
Abstract
The impacts of land use and land cover (LULC) change in buffer zones surrounding protected ecological reserves have important implications for the management and conservation of these protected areas. This study examines the spatial and temporal patterns of LULC change along the boundary of Rio Abiseo National Park in the Northern Peruvian Andes. Landscape change within four ecological zones was evaluated based on trends expected to occur between 1987 and 2001. Landsat TM and ETM imagery were used to produce LULC classification maps for both years using a hybrid supervised/unsupervised approach. LULC changes were measured using landscape metrics and from-to change maps created by post-classification change detection. Contrary to expectations, tropical upper wet montane forest increased despite being threatened by human-induced fires and cattle grazing of the highland grasslands inside the park. Within the park’s buffer zone, tropical moist forest remnants were fragmented into more numerous and smaller patches between 1987 and 2001; this was in part due to conversion into agricultural land. The methods used in this study provide an effective way to monitor LULC change detection and support the management of protected areas and their surrounding environments.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Land use has been recognized as one of the major drivers of global change in biodiversity over the past several decades (Sala and others 2000; Hansen and others 2004). Over this same period, protected areas have expanded globally in order to conserve biological and cultural resources (Rodríguez and Young 2000; Chape and others 2003; Zimmerer and others 2004). It is recognized that protected areas are effective in protecting some kinds of biodiversity (Bruner and others 2001) although extinction of native species still occurs inside reserves (Woodroffe and Ginsberg 1998). In addition, land use/land cover (LULC) change surrounding a reserve can reduce its conservation capacity, and loss of biodiversity inside a protected area may be attributed to the size or isolation of the reserve (Baker 1989; 1992; Hansen and Rotella 2001; Hansen and others 2004; DeFries and others 2005). Buffer zones with limited or restricted land use have been suggested as a management strategy to reduce the influence of surrounding land-use activity on biodiversity within the protected area (Schonewald-Cox and Bayless 1986; Schonewald-Cox 1988; Byers 2000; Lynagh and Urich 2002).
Human-driven change occurs along trajectories of land use change proceeding from wildlands to a completely developed or urban stage (Hansen and others 2004). These landscapes may be undergoing multiple phases of transition or reversals resulting in a mosaic of native habitat remnants, agriculture, and urban environments. These land-use changes can result in loss of biodiversity due to homogenization of vegetation types (Flather and others 1998), susceptibility to edge effects where habitat fragmentation has occurred (Laurance and others 2001; Harper and others 2005), or an increase in the degree of human activity such as hunting, logging, grazing, or agriculture (Brashares and others 2001). Causes of landscape change and conservation strategies to protect biodiversity will vary, but it is apparent that economic, political, and conservation forces at local, national, and global scales are drivers of land cover change (Homewood and others 2001; Jokisch and Lair 2003) and influence the designation and management of protected areas (Zimmerer and others 2004; Bonta 2005).
Much attention has been paid to LULC change within the lowlands of the Amazon basin (e.g., Skole and Tucker 1993; Alves and others 1999; Evans and others 2001), while relatively little attention has been paid to the Andes, though they are recognized as globally important for biological diversity (Stotz and others 1996; Young and León 1999; 2000; Rodríguez and Young 2000). In the tropical Andes, landscapes have been profoundly shaped by millennia-old land use practices (Gade 1973; Young 1998; Sarmiento 2003), and the role of humans in adding to, reducing, or maintaining that diversity will, therefore, be a central issue in terms of conservation goals and practices in this region. People living within the buffer zone of a protected area could change the frequency and type of disturbances and thus alter disturbance regimes and associated vegetation types. However, little research has been done to evaluate the nature and consequences of human-induced LULC change on protected areas in this region. Indeed, few monitoring tools are available to detect when and where change may be important.
A change detection and landscape pattern metric study was conducted to evaluate the implications of LULC change along the boundary of Rio Abiseo National Park, located in the northern Peruvian Andes Figure 1. Several years after the park’s establishment in 1983; Young (1993a) obtained information on the vegetation types and land use systems within the district of Pataz, which is located inside the western buffer zone of the park. He delineated four ecological zones across the cordillera where the park and human settlements are located, and provided descriptions of land use and vegetation types for each zone. Before the change detection of the current study was conducted, certain expected landscape changes were predicted (Table 1). It was expected that fires set by humans would inhibit growth of wet montane forest inside the park and expansion of agriculture would result in fragmentation of moist montane forest remnants, resulting in a loss of cover of these native but unprotected habitats.

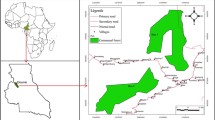
Study area in north-central Peru. Shown is Rio Abiseo National Park and the western buffer zone with five neighboring districts indicated, three of which were part of this study.
Analysis and monitoring of the rates and consequences of land-use change has been facilitated by satellite-based remote sensing (Skole and Tucker 1993; Jensen 1996; Hansen and others 2004; DeFries and others 2005), and information about land cover can serve as a proxy to evaluate effects on biodiversity based on knowledge of habitat preferences (Turner and others 2003). In this study, Landsat TM and ETM satellite imagery from 1987 and 2001 were acquired to monitor the status of landscape change along the boundary of the park several years after it became established. The analysis was done by (1) collecting field data in 2001, (2) performing from-to change detection of LULC classifications of the study area for 1987 and 2001, (3) evaluating LULC change within the study area and by ecological zone, and (4) measuring changes in the extent and configuration of cover types using landscape metrics.
Study Area
The ∼214,090-ha study area straddles the north-south oriented Eastern Cordillera of the northern Peruvian Andes extending from 7° 40′′ 36′S to 8° 05′′ 31′S and 77° 41′′ 07′W to 77° 16′′ 24′W (Figure 1). The area includes the western portion of Rio Abiseo National Park, which encompasses the upper section of the Huallaga River watershed, and the districts of Pataz, Pias, and Parcoy, which comprise the park’s western buffer zone where several towns and rural communities are located. Topography varies markedly on both sides of the cordillera with elevation ranging from ∼1100 to 4500 m.
The area is divided into ecological zones varying by elevation, climate, and vegetation. From east to west, the open tropical dry forest (1100–2300 m), the tropical moist montane (2300–3600 m), and the highland Puna grassland (3500–4500 m) zones extend across the western slopes of the cordillera. The Puna zone covers the formerly glaciated valleys and peaks of the upper cordillera and extends across and down the eastern side of the cordillera to ∼3700 m, transitioning into isolated forest patches and treeline, which marks the boundary of the wet montane forest zone below 3300 m (Young 1993a).
Humidity provinces of the Holdridge life zone system are dry (400 to 700 mm/yr), moist (700 to 1100 mm/yr), wet (1100 to 2200 mm/yr), and rain (>2200 mm/yr) (Holdridge 1967). There is an east to west directionality to precipitation; the leeward Marañon valley on the west side of the cordillera receives little precipitation from easterly tradewinds bringing moisture from the Amazon basin. The length of the dry season in the tropical dry forest zone is 8–10 mo/yr (May–January) and 3–5 mo/yr in the moist montane zone (June–October). Average daily minimum and maximum temperatures during the dry season are 25° and 33°C at ∼1100 m, and 15° and 25°C at ∼2700 m, respectively. Minimums and maximums are cooler, around 10° and 15°C, respectively, during the wet season (Young and León 1999).
On the eastern slopes of the cordillera, prevailing tradewinds, convection, and orographic uplift cause the formation of rain clouds over the tropical wet montane forest inside the park and the dry season is less than 1 mo/yr during the months of May through October. These forests are often exposed to severe weather during the rainy season when windstorms are common. Hail and frost occur above treeline and frost is especially likely during the dry season when skies are clear at night (Young 1993a). Mean annual temperatures range from 7° to 15°C in the upper montane forest belt (2500–3300 m) and from 15° to 19°C in the lower montane forest belt (1500–2500 m) (APECO 1999).
Land Use and Land Cover by Zone
The tropical dry forest zone includes scrub and cactus vegetation, along with a tall deciduous tree, Eriotheca discolor (Bombacaceae). Extensive stands of dry tropical forest are still relatively intact because agriculture is largely limited to irrigated sites around tributary outlets draining into the Marañon River. Houses dispersed across alluvial fans are surrounded by fruit-bearing orchards interspersed with maize and coca crops. Domesticated animals such as sheep and goats forage the native scrub.
The tropical moist montane ecological zone includes shrub and thorn woodland, as well as remnants of a tropical moist forest dominated by Alnus acuminata and Weinmannia glabra (Young 1993a; APECO 1999). The majority of native forest has been cleared and converted perhaps centuries ago into scrub, eucalypt plantations, or crops including maize, wheat, lentils, oca, alfalfa, barley, and potatoes. In 1961, there were 5, 4, and 23 people per km2 in the districts of Pataz, Pias, and Parcoy, respectively. In 1999, the population had increased, with respective densities of 13, 6, and 34 people per km2. Small villages and towns sit on plateaus at ∼2700 m, while more rural and dispersed agro-pastoral settlements exist on slopes above this elevation. Land use consists predominantly of artesanal and commercial mining, field agriculture, and grazing for sheep, goats, cattle, horses, and mules (Young 1993a, informal discussions during fieldwork 2001).
The Puna grassland zone is dominated by a bunch grass formation that prevails at heights above 3500 m in the central Andes Mountains (Young and others 1997). Bunch (tussock) grasses such as Calamagrostis, Cortaderia, and Festuca become dominant within this zone. Wetlands exist within valley bottoms and on slopes where soils can remain saturated from high rainfall throughout the year. Exposed rock exists above 4000 m, although grasses continue to grow above this elevation and cover most ridges despite nightly frosts and a large diurnal range in temperature (Young 1993a; Young and others 1997). Land use mainly involves cattle grazing; Puna grassland is occasionally burned to trigger regrowth of palatable grasses. It is now considered illegal to set fires inside the park, but grassland fires set outside the park’s boundary can be uncontrollable and spread across tens of square kilometers depending on moisture and wind conditions. Fire scars extending to wet montane treeline show evidence of plant mortality along the edge of this zone (León and others 2001).
The wet montane forest zone includes forest patches that exist along the slopes of glaciated valleys between 3300 to 3700 m (Young 1993b; 1993c). Although treeline elevation varies markedly across the Andes, forest within the study area continues below ∼3300 m down into the Amazonian lowlands where wet montane forest transitions into tropical rainforest. Forest structure at treeline is heterogeneous with 86 species of scandent plants and 133 species of trees (10 to 15 m) and shrubs (∼2 m). Below treeline, the upper montane forest belt has an understory typically dominated by bamboos of the genus Chusquea; above 2000 m, forest structure is complex with an irregular open canopy 10 to 25 m tall. Below 2000 m, the lower montane forest belt has a forest canopy 20–35 m tall and supports a high diversity of flora and fauna with 30 to 60 tree species per hectare (Young 1993b; 1993c; Young and León 1999).
Rio Abiseo National Park has a surface area of ∼274,520 hectares and includes both Puna and wet montane forest. It was established in 1983 after an assessment of its diverse natural and cultural resources (Young and others 1994; Leo 1995) and was added to Unesco’s World Heritage List in 1990. Before the park was created, local residents occasionally used the forest as a source of timber, medicinal plants, and game (Young 1993a; APECO 1999; Figueroa and others 2000). Potential threats to the park’s integrity include cattle grazing, fires triggered by humans, theft or destruction of archeological sites, local or commercial interest in opening the area to mining, and possible conversion of premontane forest for coca cultivation (Young 1993a; Young and others 1994; Young and others 1997; Young and León 1999).
Methods
Landsat 5 TM and 7 ETM satellite imagery were selected for LULC classification of the study area during a period when change in land cover by human activity has been recognized as an important driver of global environmental change (Lepers and others 2005). Imagery was obtained for May 15, 1987 and June 30, 2001 for path/rows 08/065 and 08/066. Selection of this imagery was based on minimal cloud cover, time of year, and the timeframe in which land cover change could be monitored since the designation of the protected area. Image pre-processing, classification, change detection, and GIS analysis were done using ERDAS IMAGINE 8.4 and ESRI’s ArcGIS 8.2. Ground control points collected in the field were used to rectify the 2001 pair of scenes. The 1987 pair was rectified to the 2001 image via image-to-image registration. Both pairs were resampled using the nearest-neighbor algorithm with RMS errors under 0.5. The pairs were mosaicked and subset to the study area. Atmospheric corrections were not done to the satellite data as the datasets were classified separately (Song and others 2001).
A classification scheme with LULC classes unique to the study area was developed prior to going into the field (Table 2) based on interpretation of the imagery, descriptions of the study area (e.g., Young 1993a; Young and León 1995; Young and others 1994; APECO 1999; Figueroa and others 2000), and cover type descriptions by Anderson and others (1976). Eleven cover types were selected for mapping at medium resolution using 30-m Landsat imagery. Orchards were separated from the agriculture class due to its unique shape, height, forest structure, and spectral characteristics. Field data were collected during the summer of 2001 with the help of a local informant. A stratified sampling scheme was developed based on collecting a number of points proportionate to the dominance of a class on the landscape. The collection of samples for certain classes was limited, however, due to the remoteness and inaccessibility to sites inside the study area.
The Landsat imagery was stratified by ecological zone to be classified separately in order to reduce class confusion errors by isolating homogenous landscape units (Hutchinson 1982; Elumnoh and Shrestha 2000; Heggem and others 2000; Hubert-Moy and others 2001). Classifications of the 1987 and 2001 imagery were produced for each stratified layer using a hybrid unsupervised/supervised classification approach (Messina and others 2000). The imagery was initially classified into 256 classes or signatures using the unsupervised Isodata algorithm. The signature file was edited within a transformed divergence table to remove signature pairs with low spectral separability values to maximize the variance between signatures (Jensen 1996; Messina and others 2000). The edited training signatures were used for supervised classification. The resulting LULC raster files were clumped and speckle pixels were removed using a 4 × 4 pixel filter. Thematic classes were coded according to the classification scheme (Table 2) and the stratified units were mosaicked together.
Several landscape metrics useful as ecological indicators of landscape composition and configuration (Forman and Godron 1986; Heggem and others 2000; Turner and others 2001) and commonly used within the literature (e.g., Hargis and others 1998; Zomer and Ustin 1999; McGarigal and others 2001; Southworth and others 2002) were chosen to analyze the character of landscape change as measured by the classification maps. Edge density, mean nearest-neighbor distance, contagion, and perimeter-area fractal dimension were computed for the study area as a whole and for each of the stratified landscape units using the software FRAGSTATS 3.3 (McGarigal and Marks 1995). An edge distance of 30 m was used to compute the total core or interior of a patch that is hypothetically unaffected by the dynamics of the surrounding ecotone. While edge effects may extend further into a patch interior (e.g., Reed and others 1996; Baker and Knight 2000), a 30-m edge corresponds with the edge influence distance measured by Young (1993b) and Young and Keating (2001) in the upper tropical montane forests of Peru and Ecuador, respectively.
Results
LULC classification maps of the study area were produced with eleven cover types for both 1987 and 2001 (see Figure 2 for examples). An error matrix for the 2001 LULC classification shows that of 295 GPS points taken in the field, 229 samples agreed with the 2001 product (Table 3). Classes with the lowest producer’s accuracy (most errors of omission) include the mixed grass/rock, agriculture, and moist montane forest classes. Classes with the lowest user’s accuracy (most errors of commission) include the wet montane shrub, wetland, and exposed surface classes. Overall accuracy was 78% with a Kappa statistic of 73%, which is below the generally accepted standard of 85%. Kappa is a commonly accepted measure of agreement between reference and classified data, but information provided by this statistic is limited in that it does not distinguish between quantification error and location error (Pontius 2000; 2002). Possible reasons for the classification errors include confusion between classes caused by the topographic effect (Colby and Keating 1998; Helmer and others 2000), an insufficient number of samples for certain classes (Congalton 1991), misclassification of mixed pixels at the edges of class boundaries (Frohn 1998; Foody 2002), or inaccurate ground control points resulting in image-to-image registration errors (Jensen 1996; Congalton and Green 1999).

Selected classes of the LULC classification of the study area for 1987 and 2001. District capitals are indicated.
Areas by cover type for 1987 and 2001 with amount and percentage of change during this time period are shown in Table 4, and a from-to change matrix indicating change trajectories is shown in Table 5. Urban, orchards, tropical wet montane forest, wet montane shrubland, and mixed exposed surface increased in area while agricultural land, moist montane forest, dry/moist shrubland, puna grassland, wetland, and mixed grass/rock decreased between 1987 and 2001. Dry/moist shrubland and puna grassland were the relatively dominant classes, comprising ∼24% and ∼32% of the study area by 2001, respectively.
Despite the expectation that wet montane forest growth would have been inhibited by fire inside the boundary of the park, this cover type actually increased in area by 23% from 6206 ha to 8606 ha between 1987 and 2001. Eighty-seven percent of forest area in 1987 remained stable in 2001. Totals of 1418 ha of wet montane shrubland, 457 ha of puna grassland, 452 ha of wetland, and 74 ha of grass/rock converted to forest, which expanded westward and upward into the puna grassland zone. Wet montane shrubs also expanded into areas that were previously puna grassland, wetland, and grass/rock, and 585 ha of wet forest converted to shrubland.
Only 35% of wetland area remained stable in 2001 although total amount of wetland declined slightly between 1987 and 2001 suggesting significant from-to change in both directions. Wetlands converted into wet montane shrub (937 ha), wet montane forest (452 ha), and mixed grass/rock (260 ha). This type of change suggests encroachment by wet montane forest and shrubland cover types into the puna grassland ecological zone. This trend is apparently a successional process of woody plants invading herbaceous vegetation.
Inside the park’s buffer zone where the districts of Pias, Pataz, and Parcoy are located, moist montane forest declined by 25% due to conversion to dry/moist shrub and agricultural land; 1648 and 246 ha converted into dry/moist shrubland and agricultural land, respectively. Over the same time period, 650 and 409 ha of dry/moist shrubland and agricultural land, respectively, reverted to moist montane forest. While forest expanded in certain areas and contracted in others, there was an overall trend of forest decline with forest converted predominantly into the shrubland class, which likely includes a certain amount of land that was cleared for future agricultural use.
Agricultural land decreased by approximately 20%, comprising ∼3.7% and ∼2.9% of the study area, respectively. It is likely, however, that the distribution and amount of agricultural lands fluctuate year to year because farmers use a 1 to 4 year rotational fallow system; certain crops and cropland may increase one year while decreasing in the next. Measuring the extent of agricultural land was also complicated by misclassification of recently cleared, fallow, or abandoned agricultural land as shrubland.
Landscape and Patch Metrics
For the study area, patch number, edge density, and total core area declined between 1987 and 2001 although these measures differ by ecological zone (Tables 6, 7). In the tropical dry forest zone, patchiness and total core area declined, but edge density changed only slightly. Increases in mean shape index and perimeter-area fractal dimension suggest that there were fewer but larger patches with more complex or irregular shapes. In the moist montane ecological zone, patch number and mean patch size did not change much although core area decreased, accompanied by an increase in patch shape complexity resulting in a higher edge density.
Fragmentation of patches occurred in the Puna grassland zone where the number of patches increased while mean patch size declined. However, total core area increased, which might be explained by the aggregation of certain patches as also suggested by a larger standard deviation in 1987 than in 2001. In the wet montane forest zone, located inside the park, patch number dropped by approximately 35%. Patches had less complex shapes with a larger core area and a decrease in edge density. This was the only zone where the Contagion metric, a measure that shows the degree of aggregation or clumping of patches, increased.
Patch dynamics differed by class within each zone (Table 8, Figure 3). Agricultural land, which is distributed across the moist montane ecological zone and to a lesser degree within the tropical dry forest zone, declined. Patch number decreased from a total of 1579 patches in 1987 to 1255 in 2001. Even though the average size of an agricultural patch increased by about 3%, mean core area declined by 20% in 1987 and 2001; respectively. The edge density of agricultural land also differed according to where patches were located. It increased only slightly in the moist montane zone, but decreased by 37% in the dry forest zone. This change suggests that agricultural land was becoming more aggregated and concentrated inside the moist forest zone.

Selected landscape pattern metrics for each ecological zone for 1987 (shaded) and 2001 (unshaded). The bottom axes refer to the eleven LULC classes.
Moist montane forest patches were fragmented into smaller pieces between 1987 and 2001. Moist forest area declined by 4% within the moist montane ecological zone, while total number of patches increased by 8%. These patches were smaller and had less core area. An increase in the edge density, shape index, and fractal dimension suggests that shape complexity increased. It is likely that clusters of remnant patches remained in the most inaccessible areas such as steep tributary ravines, while individual patches located on more gradual slopes were converted into other cover types for agriculture or grazing.
Inside the park, wet montane forest patches became larger, more numerous, and more aggregated. Patch number increased by 4% within the wet montane zone and by 607% in the puna grassland zone, providing evidence of forest expansion and new forest patch establishment in the puna grassland ecological zone over the studied time period. Mean patch size for patches within each zone varied, however. For example, mean patch size of forest within the wet montane zone was 8.2 ha in 1987 and increased to 9.5 ha in 2001. Mean patch size in the puna grassland zone, on the other hand, was only 0.2 ha in 1987, which increased to 1.3 ha in 2001. Edge density and core area of forest patches increased in both ecological zones. Forest interior, therefore, increased at a higher rate relative to the amount of edge forest adjacent to other cover types. Total core area increased by around 20%, which is surprising, especially when compared to the loss of native moist montane forest on the western slopes of the cordillera, outside the park.
Discussion
This study shows that significant landscape change occurred within the western buffer zone of Rio Abiseo National Park over a relatively short amount of time. Given an increase in human population between 1987 and 2001, we expected that land use activities would increase resulting in changes in the composition and configuration of landscapes within the study area. In the tropical moist forest zone, we predicted a continuing land-use trajectory of change due to agricultural intensification. A trend of smaller and fewer native forest patches was, therefore, expected. In the highlands, a more heterogeneous landscape with a patchwork of grassland and wetland habitats in varying stages of succession was expected due to periodic fires set by ranchers. At the edge between Puna grassland and wet montane forest, we predicted that forest or shrubland growth at treeline would be inhibited due to mortality of forest edge species by human-triggered fire.
Results show that certain land use changes were contrary to expectations. Inside the park, tropical wet montane forest expanded in area and timberline extended westward contrary to our prediction. It is likely that the reduction in fires since the establishment of the park has minimized the impact of this disturbance within the Puna and wet montane forest ecological zones although perhaps forest expanded as a consequence of climate change regardless of fire disturbance. Further studies are merited to (1) measure forest change before the park was established to compare with results from this study, (2) evaluate the effects of global climate change on upper wet montane forest over time, and (3) determine the frequency of fires set before and after the park was designated. While it was observed that recent fires have influenced edge habitat around wet montane forest patches and treeline, landscape metric results suggest that the threat of fire on forest growth was less than anticipated.
Inside the park’s buffer zone, agriculture apparently decreased somewhat in area. As mentioned, quantification of agricultural land was complicated by the possibility of misclassification between fallow land and shrubland. A more conclusive finding, perhaps, was the confirmation that patches of remnant moist montane forest were fragmented into more patches with a smaller core area. It is of concern that moist montane forest area declined by 25% because native moist montane forest is not likely to return to its 1987 extent without conservation measures to protect this type of habitat. The cause of this transformation is unclear from an interpretation of the classification products, but based on personal observations, we suggest that the land was likely cleared for agricultural use.
A possible implication of this type of change is that the influence of the surrounding habitat may contribute to further forest degradation and the loss of biodiversity (Saunders and others 1991; Laurance and others 2002; Freeman and others 2003). It is likely that certain species will be negatively affected given the contrast in structure and composition between forest habitat and agricultural land (Harper and others 2005). The loss of moist montane forest habitat may also have implications for biodiversity inside the park if there are species that have geographic ranges or populations that extend beyond the park’s boundary (Hansen and Rotella 2001; Armenteras and others 2003). It is recommended that these LULC changes with the park’s buffer zone be considered when conservation strategies are developed, keeping in mind the diverse needs and interests of the local stakeholders (Groom and others 1999; Wells and McShane 2004).
This type of LULC change study has several limitations. First, change detection with satellite imagery cannot account for all land use activities that might influence biodiversity or alter the biophysical conditions of a region. For example, subsurface mining within the buffer zone of Rio Abiseo National Park could not be monitored directly although it has changed local hydrologic systems and contributed to the degradation of a lake ecosystem in the study area. Second, the satellite study could not distinguish between native forests and forest plantations. In this case, eucalypt plantations, located in the moist montane forest zone, may have been classified as native forest remnants, and the amount of remaining native forest habitat measured has been potentially overestimated. Third, the accuracy of the 2001 classification was below the standard 85%, possibly due to registration errors, confusion between classes from effects of topography and shadow, or insufficient sample sizes as a consequence of conducting field work in a rugged and remote area.
This study of LULC change using satellite imagery provides a partial framework for monitoring land-use change trajectories in the tropics and elsewhere. The classification and stratification of Landsat 30-m satellite imagery made it possible to map land cover across a large, mountainous and remote area where other information was limited or unavailable. Mapping at this scale and extent also made it possible to monitor the park within its surrounding regional context to better understand the landscape ecological dynamics that extend across the political boundary of the protected area. The landscape pattern metrics in particular offer a rich source of additional information on the types of change trajectories. Results suggest that wet montane forest habitat is being maintained inside the park while land use inside the buffer zone itself is potentially leading to the degradation of remaining natural habitats.
Given severely limited park management resources, it is understandable that conservation strategies are focused on minimizing the impacts of human activity inside the park. It has been recognized, however, that protected habitats are influenced by their surrounding environment. This study has shown that significant land cover change occurred between 1987 and 2001 as a result of land use within the buffer zone of the park, and that one of the more significant threats to biodiversity is fragmentation of moist montane forest remnants. We, therefore, recommend that conservation managers consider strategies to minimize further loss of biodiversity within this unprotected habitat type. We suggest that conservation management within the park’s buffer zone will support conservation of biodiversity inside the park itself. This type of remote sensing study can (1) give insight into the effects of LULC change on this protected area and its buffer zone, (2) provide park mangers with information to target those land use change trajectories that conflict with the goal of conserving biodiversity inside Rio Abiseo National Park, (3) can be used as a monitoring framework for future change evaluations, and (4) can be used in other regions to evaluate the effects of LULC change on biodiversity inside and surrounding protected areas.
Literature Cited
Alves D. S., J. L. G. Pereira, C. L. D. Sousa, J. V. Soares, F. Yamaguchi. 1999. Characterizing landscape changes in central Rondonia using Landsat TM imagery. International Journal of Remote Sensing 20:2877–2883
Anderson, J. R., E. E. Hardy, J. T. Roach, and R. E. Witmer. 1976. A Land Use and Land Cover Classification System for Use with Remote Sensor Data. Geological Survey Professional Paper 964. United States Government Printing Office, Washington
APECO. 1999. Diagnostico Situacional para un Plan de Uso Publico del Sector Occidental del Parque Nacional Rio Abiseo y Zonas de Influencia. Asociación Peruana para la Conservación de la Naturaleza. PROFONANPE, Lima, Peru
Armenteras D., F. Gast, H. Villareal. 2003. Andean forest fragmentation and the representativeness of protected natural areas in the eastern Andes, Colombia. Biological Conservation 113:245–256
Baker W. L. 1989. Landscape ecology and nature reserve design in the boundary water canoe area, Minnesota. Ecology 70:23–35
Baker W. L. 1992. The landscape ecology of large disturbances in the design and management of nature reserves. Landscape Ecology 7:181–194
Baker W. L., R. L. Knight. 2000. Roads and forest fragmentation in the southern Rocky Mountains. In R. L. Knight, F. W. Smith, S. W. Buskirk, W. H. Romme, W. L. Baker (eds.), Forest fragmentation in the Southern Rocky Mountains. University Press of Colorado, Boulder. Pages 97–122
Bonta M. 2005. Becoming-forest, becoming-local: Transformations of a protected area in Hondouras. Geoforum 36: 95–112
Brashares J. S., P. Arcese, M. K. Sam. 2001. Human demography and reserve size predict wildlife extinction in West Africa. Proceedings of the Royal Society of London B 268: 2473–2478
Bruner A. G., R. E. Gullison, R. E. Rice, G. A. B. da. Fonseca. 2001. Effectiveness of parks in protecting tropical biodiversity. Science 291:125–128
Byers A. C. 2000. Contemporary landscape change in the Huascaran National Park and Buffer Zone, Cordillera Blanca, Peru. Mountain Research and Development 20:52–63
Chape, S., S. Blyth, P. Fox and M. Spalding. 2003. United Nations List of Protected Areas. IUCN, Gland, Switzerland and Cambridge, UK and UNEP-WCMC, Cambridge, UK
Colby J. D., P. L. Keating. 1998. Land cover classification using Landsat TM imagery in the tropical highlands: the influence of anisotropic reflectance. International Journal of Remote Sensing 19:1479–1500
Congalton R. G. 1991. A review of assessing the accuracy of classifications of remotely sensed data. Remote Sensing of Environment 37:35–46
Congalton R. G., K. Green. 1999. Assessing the accuracy of remotely sensed data : principles and practices. Lewis Publishers, Boca Raton, FL
DeFries R., A. Hansen, A. C. Newton, M. C. Hansen. 2005. Increasing isolation of protected areas in tropical forests over the past twenty years. Ecological Applications 15:19–26
Elumnoh A., R. P. Shrestha. 2000. Application of DEM data to Landsat image classification: Evaluation in a tropical wet-dry landscape of Thailand. Photogrammetric Engineering and Remote Sensing 66:297–304
Evans T. P., A. Manire, F. De Castro, E. Brondizio, S. McCracken. 2001. A dynamic model of household decision making and parcel level landcover change in the eastern Amazon. Ecological Modeling 143:95–113
Figueroa, C., A. Joaquina, J. Guerra, and K. Casas. 2000. Informe final del Estudio Socioeconomico y de uso de recursos de la poplacion aledana al Parque Nacional del Rio Abiseo: Monitoreo y Modelizacion de los Impactos de los Cambios de las Politicas Gubernamentales para la Conservacion de la Biodiversidad en Los Andes. APECO/BIOANDES, Lima
Flather C. H., M. S. Knowles, I. A. Kendall. 1998. Threatened and endangered species geography: Characteristics of hot spots in the conterminous United States. BioScience 48:365–376
Foody G. M. 2002. Status of land cover classification accuracy assessment. Remote Sensing of Environment. 80:185–201
Forman R. T. T., M. Godron. 1986. Landscape Ecology. Wiley, New York
Freeman R. E., E. H. Stanley, M. G. Turner. 2003. Analysis and conservation implications of landscape change in the Wisconsin River floodplain, USA. Ecological Applications 13:416–431
Frohn R. C. 1998. Remote sensing for Landscape Ecology: New Metric Indicators for Monitoring, Modeling, and Assessment of Ecosystems. Lewis Publishers, Boca Raton, FL
Gade D. W. 1973. Environment and disease in the land use and settlement of Apurimac department, Peru. Geoforum 16:37–45
Groom B., D. B. Jensen, R. L. Knight, S. Gatewood, L. Mills, D. Boyd-Heger, L. S. Mills, M. E. Soule. 1999. Buffer zones: Benefits and dangers of compatible stewardship. In M. E. Soule, J. Terborgh (eds.), Continental conservation: Scientific foundations of regional reserve networks. Island Press, Washington, D.C. Pages 171–197
Hansen A. J., J. J. Rotella. 2001. Biophysical factors, land use, and species viability in and around nature reserves. Conservation Biology 16:1112–1122
Hansen A. J., R. DeFries, W. Turner. 2004. Land use change and biodiversity: a synthesis of rates and consequences during the period of satellite imagery. In G. Gutman, C. Justice (eds.), Land change science: Observing, monitoring, and understanding trajectories of change on the earth’s surface. Kluwer Academic Publishers, New York, NY. Pages 277–299
Hargis C. D., J. A. Bissonette, J. L. David. 1998. The behavior of landscape metrics commonly used in the study of habitat fragmentation. Landscape Ecology 13:167–186
Harper K. A., S. E. Macdonald, P. J. Burton, J. Chen, K. D. Brosofske, S. C. Saunders, E. S. Euskirchen, D. Roberts, M. S. Jaiteh, P. Esseen. 2005. Edge Influence on Forest Structure and Composition in Fragmented Landscapes. Conservation Biology 19:768–782
Heggem D. T., C. M. Edmonds, A. C. Neale, L. Bice, K. B. Jones. 2000. A landscape ecology assessment of the Tensas River Basin. Environmental Monitoring and Assessment 64:41–54
Helmer E. H., S. Brown, W. B. Cohen. 2000. Mapping montane tropical forest successional stage and land use with multi-date Landsat imagery. International Journal of Remote Sensing 21:2163–2183
Holdridge R. L. 1967. Life Zone Ecology. Tropical Science Center, San José, Costa Rica
Homewood K., E. F. Lambin, E. Coast, A. Kariuki, I. Kikula, J. Kivelia, M. Said, S. Serneels, M. Thompson. 2001. Long-term changes in Serengeti-Mara wildebeest and land cover: Pastoralism, population, or policies? Proceedings of the National Academy of Sciences 98: 12544–12549
Hubert-Moy L., A. Cotonnec, L. Le-Du, A. Chardin, P. Perez. 2001. A comparison of parametric classification procedures of remotely sensed data applied on different landscape units. Remote Sensing of Environment 75:174–187
Hutchinson C. F. 1982. Techniques for combining landsat and ancillary data for classification improvement. Photogrammetric Engineering and Remote Sensing 48:123–130
Jensen J. R. 1996. Introductory digital image processing : A remote sensing perspective. Second Edition. Prentice Hall, Upper Saddle River, NJ
Jokisch B. D., B.M. Lair. 2003. One last stand? Forests and change on Ecuador’s Eastern Cordillera. Geographical Review 92: 235–256
Laurance W. F., T. E. Lovejoy, H. L. Vasconcelos, E. M. Buruna, R. K. Didham, P. C. Stouffer, C. Gascon, R. O. Bierregaard, S. G. Laurance, E. Sampaio. 2002. Ecosystem decay of Amazonian forest fragments: A 22-year investigation. Conservation Biology 16:605–618
Leo M. 1995. The importance of tropical montane cloud forest for preserving vertebrate endemism in Peru: The Rio Abiseo National Park as a case study. In L. S. Hamilton, J. O. Juvik, F. N. Scatena (eds.), Tropical montane cloud forests. Springer-Verlag, New York. Pages 198–211
León, B., K. R. Young, J. Roque, and D. B. Kintz. 2001. Evaluacion de la influencia antropogenica sobre la flora y vegetación en la zona montana alta del valle del Río Abiseo, Parque Nacional Río Abiseo. Informe Final de Investigacion. APECO, Asociación Peruana para la Conservación de la Naturaleza, Lima, Peru
Lepers E., E. F. Lambin, A. C. Janetos, R. DeFries, F. Achard, N. Ramankutty, R. J. Scholes. 2005. A synthesis of information on rapid land-cover change for the period 1981–2000. BioScience 55:115–124
Lynagh F. M., P. B. Urich. 2002. A critical review of buffer zone theory and practice: A Philippine case study. Society and Natural Resources 15:129–145
McGarigal, K., and B. J. Marks. 1995. FRAGSTATS: spatial pattern analysis program for quantifying landscape structure. General Technical Report, US Department of Agriculture, Forest Service
McGarigal K., W. H. Romme, M. Crist, E. Roworth. 2001. Cumulative effects of roads and logging on landscape structure in the San Juan Mountains, Colorado (USA). Landscape Ecology 16:327–349
Messina, J. P., K. A. Crews-Meyer, and S. J. Walsh. 2000. Scale-dependent pattern metrics and panel data analysis as applied in a multiphase hybrid land cover classification scheme. Proceedings 2000 ASPRS Annual Convention, Washington, D.C
Pontius R. G., Jr. 2000. Quantification error versus location error in comparison of categorical maps. Photogrammetric Engineering and Remote Sensing 66:1011–1016
Pontius R. G., Jr. 2002. Statistical methods to partition effects of quantity and location during comparison of categorical maps at multiple resolutions. Photogrammetric Engineering and Remote Sensing 68:1041–1049
Reed R. A., J. Johnson-Barnard, W. L. Baker. 1996. Contribution of roads to forest fragmentation in the Rocky Mountains. Conservation Biology 10:1098–1106
Rodríguez L. O., K. R. Young. 2000. Biological diversity of Peru: Determining priority areas for conservation. Ambio 29:329–337
Sala O. E., F. S. Chapin III, J. J. Armesto, R. Berlow, J. Bloomfield, R. Dirzo, E. Huber-Sanwald, L. F. Huenneke, R. B. Jackson, A. Kinzig, R. Leemans, D. Lodge, H. A. Mooney, M. Oesterheld, N. L. Poff, M. T. Sykes, B. H. Walker, M. Walker, D. H. Wall. 2000. Global biodiversity scenarios for the year 2100. Science 287:1770–1774
Sarmiento F. O. 2003. Anthropogenic change in the landscapes of highland Ecuador. Geographical Review 92:213–234
Saunders D. A., R. J. Hobbs, C. R. Margules. 1991. Biological consequences of ecosystem fragmentation: A review. Conservation Biology 5:18–32
Schonewald-Cox C. M. 1988. Boundaries in the protection of nature reserves. BioScience 38:480–486
Schonewald-Cox C. M., J. W. Bayless. 1986. The boundary model: A geographical analysis of design and conservation of nature reserves. Biological Conservation 38:305–322
Skole D., C. Tucker. 1993. Tropical deforestation and habitat fragmentation in the Amazon: Satellite data from 1978 to 1988. Science 260:1905–1921
Song C., C. E. Woodcock, K. C. Seto, M. P. Lenney, S. A. Macomber. 2001. Classification and change detection using Landsat TM data: When and how to correct atmospheric effects? Remote Sensing of Environment 75:230–244
Southworth J., H. Nagendra, C. Tucker. 2002. Fragmentation of a landscape: Incorporating landscape metrics into satellite analyses of land-cover change. Landscape Research 27:253–269
Stotz D. F., J. W. Fitzpatrick, T. Parker III, D. K. Moskovits. 1996. Neotropical birds: Ecology and conservation. University of Chicago Press, Chicago
Turner M. G., R. H. Gardner, R. V. O’Neill. 2001. Landscape ecology in theory and practice: pattern and process. Springer, New York
Turner W., S. Spector, N. Gardiner, M. Fladeland, E. Sterling, M. Steininger. 2003. Remote sensing for biodiversity science and conservation. Trends in Ecology and Evolution 18:306–314
Wells M. P., T. O. McShane. 2004. Integrating protected area management with local needs and aspirations. Ambio 33:513–519
Woodroffe R., R. G. Joshua. 1998. Edge effects and the extinction of populations inside protected areas. Science 280:2126–2128
Young K. R. 1993a. National park protection in relation to the ecological zonation of a neighboring human community: An example from northern Peru. Mountain Research and Development 13:267–280
Young K. R. 1993b. Woody and scandent plants on the edges of an Andean timberline. Bulletin of the Torrey Botanical Club 120:1–18
Young K. R. 1993c. Tropical timberlines: Changes in forest structure and regeneration between two Peruvian timberline margins. Arctic and Alpine Research 25:167–174
Young K. R. 1998. Deforestation in landscapes with humid forests in the central Andes: Patterns and processes. In K. S. Zimmerer, K. R. Young (eds.), Nature’s geography: New lessons for conservation in developing countries. University of Wisconsin Press, Madison. Pages 75–99
Young K. R., P. L. Keating. 2001. Remnant forests of Volcan Cotachi, northern Ecuador. Arctic, Antarctic, and Alpine Research 25:165–172
Young K. R., B. León. 1995. Connectivity, social actors, and conservation policies in the central Andes: The case of Peru’s montane forests. In S. P. Churchill, H. Balslev, E. Forero, J. L. Luteyn (eds.), Neotropical Montane Forest Biodiversity and Conservation Symposium. New York Botanical Garden, New York. Pages 653–661
Young, K. R., and B. León. 1998. Peru’s humid eastern montane forests: An overview of their physical settings, biological diversity, human use and settlement, and conservation needs. DIVA, Technical Report No. 5:1–97. http://www.diva.dmu.dk/1_viden/2_miljoe-tilstand/3_natur/diva/diva_rep.htm
Young K. R., B. León. 2000. Biodiversity conservation in Peru’s eastern montane forests. Mountain Research and Development 20:208–211.
Young K. R., W. B. Church, M. Leo, P. F. Moore. 1994 Threats to Rio Abiseo National Park, Northern Peru. Ambio 23:312–314
Young K. R., B. León, A. Cano. 1997. Peruvian Puna. In S. D. Davis, V. H. Heywood, O. Herrera-MacBryde, J. Villa-Lobos, A. C. Hamilton (eds.), Centres of plant diversity: A guide and strategy for their conservation. WWW and IUCN, Cambridge, UK. Pages 470–476
Zimmerer K. S., R. E. Galt, M. V. Buck. 2004. Globalization and multi-spatial trends in the coverage of protected-area conservation (1980–2000). Ambio 33:520–529
Zomer, R., and S. L. Ustin. 1999. Land Cover Change Along Tropical and Subtropical Riparian Corridors Within the Makalu Barun National Parkland Conservation Area, Nepal. University of California, Davis, Davis
Acknowledgments
Funding for this research was graciously provided by grants from the University of Texas Study Abroad Office, the Tinker Foundation, and the University of Texas Environmental Science Institute. We are grateful to Esteban Antonio Alibriceño, Carolina Casaretto, Peric Franco Laisa, Mariella Leo, Blanca León, Orlando Ramirez, and Rogelio Cueva Salirrosas for their assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kintz, D.B., Young, K.R. & Crews-Meyer, K.A. Implications of Land Use/Land Cover Change in the Buffer Zone of a National Park in the Tropical Andes. Environmental Management 38, 238–252 (2006). https://doi.org/10.1007/s00267-005-0147-9
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-005-0147-9