
Abstract
We propose a biodiversity credit system for trading endangered species habitat designed to minimize and reverse the negative effects of habitat loss and fragmentation, the leading cause of species endangerment in the United States. Given the increasing demand for land, approaches that explicitly balance economic goals against conservation goals are required. The Endangered Species Act balances these conflicts based on the cost to replace habitat. Conservation banking is a means to manage this balance, and we argue for its use to mitigate the effects of habitat fragmentation. Mitigating the effects of land development on biodiversity requires decisions that recognize regional ecological effects resulting from local economic decisions. We propose Landscape Equivalency Analysis (LEA), a landscape-scale approach similar to HEA, as an accounting system to calculate conservation banking credits so that habitat trades do not exacerbate regional ecological effects of local decisions. Credits purchased by public agencies or NGOs for purposes other than mitigating a take create a net investment in natural capital leading to habitat defragmentation. Credits calculated by LEA use metapopulation genetic theory to estimate sustainability criteria against which all trades are judged. The approach is rooted in well-accepted ecological, evolutionary, and economic theory, which helps compensate for the degree of uncertainty regarding the effects of habitat loss and fragmentation on endangered species. LEA requires application of greater scientific rigor than typically applied to endangered species management on private lands but provides an objective, conceptually sound basis for achieving the often conflicting goals of economic efficiency and long-term ecological sustainability.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Habitat loss and fragmentation are the leading causes of species endangerment in the United States (Wilcox and Murphy 1985, Noss and others 1997, Wilcove and others 1998). Economic goals of the expanding human population serve as the primary driver of land development (Czech and others 2000, Liu and others 2003). Private landowners have the potential to contribute significantly to biodiversity conservation or loss. Roughly 80% of endangered species occur on private lands, and 50% of these rely exclusively on privately owned habitat for survival (Noss and others 1997, Wilcove and others 1998). The increase in per capita demand for land (commonly referred to as sprawl) has been shown to be a better predictor of biodiversity loss than the rate of human population growth (Liu and others 2003). Rates of land conversion from habitat to development increase with economic value of land.
Economic incentives have been promoted as a mechanism to mitigate habitat loss on private land (Kennedy and others 1996, Shogren and others 1999). Conservation easements (Section 170(h) of the Internal Revenue Code) and conservation banking (United States Fish and Wildlife Service (USFWS) 2003) represent two approaches allowing private landowners to partially offset the cost of protecting habitat. The financial benefits of developing habitat are still often greater than cost savings provided by these policies, and Section 10 Incidental Take Permit applications are increasing (Harding and others 2001).
Mitigating the effects of habitat loss and fragmentation is challenging, because it requires accounting for the regional ecological effects resulting from local economic decisions (Fahrig and Merriam 1994, Dreschsler and Wissel 1998, Cox and Engstrom 2001). Many endangered species persist through regional exchange of individuals and genes between local populations or subpopulations (Homes and Semmens 2004). Some exist as metapopulations, or a group of subpopulations sharing immigrants at a sufficiently low rate, permitting the exchange of genes while preventing spatially correlated demographic cycles (Hanski and Gaggiotti 2004). To prevent local economic decisions from reducing metapopulation persistence, Section 10 mitigation requirements should specify that local economic decisions should not change subpopulation growth rates and migration rates estimated at the regional (or metapopulation) scale. However, current mitigation planning often ignores regional ecological effects of local economic decisions owing to a lack of resources and data (Harding and others 2001, Smallwood and others 1999).
Economic incentives that account for the regional ecological effects of local decisions may minimize and reverse the effects of habitat loss and fragmentation and may even provide justification for increased data collection and analysis. Metapopulations provide ecological services, and changes in metapopulation services are externalities of a local economic decision. We propose basing local economic incentives for habitat protection on changes in these externalities estimated at the regional scale, which provides a conservative approach to mitigating the effects of habitat loss and fragmentation on endangered species. Costs of habitat protection will differ among landowners, and their land will also differ in its contribution to ecological service flows that derive from metapopulations utilizing their land. Habitat trades based on both economic and ecological considerations are more likely to adequately balance conflicts between economic efficiency (often measured financially) and environmental sustainability (often estimated biologically). Such incentives would require linking ecological, evolutionary, and economic theory within existing policy to scale incentives appropriately.
In this article, we present a method that connects scientific theory with environmental policy to allow private landowners to profit from habitat protection while directing development (sprawl) around critical habitats within the landscape. By integrating the legal mechanism of the Endangered Species Act (ESA) with ecological, evolutionary, and economic theory, the influence of sprawl on biodiversity can be incorporated into the market value of land. We propose Landscape Equivalency Analysis (LEA), a derivation of Habitat Equivalency Analysis (National Oceanic and Atmospheric Administration (NOAA) 1999), as a method to make tradeoffs between regional conservation goals and local economic decisions. This article first outlines the theoretical basis for examining regional externalities resulting from land conversion at local scales. Then, the conservation value of a patch is defined in a manner congruent with the goals of the ESA and metapopulation theory. Finally, we synthesize theory and policy within LEA, outlining spatially explicit credits for habitat trades intended to minimize and reverse the effects of habitat loss and fragmentation.
Externalities from Land Conversion
Ecological functions can be treated as goods and services when a direct or indirect benefit to humans can be demonstrated (deGroot and others 2002). The direct human benefits of protecting endangered species include use value (e.g., seeing the species), option value (e.g., possibility that genetic variance provided by the species may contribute to medical or agricultural advances), existence value (i.e., knowing the species exists), and bequest value (i.e., knowing the species will be present for future generations) (Loomis and White 1996). Genetic variance of an endangered metapopulation also provides indirect benefits for humans. Adaptive genetic variance is required for population persistence (Fisher 1930). Neutral genetic variance is useful for determining how habitat loss or restoration affects gene flow, genetic drift, and inbreeding (Hedrick 2001). Thus, endangered species habitat is a form of natural capital, defined as a stock of resources providing useful services (deGroot and others 2000). In this analysis, we assume that the ecological services that provide these benefits are abundance and genetic variance (deGroot and others 2002).
Although total social costs of endangered species protection often fall well below total social benefits (Loomis and White 1996), the actual cost of endangered species protection often falls on relatively few households. An economic opportunity cost is incurred by private landowners based on the foregone revenue from not developing due to the presence of an endangered species (Shogren and others 1999). We define this as an economic opportunity cost due to habitat protection, OC-P. A private landowner’s use of an Incidental Take Permit (Section 10, USFWS 1988) will be based on the size of OC-P compared to costs to meet mitigation requirements preventing take and jeopardy, defined by the ESA as harming, harassing, or killing individuals and decreasing the likelihood of species survival. The ESA effectively assigns an infinite economic value to endangered species habitat as no otherwise lawful activity justifies causing a take or increasing jeopardy (Brown and Shogren 1998). In this way, the ESA theoretically prevents the loss of abundance services from endangered species habitat.
Externalities result when we are unable to protect and restore habitat at a local scale in a manner that prevents the loss of abundance and genetic variance at a larger scale. In endangered metapopulations, externalities may result from removal or addition of habitat within the landscape or changes in land use among habitat patches. Strategic protection of habitat is necessitated when migration rates among subpopulations are affected by distance or land use (e.g., roads or residential development). The contribution of a habitat patch to metapopulation service flows is determined not only by the size and vegetative composition of the patch, but also by the location of that patch relative to other patches, roads, and residential development (i.e., its spatial context). Instead of basing restoration decisions on the cheapest land to restore, metapopulation sustainability may benefit from restoration or protection of land deemed valuable for traditional economic development (i.e., high OC-P).
Defining Patch Conservation Value Inclusive of Landscape Spatial Structure
We believe that integrating population genetic theory with demographic observations would be an effective approach for defining patch conservation value. Under the ESA, habitat conservation value is often defined as its ability to maintain or decrease the probability of population extinction (Montgomery and others 1994, National Research Council (NRC) 1995). Take is typically estimated by changes in abundance, but adverse changes to habitat have also been interpreted as a take (Dwyer and others 1995). Thus, for metapopulations, the conservation value of a patch should reflect its contribution to sustainability measured at the regional scale.
The goal of the ESA is protection of enough habitat to achieve sustainable populations justifying delisting (USFWS 1988). Many believe the goal of conservation should be to protect land such that evolutionary processes are maintained (i.e., protect functional landscapes) (Frankel 1974, Meffe 1996, Moritz 2002). From a metapopulation perspective, a functional landscape would be the allocation of habitat providing rates of subpopulation growth and migration rates similar to those observed prior to habitat loss and fragmentation (Meffe 1996). As habitat patches are reduced in size and/or become isolated, the probability that deleterious recessive mutations will be expressed due to mating among related individuals, or inbreeding depression, will increase (Higgins and Lynch 2001). Conversely, changes in habitat spatial structure that increase migration rates may cause the disruption of co-adapted gene complexes, or outbreeding depression (Dudash and Fenster 2000).
Given existing uncertainties of landscape-scale management, we propose that recovery goals should incorporate genetic criteria to help define spatial allocation of habitat most likely to support sustainable metapopulations. Studies only examining abundance have not been able to resolve the relative importance of the loss of total habitat (amount) versus the change in habitat connectivity (pattern) in driving the decline or recovery of a metapopulation (Wiegand and others 1999, Fahrig 2001, Flather and Bevers 2002). Based on existing empirical evidence, the effect of changing habitat area versus connectivity on extinction risk will depend on both landscape- and species-specific variables (Debinski and Holt 2000, MacNally and others 2000).
Metapopulations must achieve a balance between growth within each subpopulation and migration between subpopulations to prevent inbreeding and outbreeding depression, and to maintain genetic variance needed for adaptation through natural selection at both individual and group levels (Harrison and Hastings 1996, Mills and Allendorf 1996). Field studies have demonstrated that mammalian metapopulations have evolved behaviors to simultaneously prevent loss of genetic variance within while maintaining genetic variance among subpopulations (Dobson and others 1997, Storz 1999, Coltman and others 2003). In a disturbed landscape, the ability of a metapopulation to balance genetic variance within and among subpopulations is likely impaired due to loss of habitat area and connectivity. Changes in landscape spatial structure have been shown to affect how genetic variance is partitioned over space (Hale and others 2001, Mech and Hallet 2001) and result in inbreeding depression (Bouzat and others 1998, Saccheri and others 1998).
Moritz (2002) stipulates that protecting the environmental context that produced existing patterns of biodiversity is the best way to maintain evolutionary processes. We can define a recovery objective as the allocation of habitat yielding the spatial apportionment of genetic variance (e.g., as would be determined using neutral genetic markers) observed prior to habitat loss and fragmentation (Meffe 1996). This recovery objective meets the definition of a functional landscape for a metapopulation given above. Given existing loss of habitat, this goal will often be unachievable, but it does serve as an objective criterion for defragmenting endangered species habitat to protect evolutionary processes at the landscape scale.
If the natural history of the organism is well known, demographic–behavioral models (Sugg and others 1996, Lacy 2000) could be used to reconstruct the spatial distribution of neutral genetic variance prior to habitat loss and fragmentation (i.e., baseline levels). This would save the expense of conducting genetic analysis. However, baseline levels of neutral genetic variance within and among subpopulations may also be estimated by integrating genetic analysis of museum specimens and extant conspecifics (Bouzat 2001, Matocoq and Villablanca 2001) with demographic–behavioral models.
The degree to which variance in neutral genetic markers as measured using molecular or biochemical markers and variance in quantitative genetic traits of adaptive significance are positively correlated is unresolved (Hedrick 2001, Reed and Frankham 2001). Positive correlations between levels of neutral and adaptive genetic variance are expected to be greater when the effects of drift exceed natural selection as the dominant evolutionary force, as is expected in populations of small size (Reed and Frankham 2001); a common problem for endangered species. Although estimates of neutral variance are available for some endangered species, estimates of adaptive genetic variance for endangered populations are often unavailable and difficult to acquire (Neel and Cummings 2003). Subpopulation growth rates, an adaptive trait (Fisher 1930, Wright 1940), can be estimated by tracking changes in abundance and will be useful for estimating the correlation between neutral and adaptive variance.
We propose that comparing observed or predicted levels of abundance and neutral genetic variance within and among subpopulations to baseline levels will allow a more thorough assessment of the tradeoffs between habitat area and connectivity than abundance alone. We thus define conservation value of habitat patch as its contribution to the maintenance of three services: (1) abundance and genetic variance (2) within and (3) among subpopulations. By incorporating baseline estimates of neutral variance both within and among subpopulations into the definition of conservation value, the spatial allocation of habitat that permitted adaptive evolution now serves as an explicit goal. This approach may replace general “rules of thumb” (e.g., one migrant per generation; Mills and Allendorf 1996) used to prescribe adequate levels of migration for endangered metapopulations. The definition of conservation value may be viewed as a species-specific and spatially explicit version of Karr and Dudley’s (1981) definition of biotic integrity. In the context of metapopulation management, a balanced, integrated, adaptive assemblage of subpopulations having the functional organization comparable to that of a natural landscape would have a high level of biotic integrity.
In summary, the conservation value of a habitat patch derives from its incremental (marginal) contribution to metapopulation sustainability. Recovery goals for the metapopulation can be translated into species-specific sustainable service flows of abundance and genetic variance. A patch’s conservation value would then equal its marginal contribution toward meeting the recovery goal, or, in the case of mitigation, the marginal decline in service flows that would result if the patch were removed (e.g., Petit and others 1998). The latter is a negative externality at the regional scale, and its magnitude provides an estimate of the ecological opportunity cost resulting from a change in landscape structure. We define this as the opportunity cost due to habitat disturbance, OC-D. Using the three estimates of metapopulation services, the OC-D represents lost opportunities for population growth and adaptive evolution.
Resource-Based Compensation
A common way to balance economic activities with conservation goals is through “service-to-service” compensation (NOAA 1999). When natural capital is injured (e.g., wetland impacted by oil spill), ecological restoration or enhancement can increase ecological service flows in a manner to equate an individual’s well-being before habitat destruction with their well-being after habitat destruction (Mazzotta and others 1994, Jones and Pease 1997). This is called resource-based compensation and is used to plan ecological mitigation to prevent the loss of social welfare. When environmental regulations stipulate in-kind replacement of ecological resources, as in the ESA, compensation must be made using the same type of resource and services that were lost (Mazzotta and others 1994).
Habitat Equivalency Analysis (HEA) is the most widely used service-to-service approach (NOAA 1999, Penn and Tomasi 2002, Strange and others 2002). HEA is a “scaling methodology” that equates losses in services due to destruction of an ecological resource in one location (injury) to gains in said services provided by an ecological resource at another location (compensatory restoration). This permits comparison of different ecological resources based on the level of service flows they provide (King 1997). The social time preference for capital assets is incorporated into HEA by discounting ecological service flows over time (Mazzotta and others 1994, NOAA 1999). The rate of discounting reflects a society’s willingness to substitute future “consumption” for present “consumption” of the ecological service (NOAA 1999).
Managing Externalities with Cap and Trade Policies
Creating a market for the exchange of positive and negative externalities at local scales is one way to prevent negative externalities at a regional scale. Marketable permit systems, or “cap and trade,” must first set a limit on the amount of externalities allowed. A regulated entity with high compliance costs of meeting the cap can purchase permits from those with low compliance costs. The increased flexibility reduces the economic costs of the meeting the cap, and the limited number of permits assures that the cap is met. In essence, a market is created so that some of the external (social) costs are incorporated into the price of the good or service that created the externality (e.g., Air Pollution Trading; Tietenberg 2004).
The ESA can be thought of as imposing a cap on further loss of abundance, as represented by the take and jeopardy standards. Conservation banking provides a system similar to a cap and trade system, wherein the purchase of a credit from the bank represents trading of access rights to endangered species habitat (USFWS 2003). Issuance of an Incidental Take Permit requires that the take be mitigated to the greatest extent practicable and that no appreciable reduction in the likelihood of species survival results (Stanford Environmental Law Society 2001). Recognizing the importance of metapopulation processes for many endangered species, Federal guidance stipulates locating a conservation bank within the landscape to minimize the effects of habitat loss and fragmentation (USFWS 2003). Also, it is critical that an accounting system be developed to ensure that credits are not oversold by a bank (i.e., creating a take or otherwise increasing risk of extinction). Since all “trades” under conservation banking are individually evaluated and subject to regulator conditions, these are not pure market systems (Shabman and Scodari 2004), but this pseudo-market system can enhance flexibility and lower compliance costs while ensuring that trades do not decrease regional service flows. We propose resource-based compensation as the basis for a landscape-scale tradable permit system.
Landscape Equivalency Analysis
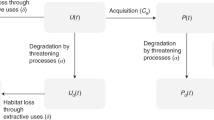
HEA does not include any ecological interaction between site of injury and site of compensatory restoration or between these sites and adjacent patches. Therefore, it is not suitable for managing metapopulations. By applying a landscape perspective to resource-based compensation, we provide a method for making tradeoffs between changes in patch conservation value, estimated as changes in OC-D at the regional scale, and OC-P at the local scale, as described in Figure 1. Applying the principles of landscape ecology to HEA will capture externalities that result from changes in habitat spatial structure. An equitable habitat trade is one that prevents OC-D from being incurred at the regional scale when economic development is pursued. We call this new formulation of HEA Landscape Equivalency Analysis (LEA).

Flow chart of proposed integrative decision analysis to mitigate the negative effects of habitat loss and fragmentation at the regional scale due to economic decisions at the local scale. Landscape Equivalency Analysis (LEA) estimates credits as discounted Landscape Service Years (dLSYs) for three metapopulation services including abundance (N), average expected heterozygosity (HS), and average genetic divergence (DST).
LEA treats the landscape as a single unit of ecological resource providing a set of services. A goal of LEA is to identify landscape configurations that provide equivalent levels of services despite changes in landscape structure that result from losing a patch or changing matrix quality. As with HEA, calculation of equivalency is based on expected changes in services over time, appropriately discounted. HEA allows adjustment for differences in efficiency (differences in service levels) across two sites by adjusting the total acreage (size of the resource) (NOAA 1999). The level of service is summarized per unit of resource (e.g., sediment retention per wetland acre), and the quality of the two sites is compared based on discounted Service Acre Years (dSAYs). dSAYs are a time-integrated estimate of resource quality based on area-weighted service flows emanating from the resource.
Landscape components must be classified so that their role in metapopulation growth and migration can be estimated (McIntyre and Hobbs 1999). Habitat within the landscape can be defined as the “resources and conditions present in an area that produce occupancy – including survival and reproduction – by a given organism” (Hall and others 1997, p. 175). Habitat patches are distinguished by greater habitat quality than surrounding areas. Areas outside of the habitat patch that allow low occupancy rates (lower habitat quality) are classified as the matrix and may be differentially permeable to migration among subpopulations. For our purposes, a landscape is defined as the patches and matrix that interact and contribute to the same set of landscape services that we wish to manage.
When the resource is a landscape, it cannot be merely assumed that differences in levels of service over time can be compensated by adjusting the total area of patches that contribute to the service. The level of landscape services will also depend on the spatial associations of patches (i.e., connectivity), and the relationship between connectivity and service flows is not necessarily monotonically increasing. No simple spatial variable (e.g., area or connectivity) will sufficiently predict changes in landscape services (Crow 2002). Therefore, LEA will use discounted Landscape Service Years (dLSYs), in which the quantification of landscape services implicitly includes the spatial aspects (area and location) of the action being considered.
The conservation value of tradeoffs between habitat amount and connectivity can be assessed by changes in dLSYs estimated at the landscape (regional) scale. LEA estimates marginal changes in externalities at the metapopulation scale based on the marginal decision to destroy or restore another patch or change matrix quality within the landscape. dLSYs is a time-integrated estimate of the externality caused by marginal changes in landscape spatial structure. LEA can also be used to estimate positive externalities resulting from habitat restoration or enhancement that cause service flows to move closer to sustainability goals.
Externalities at the regional scale due to local decisions can be estimated with spatially explicit population models (SEPMs). SEPMs describe the interaction among landscape structure and metapopulation processes (Turner and others 1995) and have been used to predict the effects of landscape management on endangered metapopulations (Liu and others 1995, Letcher and others 1998). SEPMs have also been used to predict changes in genetic variance within and among subpopulations (Lacy and Lindenmayer 1995). For example, VORTEX is an individual-based model that tracks the movement and reproductive success of each genotype (or individual) in a metapopulation (Lacy 2000). Making predictions at the subpopulation (patch) level with a SEPM is challenging due to the number of parameters required to link demography (subpopulation growth rate) and behaviors (rates and patterns of migration) to habitat quality and structure in fragmented landscapes (Ruckelshaus and others 1997, South 1999). Verifying and updating models using both demographic and genetic observations has been suggested as one approach to reduce uncertainty (Lindenmayer and Lacy 2002).
LEA provides a mechanism for integrating demographic and genetic data for decision analysis. Including genetic variance as an ecological service flow in resource-based compensation will increase our ability to make tradeoffs between local economic decisions and regional ecological effects (Figure 1).
Quantifying Genetic Variance for LEA
Species and populations within species may differ from each other in their baseline level of genetic variance (Matocq and Villablanca 2001). Genetic markers (e.g., allozymes, mitochondrial haplotypes, or microsatellites) provide estimates of whether neutral genetic variance within and among subpopulations have departed from baseline due to changes in the spatial structure of habitat (Hedrick 2001). Nei (1973) describes how data from genetic markers can be used to estimate the apportionment of neutral genetic variance at different spatial scales. When two levels of spatial organization are present (i.e., at subpopulation and metapopulation levels), Nei’s theory can be summarized as:
H T equals the total genetic diversity in the metapopulation and represents the probability that any two alleles chosen at random, one from each of two individuals, are independent (Nei 1973). The average expected genetic diversity within a subpopulation can be estimated as the average frequency of heterozygotes in a subpopulation (H S ) under Hardy-Weinberg Equilibrium. The remaining genetic variance in a metapopulation is due to divergence in allele frequencies among subpopulations, which is estimated using a measure of average minimum genetic distance (D ST ). Levels of average expected heterozygosity (H S ), the average genetic divergence (D ST ), and abundance (N) will serve as estimates of metapopulation services, which will be estimated at the regional scale.
Applying LEA to an Endangered Metapopulation
At each point in time when a decision is made regarding habitat restoration or habitat loss, an investment or withdrawal of natural capital results, changing the rate of appreciation or depreciation. A withdrawal that drives services below current levels violates ESA, representing a take and may increase the risk of extinction. Sufficient investment above current levels, without withdrawals, eventually leads to species recovery. LEA facilitates a tradable credit market that is driven by private landowners’ interest in maximizing land values but is constrained by the Federal goal of achieving sustainable populations (by ensuring credit trades do not violate ESA take and jeopardy standards). The uniqueness of this approach lies in the ability of LEA to incorporate the unequal contribution of habitat patches and landscape matrix to multiple metapopulation service flows.
In the remainder of this section, we illustrate the LEA approach using several simple qualitative descriptions of the effects of spatial structure on metapopulation services. A fully quantitative approach would require development and description of a SEPM, which is beyond the scope of this paper. The critical output from a quantitative analysis would be expected levels of abundance (N), average expected heterozygosity (H S ), and average genetic divergence (D ST ) over time under different landscape configurations. To illustrate LEA, Figure 2 presents three hypothetical landscape structures. For the illustration, several assumptions are made that need not hold for general applications of LEA. Habitat patches in the landscape differ in the amount of habitat destruction they experienced and in ownership (patches with a G or P represent government or private land, respectively). Even though patches differ in level of connectivity and habitat area, the illustration assumes equal habitat quality across patches for all extant subpopulations. We make the simplifying assumption that the matrix is homogeneous, and dispersal is only limited by the distance between extant patches. In a real landscape, habitat and matrix quality will vary. We also assume that movement of individuals across proximate subpopulations is constant over time. We do not consider that metapopulations often require empty but suitable habitat patches to colonize when local extinction occurs elsewhere (Thomas and Hanski 1997).

Change in spatial structure of hypothetical landscape over time with multiple landowners. Patches labeled “P” are each owned by different private parties and those labeled “G” are owned by the Federal government. Open areas represent endangered species habitat that has been lost to economic development. Filled areas represent habitat supporting the endangered population. Length of double-sided arrows equals the species maximum dispersal distance, indicating connectivity. The matrix is homogeneous, and dispersal is only limited by the distance between patches. (A) Shaded and nonshaded areas combined represent the baseline habitat distribution that would provide a sustainable metapopulation (b-curve in Figure 3). Shaded areas represent the current remnant subpopulations (j-curve in Figure 3). (B) Conservation bank is added to metapopulation (m-curve) changing landscape spatial structure. (C) Three possible choices for economic growth leading to an endangered species take are displayed (withdrawal of credits: w1-, w2-, and w3-curves in Figure 3). Open arrows represent connectivity lost because of economic development.
Figure 2A displays a landscape structure meeting the sustainability (recovery) goal, in which the combined shaded and nonshaded areas represent the historic geographic range of the species. Expected service levels from this landscape are referred to as the b-trajectory (baseline) in Figure 3. Here we assume that the recovery goal is to restore the historic geographic range of the species. Figure 2A also displays habitat remnants (shaded areas) that provide the status quo level of services (j-trajectory in Figure 3). The status quo services will reflect service levels below which constitute a take. In general, as populations become subdivided, gene flow is restricted and genetic drift increases, causing a loss of genetic variance within a subpopulation but an increase in genetic divergence among populations (Whitlock 2004). Given the habitat loss and fragmentation indicated in Figure 2A, we can expect a large reduction in abundance within the metapopulation, a decrease in average heterozygosity within subpopulations, and an increase in average genetic divergence among subpopulations (Figure 3).

Trajectories of metapopulation services used to calculate Landscape Equivalency Analysis (LEA). Figure 2 reports the landscape spatial structure hypothesized to give each service flow trajectory. The expected service flow trajectories are represented by lowercase letters. When time = M, a conservation bank is added to the metapopulation. When time = W, services flows resulting from one of three possible withdrawals of credits from the bank are projected. Each time the landscape structure is changed (i.e., time = M and time = W), service flows are estimated assuming no changed occurred, as indicated by the dotted lines. These estimates are required so that LEA can calculate the change in appreciation or depreciation of service flows from natural capital.
Figure 2B indicates that a conservation bank has been added to the metapopulation at a later time (M in Figure 3). Assuming a SEPM can be constructed, the interaction between a species’ natural history and landscape structure can be modeled. Therefore, the SEPM can be used to estimate metapopulation services resulting from the restoration of different sites, given landscape- and species-specific conditions, in order to find the best location for the bank. The bank location in the hypothetical landscape was chosen because it has a high probability of being colonized by individuals from adjacent habitat (P3) and sharing migrants with the smallest habitat remnants (i.e., patches with highest probabilities of extinction, G2, P2, and P4). Figure 3 illustrates that the placement of the bank should move services closer to sustainability goals, helping to reverse the negative effects of habitat loss and fragmentation (m-trajectory).
The number of credits available in a bank is conditional upon the ability of the mitigation plan to increase conservation values (USFWS 2003), estimated in our analysis as increased service flows. Sale of credits represents a decrease in service flows due to the decision of another private landowner to develop a habitat patch. Figure 2C displays three possible directions of economic growth, each resulting in a take. The SEPM is used to estimate the change in service flows due to each possible direction of growth (the w-trajectories in Figure 3). Expected changes in genetic variance will depend on interactions between organismal and landscape history. Furthermore, the magnitude of changes in average expected heterozygosity (H S ) and average genetic divergence (D ST ) should be especially sensitive to changes in landscape structure.
Direction 1 results in losing a habitat patch that recently connected organisms inhabiting private and public land (i.e., a stepping stone is lost, P2) due to conservation banking. The subpopulation in G2, which is publicly owned, would then suffer from increased genetic drift and inbreeding due to the loss of gene flow. In this example, the service most affected by this trade would be average genetic divergence among subpopulations, because the increased connectivity provided by the bank is lost. Also, metapopulation abundance and average expected heterozygosity would be reduced (Figure 3, trajectory w1).
Direction 2 results in reducing the size of a historically large subpopulation (P1). Also, gene flow between the two largest subpopulations is lost. The services most likely affected by this trade are abundance and average expected heterozygosity within each subpopulation. This larger subpopulation contributed disproportionately to abundance and average expected heterozygosity measured at the regional scale, but contributed little to average genetic divergence, owing to its larger subpopulation size and comparatively high exchange of migrants with another large subpopulation. Because the subpopulation experiencing the take has been historically outbred, it may be more susceptible to inbreeding depression due to harboring more deleterious recessive mutations than smaller, peripheral subpopulations (Frankham and others 2001). Therefore, there is a greater probability that inbreeding depression will threaten population persistence in both P1 and G1, which is why the development scenario is projected to have the biggest impact on metapopulation abundance (Figure 3). Average genetic divergence will also increase over time, but more slowly, compared to the smaller peripheral patches.
Direction 3 results in losing the most isolated patch (P4). Subpopulation P4 is likely the most genetically distinct, and its loss has a disproportionate effect on average genetic divergence at the metapopulation scale, bringing the subpopulation closer to baseline (Figure 3). Although Nei’s (1973) estimates of genetic variance are weighted by population size, loss of the isolated patch may slightly increase the average expected heterozygosity at the metapopulation scale. The reduction in metapopulation abundance is expected to be relatively small compared to the other possible takes (w1 and w2). Even though the area of habitat lost in directions 1 and 3 are very similar, a greater decline in abundance is expected under direction 1 because the extinction risk for the subpopulation G2 is increased by the loss of the stepping stone (P2). This simple example described how each metapopulation service differs in sensitivity to changes in habitat area and connectivity.
If the loss of any habitat results in loss of alleles only found in that subpopulation (i.e., private alleles), Nei’s estimates of total genetic diversity may be inadequate for decision making, and measures of allelic richness should be considered (Petit and others 1998, Neel and Cummings 2003). In this case, translocation of organisms may also be considered (Moritz 1999). These topics are beyond the scope of this article, however.
Calculating Biodiversity Credits using LEA
The number of credits the private landowner must purchase to offset externalities that result from habitat alterations can be calculated by comparing changes in service trajectories. The number of credits required to offset the local and regional loss of abundance due to losing a habitat patch can be calculated as discounted Landscape Service Years – Abundance (dLSYN):
where loss of service flows due to habitat loss for the endangered metapopulation begins at year W, r is the social discount rate, b N t is the expected abundance for the metapopulation at year t (representing the recovery goal), m N t is the expected abundance at year t inclusive of adding or enhancing the bank patch that will sell credits, and w N t is the expected abundance at year t, reflecting anticipated loss of habitat or connectivity.
dLSYN represents the marginal conservation value of the lost patch, given its spatial context within the metapopulation relative to a recovery goal (i.e., the negative externality in abundance that results from losing that patch to development). The credit represents the fraction of problem solved by mitigation minus the fraction of the solution lost due to take elsewhere in the landscape. Because the public has a positive time preference for services from capital assets, discounting modifies the number of credits associated with the change in abundance, as in HEA. In other words, the more slowly the withdrawal decreases population size, the fewer credits must be purchased.
Calculating credits associated with changes in genetic variance is more complex. The management goal is to approximate the distribution of habitat in which the organism evolved (baseline landscape) (Meffe 1996, Moritz 2002). Greater genetic diversity within a subpopulation or greater genetic divergence among subpopulations is not always better for sustainability (Bouzat 2001). As estimates of genetic variance within and among subpopulations move farther away from baseline levels due to losing a patch or connectivity, the larger the credit purchased from the bank will have to be. The credit representing the magnitude of externality in genetic services due to losing a patch elsewhere can be calculated as discounted Landscape Service Years – Genetic Variance (dLSYG):
where G is the genetic variance component estimated (H S or D ST ), b G t is the expected level of genetic variance at year t representing the recovery goal or baseline levels, m G t is the expected level of genetic variance at year t inclusive of adding or enhancing the bank patch that will sell credits, and w G t is the expected level of genetic variance at year t reflecting anticipated loss of habitat or connectivity.
For any positive discount rate, the more slowly a landscape change moves the population away from baseline, the fewer credits per change in service level are incurred. Conversely, habitat changes resulting in a large departure from baseline in the immediate future will be charged many credits per change in service level.
Assuming that neutral variance serves as a surrogate for adaptive variance in a small population, when H S and D ST are close to baseline the probability of inbreeding and outbreeding depression will be reduced, while opportunities for natural selection at individual and group levels are protected. Conversely, if we assume no correlation between neutral and adaptive variance, when H S and D ST are close to baseline, subpopulation growth rates and migration rates (i.e., metapopulation processes) observed prior to habitat loss and fragmentation have been closely approximated.
It is possible for m G t , j G t , or w G t to oscillate about b G t and each other over time. The amplitude in the oscillations and average distance from b G t will be reflected in the credit estimate using the equation above. If the loss of a patch (wG) produces large oscillations and pushes genetic variance farther away from bG than previously observed under mitigation-level scenario (mG), dLSYG will be large.
Because of the denominator, the smaller the baseline level of genetic variance (bG), the larger dLSYG would be per change in service levels. Therefore, sensitivity of the dLSYG measure increases as the baseline genetic variance moves closer to zero. If the baseline average expected heterozygosity within a subpopulation were low (bHs → 0), genetic drift caused by habitat loss and fragmentation may quickly drive H S to zero. Conversely, if mitigation increased H S well above baseline because of immigration, disruption of locally adaptive gene complexes may result in outbreeding depression (Dudash and Fenster 2000). If the baseline level of genetic divergence were low (bDst → 0), the metapopulation historically experienced high rates of gene flow among patches. Therefore, dLSYDst would be more sensitive to changes in habitat connectivity.
Recommendations for Basic Conservation Banking Scenario
In the simple example described in Figures 2 and 3, the size of credits can be estimated by examining the graphs regardless of the discount rate, because trajectories of service levels do not cross. Economic development in direction 3 would require the smallest number of credits to be purchased from the bank for each metapopulation service. Direction 2 results in the largest externalities as estimated by abundance and average expected heterozygosity, but a relatively smaller externality as estimated by changes in genetic divergence when compared to direction 1. Direction 1 results in an intermediate externality for abundance and average expected heterozygosity, but the largest externality for genetic divergence credits.
Results from the hypothetical example are obvious because only the influence of the organism’s maximum dispersal distance and previous landscape structure are considered. Incorporating more details on landscape structure and an organism’s natural history may lead to a different placement of the bank and/or a different recommended direction for economic growth. However, even in our simple example, we show that by only examining patch-level changes in abundance at the site of take and bank, and ignoring metapopulation dynamics, all three directions of growth are likely to be considered equally undesirable, because the habitat area lost for each direction of growth is roughly equal. LEA incorporates information on existing service levels that will help capture previous loss of connectivity, reduction in population size, and extinction of subpopulations, occurring at different times, that would be missed if trades were based solely on demographics (e.g., Frankham 1995, Luikart and others 1998).
Trading Metapopulation Credits
A private landowner wishing to purchase an Incidental Take Permit from the conservation bank would have to buy credits for each service individually. The price of the credit represents the in-kind replacement value for each service inclusive of landscape spatial structure. Applying resource-based compensation to habitat trades within a metapopulation will require institutional arrangements that outline how the Federal government, conservation bank, and private landowners interact to achieve sustainability goals. A market for metapopulation service flows from a landscape will likely be a centralized market (i.e., a pseudo-market) in which Federal oversight is needed to ensure that sufficient scientific certainty for trading exists and that banks do not oversell credits (USFWS 2003, Shabman and others 1996). Federal oversight will be especially important when managing metapopulations because of the potential for a neighbor reducing habitat quality, subsequently reducing the marginal conservation value of adjacent patches (i.e., an unmitigated take). Trading rules enforced by regulators should promote trading of credits (i.e., allowing development) while not further endangering the population (Shabman and others 1996). Some preliminary rules are listed below.
Rule #1. Trades must not violate take and jeopardy standards. Two necessary conditions for all trades could be that no take results from the action (i.e., mean-wN > mean-jN, where jN is expected abundance under status quo conditions) or the probability of extinction under the j-trajectory (Ej) must be greater than or equal to the probability of extinction under the w-trajectory (Ew) (i.e., P[EJ] ≥ P[Ew]). Violation of these conditions (i.e., take and jeopardy, respectively) means that the trade would result in overdrawing credits (i.e., cap is exceeded). This indicates that the bank has not yet provided a sufficient increase in ecological services to make the trade. More time and/or restoration are required before trading would be allowed, or a different ‘w-action’ could be considered.
Rule #2. Trades should not produce an allocation of habitat that drives the spatial apportionment of genetic variance farther away from baseline levels. If the projected maximum absolute difference between bG (baseline levels of the genetic variance component) and wG across all time is greater than that between bG and jG (i.e., max |bG – jG| ≥ max |bG – wG|, where jG is the expected level of genetic variance in status quo landscape), then the loss of the patch may move the metapopulation farther away from baseline than was observed under status quo conditions. Changes in spatial distribution of genetic variance are not currently used to define a take. However, habitat trades that drive genetic variance farther away from baseline may exacerbate effects of habitat loss and fragmentation. Accordingly, we recommend that trades among genetic services (e.g., dLSYHs for dLSYDst) should not be allowed. Genetic variance within and among subpopulations both contribute to evolution but in different ways (i.e., natural selection at individual versus group level), and we are uncertain of the long-term effects of such tradeoffs (Moritz 1994). Under certain circumstances, for example, moving DST farther away from baseline to alleviate inbreeding depression (increase HS), these tradeoffs may be advised. LEA provides a framework for managing these long-term and short-term goals.
Rule #3. Private investment in natural capital leads to marketable credits, whereas public investment in natural capital leads to species recovery. The trading of habitat patches among private parties using LEA minimizes the effect of habitat loss and fragmentation. Private landowners are only legally responsible for not increasing the probability of extinction or otherwise causing a take relative to the status quo of the population (i.e., increasing the current rate of depreciation) (Harding and others 2001). However, purchase of credits by public agencies or NGOs for reasons other than mitigating a take represent a net investment in natural capital leading to habitat defragmentation. This would be a cost-effective approach to promote species recovery.
Rule #4. Begin discounting of service flows from the time the trade occurs. Our goal is to meet the preferences of the current generation without sacrificing the welfare of future generations. Each time a trade is made, the credit will be estimated with the current estimate of that generation’s rate of positive time preference. The welfare of future generations is protected at a minimum level by preventing trades that cause a take or increase jeopardy.
Rule #5. Monitoring data must be continually used to update the SEPM, so that conservation banking helps reduce uncertainty. Conservation banking provides a financial incentive for reducing uncertainty. Conservation banking guidance (USFWS 2003) indicates that the size of the endowment used to fund perpetual management should be proportional to the risk the banker is accepting, and the cost of maintaining that account should be incrementally offset by the sale of each credit. If uncertainty regarding an endangered species' natural history were too great, conservation banking should be cost prohibitive. Increasing OC-P would provide economic drivers for collecting data to reduce uncertainty, facilitating market entry.
SEPMs have been heavily criticized because detailed datasets are required to parameterize the models (Beissinger 2002). Beissinger (2002) observed that when large opportunity costs are associated with species protection (OC-P), analysis of management scenarios using detailed datasets often result. Examples include models for the northern spotted owl (Lamberson and others 1994), California gnatcatcher (Akcakaya and Atwood 1997), Florida scrub jay (Breininger and others 1999), red cockaded woodpecker (Letcher and others 1998), and Bachman’s sparrow (Liu and others 1995). The ever increasing economic value of real estate and human population growth (Liu and others 2003) suggest that the OC-P will increase in the future for most species. In other words, despite the uncertainties associated with SEPM, modeling should become financially feasible in the future for more species as the economic value of land increases.
Differences in mitigation costs result from the influence of landscape spatial structure on metapopulation services (i.e., patches differ in influence on OC-D) and from differences in land values as determined by traditional markets (i.e., OC-P; Ando and others 1998). The mitigation costs for externalities (price of dLSY for three metapopulation services: N, H S , D ST ) relative to the expected financial benefit of economic growth ultimately determine the spatial allocation of habitat (Figures 1 and 2). If we assume that all participants are price takers (i.e., the banker will try to maximize profit and purchaser will minimize mitigation costs) and share perfect information, the market would prevent the spatial allocation of habitat from moving farther away from the allocation of habitat that permitted adaptive evolution. Bank patches strategically located will be able to sell many credits. Patches that contribute greatly to metapopulation size and genetic variance will cause a greater withdrawal from the bank if lost to development. LEA provides an effective accounting system for conservation banking while linking regional ecological effects to local economic decisions (Figure 1). Trading habitat credits with LEA will distribute the cost to comply with the ESA more evenly among private landowners (Olson and others 1993).
Discussion
We have introduced a novel approach for calculating biodiversity credits that are sensitive to landscape spatial structure, a species’ natural history, existing service levels, and society’s rate of positive time preference (i.e., discount rate). The price of the credit represents the in-kind replacement value for three metapopulation services. Purchase of a credit includes the influence of sprawl on biodiversity at the regional scale, such that a local economic decision will only proceed if the price of the biodiversity credit is sufficiently less than the expected net economic benefit of destroying habitat (Figure 1). We have summarized information so that a private landowner’s decision reflects the tradeoff between OC-P and mitigation costs to prevent loss of conservation value measured at the regional scale (OC-D).
Whenever off-site mitigation compensates for habitat loss, management decisions change the spatial structure of the landscape (Huxel and Hastings 1999). Incorporating spatial biological processes into decision making may indicate tradeoffs between habitat area and connectivity (Lamberson and others 1994, Cox and Engstrom 2001). These tradeoffs may find habitat allocations that protect endangered species without preventing economic growth. This paper provides a conservative approach for assessing these tradeoffs while providing an economic incentive for greater data collection and analysis.
Usually, the number of credits available in a conservation bank are based on habitat suitability indices, size of habitat, and/or number of individuals observed within the patch (USFWS 2003). This approach ignores the importance of spatial structure. Calculation of conservation credits for a bank for red legged frog in California included habitat connectivity, habitat shape, and habitat location criteria (USFWS 2001). These indices are based on generalizations relating the natural history of vertebrate organisms to landscape structure (i.e., bigger habitat with less edge and more connectivity is always best). However, we lack a theory describing the effects of habitat loss and fragmentation on populations (Fahrig 2003). Furthermore, evolutionary theory and empirical data indicate that some isolation between subpopulations plays an important role in adaptive evolution (Lesica and Allendorf 1995). The effects of habitat loss and fragmentation vary based on species history and landscape history. Thus, no prior generalizable approach for calculating spatially explicit credits has been derived.
The lack of theory and presence of uncertainty have not stopped economic decisions that change the landscape spatial structure (Wilcove and others 1998, Smallwood and others 1999, Harding and others 2001). In lieu of analytical models, we propose a solution that integrates case-specific landscape simulation models with resource-based compensation. For some endangered species, use of the best available science and data (Smallwood and others 1999) entails constructing a SEPM so that demography, behavior, and genetics can be related to landscape structure. Simulation modeling may reveal critical landscape components not identified by ignoring interactions between natural history and spatial structure. LEA could be used to justify protecting and restoring these critical landscape components and directing sprawl around these areas.
The Federal government requires monitoring of conservation values in a bank, because ecosystems will not be static over time (USFWS 2003), suggesting that conservation banking may benefit from Adaptive Management. Adaptive Management is a method of incorporating the role of uncertainty in decision making by treating decisions as hypotheses regarding the system’s response to management actions (Walters 1986). Monitoring data are used to test the validity of alternative hypotheses, given existing knowledge of the system. Using monitoring data to continually improve our understanding of the system improves future decisions. LEA would benefit from SEPMs incorporating different hypotheses relating genetic variance to demographic parameters. However, the curse of dimensionality (Ludwig and Walters 2002) and difficulty of comparing replicate metapopulations (Dunning 2002) are significant barriers to applying Adaptive Management to conservation banking at the landscape scale.
Previous authors have recognized the need for a tool like LEA. Kennedy and others (1996) outlined an incentive system to permit trading of endangered species habitat. These authors recognized that demographic and genetic characteristics would be important measures of conservation value upon which the incentive system would be based, but provided no mechanistic approach for meeting this goal. The NRC emphasized the need for explicit decision-making tools that incorporate the influence of uncertainty when making tradeoffs between economic and conservation values under ESA (NRC 1995). NRC (1995) also recommended using a landscape perspective when making management decisions despite the scientific uncertainties associated with the large geographic scale of analysis. Similarly, an NRC study of wetland mitigation banking stressed the need to site banks within a landscape so that trading of credits prevents the loss of wetland function necessary for ecological sustainability (NRC 2001). Wetland scientists may find LEA useful for meeting this goal.
Conclusion
Ecological goods and services are often not given adequate consideration during decision making because we lack markets to value them. A market for trading habitat, as provided by conservation banking, could serve as a market for trading ecological services (Daily and Ellison 2003) such as abundance and genetic variance of an endangered species. Applying a landscape perspective to conservation banking also provides a means for conservationists to justify the protection of real estate that is considered valuable by the traditional market. Protected lands have historically been low in economic value, or lands provided by individuals with a strong conservation ethic (i.e., opportunistic protection). Strategic methods of habitat protection sensitive to economic behavior will be critical to the success of biodiversity conservation. Strategic decisions incorporating economic, ecological, and evolutionary components can be made by comparing development scenarios with LEA. We can now apply an evolutionary perspective to describe the biophysical implications of economic growth.
Literature Cited
Akcakaya H. R., J. L. Atwood. 1997. A habitat-based metapopulation model of the California Gnatcatcher. Conservation Biology 11:422–434
Ando A., J. Camm, S. Polasky, A. Solow. 1998. Species distributions, land values, and efficient conservation. Science 279:2126–2128
Beissinger S. R. 2002. Population viability analysis: past, present, and future. in S. R. Beissinger, D. R. McCullough (eds.), Population viability analysis. The University of Chicago Press, Chicago, Illinois, Pages 5–17
Bouzat J. L. 2001. The importance of control populations for the identification and management of genetic diversity. Genetica 110:109–115
Bouzat J. L., H. H. Cheng, H. A. Lewin, R. L. Westemeier, J. D. Brawn, K. N. Paige. 1998. Genetic evaluation of a demographic bottleneck in the greater prairie chicken. Conservation Biology 12:836–843
Breininger D. R., M. A. Burgmen, B. M. Stith. 1999. Influence of habitat quality, catastrophes, and population size on extinction risk of the Florida scrub-jay. Wildlife Society Bulletin 27:810–822
Brown Jr. G. M., J. F. Shogren. 1998. Economics of the Endangered Species Act. Journal of Economic Perspectives 12:3–20
Coltman D. W., J. G. Pilkington, J. M. Pemberton. 2003. Fine-scale genetic structure in a free-living ungulate population. Molecular Ecology 12:733–742
Cox J., R. T. Engstrom. 2001. Influence of the spatial pattern of conserved lands on the persistence of a large population of red-cockaded woodpeckers. Biological Conservation 100:137–150
Crow T. R. 2002. Putting multiple use and sustained yield into a landscape context. In: J. Liu, W. W. Taylor (eds.), Integrating landscape ecology into natural resource management. Cambridge University Press, Cambridge, Pages 349–365
Czech B., P. R. Krausman, P. K. Devers. 2000. Economic associations among causes of species endangerment in the United States. BioScience 50:593–601
Daily G. C., K. Ellison. 2003. The new economy of nature: the quest to make conservation profitable. Island Press, Washington, D.C., 260 pp
Debinski D. M., R. D. Holt. 2000. A survey and overview of habitat fragmentation experiments. Conservation Biology 14:342–355
deGroot R. S., J. van der Perk, A. Chiesura, S. Marguliew. 2000. Ecological functions and socio-economic values of critical natural capital as a measure of ecological integrity and environmental health. In: P. Crabbe, A. Holland, L. Ryszkowski, L. Westra (eds.), Implementing ecological integrity: restoring regional and global environmental and human health. Kluwer Academic Publishers, Boston, Pages 191–214
deGroot R. S., M. A. Wilson, R. M. J. Boumans. 2002. A typology for the classification, description and valuation of ecosystem functions, goods, and services. Ecological Economics 41:393–408
Dobson S. F., R. K. Chesser, J. L. Hoogland, D. W. Sugg, D. W. Foltz. 1997. Do black-tailed prairie dogs minimize inbreeding? Evolution 51:970–978
Drechsler M., C. Wissel. 1998. Trade-offs between local and regional scale management of metapopulations. Biological Conservation 83:31–41
Dwyer L. E., D. D. Murphy, P. R. Ehrlich. 1995. Property-rights case law and the challenge to the Endangered Species Act. Conservation Biology 9:725–741
Dudash M. R., C. B. Fenster. 2000. Inbreeding and outbreeding depression in fragmented populations. In: A. G. Young, G. M. Clarke (eds). Genetics, demography, and viability of fragmented populations. Cambridge University Press, New York. pp 35–53
Dunning Jr. J. B. 2002. Landscape ecology in highly managed regions: the benefits of collaboration between management and researchers. In: J. Liu, W. W. Taylor (eds). Integrating landscape ecology into natural resource management. Cambridge University Press, Cambridge. pp 334–345
Fahrig L. 2001. How much habitat is enough? Biological Conservation 100:65–74
Fahrig L. 2003. Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution, and Systematics 34:487–515
Fahrig L., G. Merriam. 1994. Conservation of fragmented populations. Conservation Biology 8:50–59
Fisher R. A. 1930. The genetical theory of natural selection. Clarendon Press, Oxford, 318pp
Flather C. H., M. Bevers. 2002. Patchy reaction-diffusion and population abundance: the relative importance of habitat amount and arrangement. American Naturalist 159:40–56
Frankel O. H. 1974. Genetic conservation: our evolutionary responsibility. Genetics 78:53–65
Frankham R. 1995. Inbreeding and extinction—a threshold effect. Conservation Biology 9:792–799
Frankham R., D. M. Gilligan, D. Morris, D. A. Briscoe. 2001. Inbreeding and extinction: effects of purging. Conservation Genetics 2:279–285
Hale M. L., P. W. W. Lurz, M. D. F. Shirley, S. Rushton, R. M. Fuller, K. Wolff. 2001. Impact of landscape management on the genetic structure of Red Squirrel populations. Science 293:2246–2248
Hall L. S., P. R. Krausman, M. L. Morrison. 1997. The habitat concept and a plea for standard terminology. Wildlife Society Bulletin 25:173–182
Hanski I., O. E. Gaggiotti. 2004. Metapopulation biology: past, present, and future. In: I. Hanski, O. E. Gaggiotti (eds). Ecology, genetics, and evolution of metapopulations. Elsevier Academic Press, San Diego, California. pp 3–22
Harding E. K., E. E. Crone, B. D. Elderd, J. M. Hoekstra, A. J. McKerrow, J. D. Perrine, J. Regetz, L. J. Rissler, A. G. Stanley, E. L. Walters, NCEAS Habitat Conservation Plan Working Group. 2001. The scientific foundations of Habitat Conservation Plans: a quantitative assessment. Conservation Biology 15:488–500
Harrison S., A. Hastings. 1996. Genetic and evolutionary consequences of metapopulation structure. Trends in Ecology and Evolution 11:180–183
Hedrick P. W. 2001. Conservation genetics: where are we now? Trends in Ecology and Evolution 16:629–636
Higgins K., M. Lynch. 2001. Metapopulation extinction caused by mutation accumulation. Proceedings of the National Academy of Sciences USA 98:2928–2933
Homes E. E., B. X. Semmens. 2004. Viability analysis for endangered metapopulations: a diffusion approximation approach. In: I. Hanski, O.E. Gaggiotti (eds.) Ecology, genetics, and evolution of metapopulations. Elsevier Academic Press, San Diego, California. pp 565–597
Huxel G. R., A. Hastings. 1999. Habitat loss, fragmentation, and restoration. Restoration Ecology 7:309–315
Jones C. A., K. A. Pease. 1997. Restoration-based compensation measures in natural resource liability statutes. Contemporary Economic Policy 15:111–122
Karr J. R., D. R. Dudley. 1981. Ecological perspective on water quality goals. Environmental Management 5:55–68
Kennedy E. T., R. Costa, W. M. Smathers Jr. 1996. New directions for red-cockaded woodpecker habitat conservation: economic incentives. Journal of Forestry 94:22–26
King, D. 1997. Comparing ecosystem services and values. U. S. Department of Commerce, NOAA, Damage Assessment and Restoration Program, Silver Spring, Maryland
Lacy R. C. 2000. Structure of the VORTEX simulation model for population viability analysis. Ecological Bulletins 48:191–203
Lacy R. C., D. B. Lindenmayer. 1995. A simulation study of the impacts of population subdivision on the mountain brushtail possum Trichosurus caninus Ogilby (Phalangeridae: Marsupialia), in south-eastern Australia. II. Loss of genetic variance within and between subpopulations. Biological Conservation 73:131–142
Lamberson R. H., B. R. Noon, C. Voss. 1994. Reserve design for territorial species—the effects of patch size and spacing on the viability of the northern spotted owl. Conservation Biology 8:185–195
Lesica P., F. W. Allendorf. 1995. When are peripheral populations valuable for conservation? Conservation Biology 9:753–760
Letcher B. H., J. A. Priddy, J. R. Walters, L. B. Crowder. 1998. An individual-based, spatially-explicit simulation model of the population dynamics of the endangered red-cockaded woodpecker, Picoides borealis. Biological Conservation 86:1–14
Lindenmayer D. B., R. C. Lacy. 2002. Small mammals, habitat patches and PVA models: a field test of model predictive ability. Biological Conservation 103:247–265
Liu J., J. B. Dunning Jr., H. R. Pulliam. 1995. Potential effects of a forest management plan on Bachman’s sparrows (Aimophila aestivalis): linking a spatially explicit model with GIS. Conservation Biology 9:62–75
Liu J., G. C. Daily, P. R. Ehrlich, G. W. Luck. 2003. Effects of household dynamics on resource consumption and biodiversity. Nature 421:530–533
Loomis J. B., D. S. White. 1996. Economic benefits of rare and endangered species: summary and meta-analysis. Ecological Economics 18:197–206
Ludwig D., C. J. Walters 2002. Fitting population viability analysis into adaptive management. In: S. R. Beissinger, D. R. McCullogh (eds). Population viability analysis. The University of Chicago Press, Chicago, Illinois. pp 511–520
Luikart G., W. B. Sherwin B. M. Steele F. W. Allendorf. 1998. Usefulness of molecular markers for detecting population bottlenecks via monitoring genetic change. Molecular Ecology 7:963–974
MacNally R., A. F. Bennett, G. Horrocks. 2000. Forecasting the impacts of habitat fragmentation. Evaluation of species-specific predictions of the impact of habitat fragmentation on birds in the box-ironbark forests of central Victoria Australia. Biological Conservation 95:7–29
Matocoq M. D., F. X. Villablanca. 2001. Low genetic diversity in an endangered species: recent or historical pattern? Biological Conservation 98:61–68
Mazzotta M. J., J. J. Opaluch, T. A. Grigalunas. 1994. Natural resource damage assessment: the role of resource restoration. Natural Resources Journal 34:153–175
McIntyre S., R. Hobbs. 1999. A framework for conceptualizing human effects on landscapes and its relevance to management and research models. Conservation Biology 13:1282–1292
Mech S. G., J. G. Hallet. 2001. Evaluating the effectiveness of corridors: a genetic approach. Conservation Biology 15:467–474
Meffe G. K. 1996. Conserving genetic diversity in natural systems. In: R. C. Szaro, D. W. Johnston (eds). Biodiversity on managed landscapes: theory and practice. Oxford University Press, New York. pp 41–57
Mills L. S., F. W. Allendorf. 1996. The one-migrant-per-generation rule in conservation and management. Conservation Biology 10:1509–1518
Montgomery C. A., G. M. Brown Jr., D. M. Adams. 1994. The marginal cost of species preservation: the northern spotted owl. Journal of Environmental Economics and Management 26:111–128
Moritz C. 1999. Conservation units and translocations: strategies for conserving evolutionary processes. Hereditas 130:217–228
Moritz C. 2002. Strategies to protect biological diversity and the evolutionary processes that sustain it. Systematic Biology 51:238–254
Mortitz C. 1994. Applications of mitochondrial DNA analysis in conservation: a critical review. Molecular Ecology 3:401–411
NOAA (National Oceanic and Atmospheric Administration). 1999. Habitat equivalency analysis: an overview. Policy and technical paper series, No. 95-1. Technical paper 99-1. Damage Assessment and Restoration Program, Damage Assessment Center. Silver Springs, Maryland
NRC (National Research Council). 1995. Science and the endangered species act. National Academy Press, Washington, D.C., 271 pp
NRC (National Research Council). 2001. Compensating for wetland losses under the Clean Water Act. National Academy Press. Washington, D.C., 322 pp
Neel M. C., M. P. Cummings. 2003. Genetic consequences of ecological reserve design guidelines: an empirical investigation. Conservation Genetics 4:427–439
Nei M. 1973. Analysis of gene diversity in subdivided populations. Proceedings of the National Academy of Science USA 70:3321–3323
Noss R. F., M. A. O’Connell D. D. Murphy. 1997. The science of conservation planning. Island Press, Washington, D.C., 263 pp
Olson T. G., D. D. Murphy, R. D. Thornton. 1993. The habitat transaction method: a proposal for creating tradable credits in endangered species habitat. In: W. E. Hudson (eds). Building economic incentives into the Endangered Species Act. Defenders of Wildlife, Washington, D.C. pp 27–36
Penn T., T. Tomasi. 2002. Calculating resource restoration for an oil discharge in Lake Barre, Louisiana, USA. Environmental Management 29:691–702
Petit R. J., A. E. Mousadik, O. Pons. 1998. Identifying populations for conservation on the basis of genetic markers. Conservation Biology 12:844–855
Reed D. H., R. Frankham. 2001. How closely correlated are molecular and quantitative measures of genetic variance? a meta-analysis. Evolution 55:1095–1103
Ruckelshaus M., C. Hartway, P. Kareiva. 1997. Assessing the data requirements of spatially explicit dispersal models. Conservation Biology 11:1298–1306
Saccheri I., M. Kuussaari, M. Kankare, P. Vikman, W. Fortelius, I. Hanski. 1998. Inbreeding and extinction in a butterfly metapopulation. Nature 392:491–494
Shabman, L. A., and P. Scodari. 2004. Past, present, and future of wetlands credit sales. Discussion paper 04–48, Resources for the Future, Washington, DC
Shabman L. A., P. Scodari, D. King 1996. Wetland mitigation banking markets. In: L. L. Marsh, D. R. Porter, D. A. Salvesen (eds). Mitigation banking: theory and practice. Island Press, Washington, D.C. pp 109–138
Shogren J. F., J. Tschirhart, T. Anderson, A.W. Ando, S. R. Beissinger, D. Brookshire, G. M. Brown Jr., D. Coursey, R. Innes, S. M. Meyer, S. Polasky. 1999. Why economics matters for endangered species protection. Conservation Biology 13:1257–1261
Smallwood K. S., J. Beyea, M. L. Morrison. 1999. Using the best scientific data for endangered species conservation. Environmental Management 24:421–435
South A. 1999. Dispersal in spatially explicit population models. Conservation Biology 13:1039–1046
Stanford Environmental Law Society. 2001. The Endangered Species Act. Stanford University Press, Stanford, California, 296 pp
Storz J. F. 1999. Genetic consequences of mammalian social structure. Journal of Mammalogy 80:553–569
Strange E., H. Galbraith, S. Bickel, D. Mills, D. Beltman, J. Lipton. 2002. Determining ecological equivalence in service-to-service scaling of salt marsh restoration. Environmental Management 29:290–300
Sugg D. W., R. K. Chesser, F. S. Dobson, J. L. Hoogland. 1996. Population genetics meets behavioral ecology. Trends in Ecology and Evolution 11:338–342
Thomas C. D., I. Hanski. 1997. Butterfly metapopulations. In: I. Hanski, M. E. Gilpin (eds). Metapopulation biology. Academic Press, San Diego, California. pp 359–386
Tietenberg, T. 2004. The tradable permits approach to protecting the commons: what have we learned? Working paper written for National Research Council’s Institutions for Managing the Commons Project. www.colby.edu/personal/thtieten/TT.NRC4.pdf
Turner M. G., G. J. Arthaud, R. T. Engstrom, S. J. Hejl, J. G. Liu, S. Loeb, K. McKelvey. 1995. Usefulness of spatially explicit population-models in land management. Ecological Applications 5:12–16
USFWS (United States Fish and Wildlife Service). 1988. Endangered Species Act of 1973 as amended through the 100th Congress. Department of the Interior, Washington, DC
USFWS (United States Fish and Wildlife Service). 2001. Method for determining the number of available credits for California red-legged frog conservation banks. United States Department of the Interior. Sacramento Fish and Wildlife Office. September 4, 2001
USFWS (United States Fish and Wildlife Service). 2003. Guidance for the establishment, use, and operation of conservation banks. United States Department of the Interior. Memorandum to Regional Directors, Regions 1-7 and Manager, California Nevada Operations. May 2, 2003
Walters C. 1986. Adaptive management of renewable resources. The Blackburn Press, Caldwell, New Jersey, 374 pp
Whitlock M. C. 2004. Selection and drift in metapopulations. In: I. Hanski O. E. Gaggiotti (eds). Ecology, genetics, and evolution of metapopulations. Elsevier Academic Press, San Diego, California. pp 153–174
Wiegand T., K. A. Moloney, J. Naves, F. Knauer. 1999. Finding the missing link between landscape structure and population dynamics: a spatially explicit perspective. American Naturalist 154:605–627
Wilcove D. S., D. Rothstein, J. Dubow, A. Phillips, E. Losos. 1998. Quantifying threats to imperiled species in the United States. Bioscience 48:607–615
Wilcox B. A., D. D. Murphy. 1985. Conservation strategy: the effects of fragmentation on extinction. American Naturalist 125:879–887
Wright S. 1940. Breeding structure of populations in relation to speciation. American Naturalist 74:232–248
Acknowledgments
This work was funded by a USEPA Science to Achieve Results Fellowship. We would like to thank those who helped review earlier versions of the manuscript: J. Bence, J. Blanchong, S. Friedman, R. Shorey, E. Laurent, K. Millenbah, A. Morzillo, L. J. Roberts, E. White, and M. Wilberg. We would also like to thank Matthew Hickey from Greater Cincinnati LISC for discussion regarding economic planning and urban sprawl and two anonymous reviewers whose comments significantly improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bruggeman, D.J., Jones, M.L., Lupi, F. et al. Landscape Equivalency Analysis: Methodology for Estimating Spatially Explicit Biodiversity Credits. Environmental Management 36, 518–534 (2005). https://doi.org/10.1007/s00267-004-0239-y
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-004-0239-y