discussion
Vegetation indices derived from the multi-temporal MODIS data were able to capture the phenological changes of reeds (Fig. 3). This pattern is very similar to the pattern exhibited by agricultural crops, where distinct phenological changes correspond to various growth stages such as leafing, maximum leaf-area, and senescence (Lo and others 1986, Ricotta and others 1999, Senay and Elliott 2000). The magnitude of change in reeds was more pronounced than in other vegetation classes found in this region. In addition to the unique temporal profile, reeds also grew mostly in large, continuous patches with more or less uniform growth pattern.
In a similar study conducted in the River Ili delta (Kazakhstan) using single-date Landsat imagery, reeds growing along the edges of water and in water bodies had a slightly different reflectance pattern than reeds growing on dry and wet soils (Sivanpillai and others 2006). However, in the Amudarya delta, reed stands were dense and in several instances completely covered the shallow water bodies. These areas do not pose high risk as a potential locust habitat because locusts will not lay their eggs in water. Satellite data acquired in winter or physiographic maps could be used to identify permanent water bodies. Using this information, reed stands submerged in water bodies could be separated from those that are growing on soils and assigned low risk in terms of potential locust habitat.
The temporal profile for those areas where reeds were mixed with shrubs was similar to the temporal profile of reeds, albeit with lower magnitude (Fig. 3). During field visits, predominance of a vegetation type at a site was the basis to assign them to classes 1 through 3. Some of the mixed reeds and shrubs sites that were misclassified as reeds could have more reeds in comparison to shrubs. In these circumstances, reflectance from reeds could have resulted in relatively higher values. However, these areas of reed and shrub mix pose more or less similar, medium risk in terms of potential locust habitat, and in most cases they are managed very similarly to the reed sites that present high risk.
Most of the shrubs and other vegetation growing in this region were not covering the soil as well as the reeds. Background soil was seen in several field sites; hence, reflectance for these classes could be a combined response of vegetation and soil. Consequently, these values were lower than reflectance of the dense, contiguous patches of reed stands. In some cases it was not possible to identify areas of pure shrub class because there were some reeds present in them. Seventeen of the 41 reference points corresponding to shrubs were either misclassified as reeds or as reeds and shrubs mixed class. If these maps were to be used for selecting locust treatment sites, care should be taken to avoid mostly shrub classes that might have few reeds within them.
In the southern portion of the study area, there were some agricultural fields, and reed stands were found along the boundaries of these fields. The relatively coarse spatial resolution of MODIS (250 m) was unable to separate these reed stands from agricultural crops because they had similar phenology. This could be a limitation of MODIS data in its ability to map reeds within an agricultural landscape. Phenological changes corresponding to crops and reeds might be similar and their temporal profiles might overlap, resulting in misclassification. Under these circumstances, relatively higher spatial resolution data such as Landsat, SPOT, or Indian Remote Sensing (IRS) Linear Self Scanning Sensor (LISS III) could be used. However, throughout the major part of the study area, there were no extensive agricultural sites, so the problem of discriminating between the reeds and the crop fields was of minor importance with regard to the locust habitat risk assessment.
MODIS data were able to identify reeds based on both phenology and magnitude of change in comparison to other vegetation classes (Fig. 4). This was particularly true for extensive patches of reed stands that were growing on soil or submerged in water. This methodology could be used for identifying and mapping the spatial distribution of reeds from the year 2000, corresponding to the launch of the satellite that carried the first MODIS sensor. This would enable the plant protection specialists to monitor spatiotemporal changes in reed distribution and growth on an annual basis. Results from this study could be applied to other wetland areas with large, continuous patches of reeds. Under these circumstances, 250-m spatial resolution of MODIS bands 1 and 2 would be adequate to monitor the phenological changes in reeds. Medium-resolution data such as Landsat or IRS satellite could be used to capture finer spatial details.
Identification of reed areas as potential Asian Migratory locust habitats is the key to successful management of this pest. Such areas constitute the main target for insecticide treatments. In the wetlands of the Amudarya River delta, ground identification of reed areas is difficult and rarely accurate. Consequently, insecticide treatments of locust infestations can miss the target and aggravate the environmental problems in the area of the Aral Sea ecological catastrophe by unnecessary overtreatment of few accessible areas (Gapparov and Latchininsky 2000). Using the MODIS imagery, it was possible to locate the potential locust habitats in space, which allows for more efficient, economic, and environmentally benign management solutions.
The vast distribution range of Locusta migratoria is larger than in any other locust species (Uvarov 1977). Nonetheless, its permanent breeding areas are strikingly similar in Africa, Eurasia, or Australia, being confined to grasslands on light soils in the vicinity of water bodies fringed with tall reed stands (COPR 1982). Therefore, this method of inventorying the Asian Migratory locust habitats in the Amudarya River delta using MODIS multi-temporal imagery could be applicable to other geographic areas where the Migratory locust is a threat to agricultural production.
Literature Cited
Borak JS, Lambin EF, Strahler AH (2000) The use of temporal metrics for land cover change detection at coarse spatial scales. Int J Remote Sensing 21:1415–1432
Campbell JB (2002) Introduction to remote sensing. The Guilford Publishers, New York, 621 pp
Congalton RG (1991) A review of assessing the accuracy of classifications of remotely sensed data. Remote Sensing Envir 37:35–46
COPR (1982) The locust and grasshopper agricultural manual. Centre for Overseas Pest Research, London, 690 pp
Fairbanks DHK, McGwire KC (2004) Patterns of floristic richness in vegetation communities of California: Regional scale analysis with multi-temporal NDVI. Global Ecol Biogeography 13:221–235
Fensholt R (2004) Earth observation of vegetation status in the Sahelian and Sudanian West Africa: Comparison of Terra MODIS and NOAA AVHRR satellite data. Int J Remote Sensing 25:1641–1659
Frieldl MA, McIver MAF, McIver DK, Hodges JCF, Zhang XY, Muchoney D, Strahler AH, Woodcock CE, Gopal S, Schneider A, Cooper A, Baccini A, Gao F, Schaaf C (2002) Global land cover mapping from MODIS: Algorithms and early results. Remote Sensing Envir 83:287–302
Gapparov FA, Latchininsky AV (2000) In: Lockwood JA, Latchininsky AV, Sergeev MG (eds) Grasshoppers and grassland health. Kluwer Academic Publishers, Dordrecht/Boston/London, pp 31–59
Glazovsky NF (1990) The Aral crisis. Priroda 10–11:27–41 (in Russian)
Hirosawa Y, Marsh SE, Kliman DH (1996) Application of standardized principal component analysis to land-cover characterization using multitemporal AVHRR data. Remote Sensing Envir 58:267–281
Huete A, Didan K, Miura T, Rodriguez EP, Gao X, Ferreira LG (2002) Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sensing Envir 83:195–213
Jensen JR (1996) Introductory digital image processing: A remote sensing perspective. Prentice Hall, New Jersey
Jin S, Sader SA (2006) MODIS time-series imagery for forest disturbance detection and quantification of patch size effects. Remote Sensing Envir 99:462–470
Justice CO, Townshend JRG, Holben BL, Tucker CJ (1985) Analysis of the phenology of global vegetation using meteorological satellite data. Int J Remote Sensing 6:1271–1318
Latchininsky AV, Gapparov FA (1996) Consequences of the drying of the Aral Sea for the locust situation in the region. Sécheresse 7:109–113 (in French)
Latchininsky AV, Sergeev MG, Childebaev MK, Chernyakhovskiy ME, Lockwood JA, Kambulin VG, Gapparov FA (2002) The acridids of Kazakhstan, Central Asia, and adjacent territories. Association for Applied Acridology International/University of Wyoming, Laramie, 387 pp (in Russian)
Lillesand TM, Kiefer RW, Chipman JW (2004) Remote sensing and image interpretation. John Wiley & Sons, Hoboken, New Jersey
Lo THC, Scarpace FL, Lillesand TM (1986) Use of multitemporal spectral profiles in agricultural land-cover classification. Photogram Eng Remote Sensing 52:535–544
Mainguet M, Létolle R, Glazovsky N (1995) Drought in the Aral-Caspian region. Sécheresse 6:135–143 (in French)
Mkhabela MS, Mkhabela MS, Mashinini NN (2005) Early maize yield forecasting in the four agro-ecological regions of Swaziland using NDVI data derived from NOAA′s AVHRR. Agric Forest Meteorol 129:1–9
Novitsky VY (1963) Ecological conditions of the permanent breeding area of the Asian Migratory locust in the Amudarya delta. Entomologicheskoe Obozrenie 42:252–263 (in Russian)
Odenweller JB, Johnson KI (1984) Crop identification using Landsat temporal-spectral profiles. Remote Sensing Envir 14:39–54
Perera J (1993) A sea turns into dust. New Scientist 140:24–27
Plisak RP (1981) Vegetation changes in the River Ili delta. Nauka, Alma-Ata, 216 pp (in Russian)
Posse G, Cingolani AM (2004) A test of the use of NDVI data to predict secondary productivity. Applied Vegetation Sci 7:201–208
Precoda N (1991) Requiem for the Aral Sea. AMBIO 20:109–114
Reed BC, Brown JF, VanderZee D, Loveland TR, Merchant JW, Ohlen DO (1994) Measuring phenological variability from satellite imagery. J Vegetation Sci 5:703–714
Ricotta C, Avena G, De Palma A (1999) Mapping and monitoring net primary productivity with AVHRR NDVI time-series: Statistical equivalence of cumulative vegetation indices. Photogram Remote Sensing 54:325–331
Schlerf M, Atzberger C, Hill J (2005) Remote sensing of forest biophysical variables using HyMap imaging spectrometer data. Remote Sensing Envir 95:177–194
Sedano F, Gong P, Ferrao M (2005) Land cover assessment with MODIS imagery in southern Africa Miombo ecosystems. Remote Sensing Envir 98:429–441
Senay GB, Elliott RL (2000) Combining AVHRR-NDVI and landuse data to describe temporal and spatial dynamics of vegetation. Forest Ecol Manage 128:83–91
Sivanpillai R, Smith CT, Srinivasan R, Messina MG, Wu XB (2005) Estimating regional forest cover in east Texas using Enhanced Thematic Mapper (ETM+) data. Forest Ecol Manage 218:342–352
Sivanpillai R, Latchininsky AV, Driese KL, Kambulin VE (2006) Mapping locust habitats in River Ili Delta, Kazakhstan, using LANDSAT imagery. Agric Ecosystems Envir 117:128–134
Stone R (1999) Coming to grips with the Aral Sea′s grim legacy. Science 284:30–33
Thenkabail PS, Nolte C, Lyon JG (2000) Remote sensing and GIS modeling for selection of a benchmark research area in the inland valley agroecosystems of West and Central Africa. Photogram Eng Remote Sensing 66:755–768
Townshend JRG, Justice CO (2002) Towards operational monitoring of terrestrial systems by moderate-resolution remote sensing. Remote Sensing Envir 43:351–359
Tsyplenkov EP (1970) Harmful Acrididae of the USSR. Kolos Publishers, Leningrad, 272 pp (in Russian). English translation 1978: Amerind Publishing Co., New Delhi/Bombay
Uvarov BP (1977) Grasshoppers and locusts. A handbook of general acridology. Vol. 2. COPR, London, 613 pp
Veroustraete F, Patyn J, Myneni RB (1996) Estimating net ecosystem exchange of carbon using the normalized difference vegetation index and an ecosystem model. Remote Sensing Envir 58:115–130
Zhan X, Sohlberg RA, Townshend JRG, DiMiceli C, Carroll ML, Eastman JC, Hansen MC, DeFries RS (2002) Detection of land cover changes using MODIS 250 m data. Remote Sensing Envir 83:336–350
Acknowledgments
Dr. Hans Wilps, Project leader - “Sustainable Control of Pest Locusts in Central Asia”, GTZ, Tashkent, Uzbekistan, coordinated the field work and aerial reconnaissance missions and his contributions to all aspects of this project are gratefully acknowledged. Ms. Evelin Kirkilionis-Wilps acquired the aerial photographs that were vital for the classification of MODIS data. Contributions of the following GTZ staff: Mr. Pawel Dobrynin, Ms. Mukhabhat Kamalova, and Mr. Nusamatdin Saparniyasov are also acknowledged. We thank Mr. Koshkimbay Djemuratov, Director, Locust Control Center, for his help with the field work. International travel assistance to the first author was provided by the International Programs Office, University of Wyoming. Partial salary support to the first author was provided by the USGS/AmericaView program through UW grant number: AMVIEW46014. International travel assistance to the second author was provided through a grant from NASA/Upper Midwest Aerospace Consortium (NDUNV48601LTCHN) and GTZ. We thank Dr. Jeffrey A. Lockwood, Dr. Kirk A. VanDyke, Dr. Kenneth L. Driese and three anonymous reviewers for their valuable suggestions.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Sivanpillai, R., Latchininsky, A.V. Mapping Locust Habitats in the Amudarya River Delta, Uzbekistan with Multi-Temporal MODIS Imagery. Environmental Management 39, 876–886 (2007). https://doi.org/10.1007/s00267-006-0193-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-006-0193-y