
Abstract
We investigated the influence of long-term (56 years) grazing on organic and inorganic carbon (C) and nitrogen (N) contents of the plant–soil system (to 90 cm depth) in shortgrass steppe of northeastern Colorado. Grazing treatments included continuous season-long (May–October) grazing by yearling heifers at heavy (60–75% utilization) and light (20–35% utilization) stocking rates, and nongrazed exclosures. The heavy stocking rate resulted in a plant community that was dominated (75% of biomass production) by the C4 grass blue grama (Bouteloua gracilis), whereas excluding livestock grazing increased the production of C3 grasses and prickly pear cactus (Opuntia polycantha). Soil organic C (SOC) and organic N were not significantly different between the light grazing and nongrazed treatments, whereas the heavy grazing treatment was 7.5 Mg ha−1 higher in SOC than the nongrazed treatment. Lower ratios of net mineralized N to total organic N in both grazed compared to nongrazed treatments suggest that long-term grazing decreased the readily mineralizable fraction of soil organic matter. Heavy grazing affected soil inorganic C (SIC) more than the SOC. The heavy grazing treatment was 23.8 Mg ha−1 higher in total soil C (0–90 cm) than the nongrazed treatment, with 68% (16.3 Mg ha−1) attributable to higher SIC, and 32% (7.5 Mg ha−1) to higher SOC. These results emphasize the importance in semiarid and arid ecosystems of including inorganic C in assessments of the mass and distribution of plant–soil C and in evaluations of the impacts of grazing management on C sequestration.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Grazing lands are the largest and most diverse single land type in the United States, covering 336 Mha or about 55% of the country’s total land surface (Sobecki and others 2001), which is more than twice the area of cultivated cropland in the United States (Lal and others 1998). The ecological health of this large land area is important to all citizens because properly managed grazing lands provide positive environmental benefits (e.g., food, clean water supplies, wildlife habitat, and recreational opportunities), whereas poor management can result in environmental and economic degradation (Krueger and others 2002). A major determinant of ecosystem health and stability is the status of the soil organic matter (SOM). SOM is the primary source of many nutrients for plant growth, provides much of the water-holding capacity of the soil, is largely responsible for the formation of stable aggregates that protect the soil from erosion, and is the largest reservoir of carbon (C) and nitrogen (N) in most grazing land ecosystems (Brady and Weil 1999, Follett 2001). Interest has grown in recent years not only in the health and sustainability of grazing lands, but also in the potential of grazing lands to sequester C as part of U.S. efforts to mitigate the greenhouse effect (Follett and others 2001). Sequestering additional organic C into grazing land SOM through the use of recommended best management practices therefore results in the important co-benefits of improving soil quality and thus ecosystem health and sustainability (NRC 1994, Doran and Parkin 1995), while effectively mitigating a large portion of the annual increase in atmospheric CO2 (Follett and others 2001).
Most U.S. grazing lands are in the semiarid and arid western states as native vegetation rangelands (Kimble and others 2001). Historically, rangeland studies have focused on the effects of grazing management on livestock production, plant community composition, and forage production, with relatively few studies evaluating belowground responses to grazing. Reliable estimates of the masses and distributions of soil C and N are therefore limited, and estimates of management-induced changes in C and N cycling are variable and sometimes contradictory (Milchunas and Lauenroth 1993, Schuman and others 2001). Inconsistencies in the reported responses of soil C and N to livestock grazing may be the result of a number of factors that differ among studies, including differences in climate, plant community composition, and grazing history (Milchunas and Lauenroth 1993), and differences in seasonal grazing frequency, intensity and duration (Naeth and others 1991, Reeder and Schuman 2002; Haferkamp and others this issue). Additionally, rangeland ecosystems are generally characterized by a high degree of spatial heterogeneity in soil C and N at both microsite and landscape-level scales (Yonker and others 1988, Tongway and Ludwig 1994, Burke and others 1999) as well as within the soil profile (Reeder and others 2001). Thus, in some instances, failure to detect significant changes in soil C and N with grazing management may be the result of high coefficients of variation caused by comparing grazing treatments across different soil series, including different soil series within a grazing treatment mean, inadequate sampling of the soil profile, or inadequate stratification of sampling to account for inherent spatial heterogeneity within a given soil series (Reeder 2002).
In contrast to croplands and pastures in humid environments, arid and semiarid rangeland soils sequester both inorganic and organic forms of C (Follett and others 2001). Soil inorganic C (SIC) in the form of carbonates is a principal component of many arid and semiarid rangeland soils, with levels ranging from 50 to 310 metric tons of C per hactare (MTC ha−1), and estimated annual rates of carbonate C sequestration ranging from 0.03 to 3.4 millions metric tons of C per year (MMT C yr−1) (Monger and Martinez-Rios 2001). Until recently, only limited attention has been given to C storage as SIC (Elbersen and others 2000). Recent studies have estimated amounts of SIC in grazing land ecosystems (Fitzpatrick and Merry 2000, Monger and Martinez-Rios 2002), but studies evaluating the responses of SIC to grazing management have not been conducted to our knowledge.
We conducted this study to assess the effects of long-term (56 years) grazing at three stocking rates on organic and inorganic C and N storage and cycling within the plant-soil system (to 90 cm) in shortgrass steppe of northeastern Colorado, and to relate differences in soil C and N to grazing-induced changes in plant species composition. We hypothesized that an increase in C4 grasses that Hart (2001) observed with long-term grazing at the heavy stocking rate would result in increases in soil C and N because those species that increased with heavy grazing have higher root-to-shoot ratios than other plant species commonly found on the shortgrass steppe.
Materials and Methods
The experiment was conducted at the Central Plains Experimental Range (CPER), a USDA-ARS research station located 40 km northeast of Fort Collins, Colorado on the northern edge of the shortgrass steppe. Elevations range from 1600 to 1690 m, the average frost-free period is 133 days, and average annual precipitation is 325 mm, of which 50 to 80% occurs during the growing season (Hart 2001). The plant community is approximately 80% perennial grasses (70% C4 warm-season species and 10% C3 cool-season species), 10% forbs, sedges, and cactus, and 10% shrubs. The C4 grasses include Bouteloua gracilis (H.B.K.) Lag. Ex Steud. (the dominant species), Buchloe dactyloides (Nutt.) Engelm., and Aristida longiseta (Steud.) Vasey. The C3 grasses include Pascopyrum smithii (Rydb.) A. Love, and Stipa comata Trin. & Rupr. Other gramminoids include Carex elecharis (Bailey), Elymus elymoides (Raf.) Swezey, and Sporobolus cryptandrus (Torr. Gray). The cactus Opuntia polycantha (Haw.) is abundant, Artemesia frigida (Wild.) is the most common half-shrub, and Sphaeralcea coccinea (Pursh Rydb.) is the most common perennial forb (Hart 2001). Annual production averages 600 kg ha−1, and ranges from 200 to 1100 kg ha−1 (Ashby and others 1993).
A grazing intensity study was established at the CPER in 1939 to evaluate continuous season-long grazing (May–October) at light, moderate, and heavy stocking rates, with 1-ha nongrazed exclosures within each replicate pasture. Grazing treatments were initiated in 1940, and have continued each year to the present time. The light stocking rate averages 18 heifer-days ha−1, which results in 20 to 35% utilization of annual forage production. The moderate stocking rate averages 27 heifer-days ha−1 (40–50% utilization), and the heavy stocking rate averages 37 heifer-days ha−1 (60–75% utilization) (Hart 2001). After 56 years of imposed grazing treatments, we sampled the light and heavy stocking rate treatments, and the 1-ha exclosures within each, for vegetation composition and plant and soil C and N. Line transects (50-m) were established in both grazing treatment pastures and their respective exclosures on the dominant topographical feature of the CPER (gently sloping upland), in areas that had been mapped in 1994 by the Natural Resource Conservation Service as Olney fine sandy loam (fine-loamy, mixed, superactive, mesic, Ustollic Haplargid), the most frequently occurring soil at the CPER. Depending on slope and aspect, depth to precipitated CaCO3 ranges from 25 to 71 cm, and thickness of solum (depth of A and Bt horizons) ranges from 30 to 100 cm, for the Olney soil. The transects for each grazing treatment and respective exclosure were positioned within 50 m of each other on the same slope and aspect to minimize variation in soil properties. No differences in soil loss by erosion were visually apparent (i.e., frequency and degree of pedestalling or rilling) between the grazed treatments and their respective exclosures.
Soil and plant samples were collected at peak standing crop during the first week of August at 10-m intervals along the 50-m transects. At each sampling location, vegetation was clipped to ground level in two 0.18 m2 frames that were positioned on either side of the line transect, approximately 1 m apart (n = 10). Clipped vegetation was partitioned into litter, standing dead, and live biomass by species. (Clipped aboveground plant biomass was influenced by grazing and does not represent annual production.) Root and soil cores were collected with a hydraulic soil sampler. Root cores (10-cm diameter by 30 cm) were collected within the clipped frames and partitioned into 0 to 15 and 15 to 30-cm increments to assess root biomass and root C and N content (n = 10). A deep soil core (4.6-cm diameter by 90 cm) was collected within one of the two clipped frames at each sampling site (n = 5). Deep cores were partitioned into 0 to 4, 4 to 8, 8 to 15, 15 to 30, 30 to 45, 45 to 60, and 60 to 90-cm increments. The first three segments, 0 to 15 cm, generally represent the soil A horizon, the next three segments, 15 to 60 cm, generally represent the Bt horizon (zone of clay accumulation), and the 60 to 90-cm segment generally represents the Bk horizon (zone in which carbonates begin to accumulate). A shallow soil core (4.6-cm diameter by 15 cm) was collected from the second clipped frame at each sampling site and composited by depth increment with the deep core samples to achieve sufficient sample for analyses. Root and soil samples were placed in sealed plastic bags, and transported in coolers to the laboratory for processing.
Bulk density was estimated on separate soil cores taken within 0.5 to 1 m of the deep cores. Intact depth increment segments of these cores were slid into 5-, 10-, or 20-cm lengths of polyvinyl chloride (PVC) tubing. The PVC tube was centered along the depth increment, the soil segment was trimmed at the tube ends, and the soil contained within the tube was transferred to a soil moisture can. Bulk density was calculated as the oven-dry mass of soil collected within the PVC tube divided by the soil core volume within the tube (calculated from the soil core diameter and the length of the PVC tube). Bulk density was estimated for the surface two depth increments combined (0–8 cm).
Soil samples were passed through a 2-mm screen to remove plant crowns and large roots. A subsample of field-moist sieved soil was taken for analysis of inorganic N (NH4-N and NO3-N), which was determined by extraction with 1 M KCl and colorimetric analyses of the extracts with a Technicon autoanalyzer (Braun & Luebbe, Delavan, WI; mention of a commercial product does not imply USDA-ARS endorsement of that product over similar products). The remaining sieved soil was air dried, and samples were stored in sealed plastic bags at room temperature until further analyses. Particle size analysis was determined by the hydrometer method (Gee and Bauder 1986). Total organic C was determined by the Walkley-Black dichromate oxidation procedure (Nelson and Sommers 1982), and inorganic C by a modified pressure-calcimeter method (Sherrod and others 2002). Total soil N was determined by the micro-Kjeldahl procedure described by Schuman and others (1973). An aerobic incubation procedure (21-day, 30°C) measured short-term C and N mineralization as indices of N availability and microbial activity (Reeder and others 1998). Reported mass values of C and N were corrected to an oven-dry basis.
Roots were separated from root cores by flotation and hand washing through a 0.1-mm screen. No attempt was made to distinguish live and dead roots. Plant components were dried at 60°C, weighed, ground, and analyzed for organic C and N content with a Carlo-Erba 1500 automated dry combustion analyzer (CE Elantech Inc., Lakewood, NJ). The C and N concentrations of all plant components were adjusted for ash content.
A four-replication study was initiated in 1939, but replicate pastures were dropped over the years until by 1970 only a single pasture (approximately 128 ha) of each stocking rate remained (Shoop and others 1989). The lack of true replication of grazing treatments limits statistical comparisons of the two stocking rates, but does not preclude comparisons within a selected landscape position or soil type. Data are reported as means ± S.E.M. One-way analysis of variance and mean separation by least significant differences (p ≤ 0.10) were used to test stocking rate effects on plant community composition (Steel and Torrie 1980). Comparisons of total plant biomass and the masses of soil C and N were limited to t-test comparisons of each grazing treatment with its nongrazed exclosure, because analysis revealed that the B horizons (15–60-cm depth) of the lightly grazed pasture and its exclosure were consistently 8 to 10% lower in clay (sandy loam) than the B horizons of the heavily grazed pasture and its exclosure (sandy clay loam). Although originally mapped as an Olney soil, the lower clay content of the B horizon of the lightly grazed pasture and exclosure more closely resembles a Remmit fine sandy loam (coarse-loamy, mixed, mesic, Ustollic Camborthid), the second-most frequently occurring soil series at the CPER. Because of lower clay content, the Remmit soil has innately lower levels of C and N in the B horizon than the Olney soil.
Results
Vegetation Components
Stocking rate influenced plant species composition (Table 1). Warm season C4 grasses (primarily B. gracilis) were dominant in both grazing treatments, but excluding grazing by livestock for 56 years resulted in dominance by cool-season C3 grasses (39% of biomass, primarily S. comata) in the exclosure of the light grazing treatment, and dominance by nongraminoid species (O. polycantha, A. frigida, and forbs, 53% of biomass) in the exclosure of the heavy-grazing treatment. The plant community of the light-grazing treatment was dominated by C4 grasses (45%, primarily B. gracilis), followed by forbs (21%), A. frigida (21%), and O. polycantha (16%). In comparison, C4 grasses (primarily B. gracilis and B. dactyloides) comprised 73% of the biomass under prolonged heavy grazing, and A. frigida and O. polycantha were virtually absent. These results are in general agreement with plant community composition reported by Hart (2001), with the exception that Hart (2001) reported higher overall abundance of O. polycantha than we observed along our transects.
Although plant community composition differed between the light-grazing treatment and its exclosure (Table 1), grazing did not significantly affect total aboveground litter and live biomasses, total root biomass (0–30-cm depth), or total plant biomass (Table 2). In contrast, compared to its exclosure, the plant community under prolonged heavy grazing was strongly dominated by the short grass B. gracilis, and had lower aboveground litter and live biomasses at peak standing crop, and higher root biomass in the 15 to 30-cm depth (Table 2).
Statistical differences in the mass and distribution of C and N in the vegetation components (data not shown) were the same as those observed for plant biomass (Table 2). Total plant C and N for the light-grazing treatment were 4.3 ± 0.3 Mg C ha−1 and 0.12 ± 0.01 MgN ha−1, and 4.8 ± 0.5 Mg C ha−1 and 0.15 ± 0.02 Mg N ha−1 for its exclosure. Total plant C and N for the heavy-grazing treatment were 5.7 ± 0.6 Mg C ha−1 and 0.15 ± 0.02 Mg N ha−1, and 5.8 ± 0.8 Mg C ha−1 and 0.17 ± 0.03 Mg N ha−1 for its exclosure. For both grazing intensities, root C:N ratios (0–15, 15–30, and 0–30-cm depth increments) were significantly higher with grazing than in adjacent nongrazed exclosures. Average root mass C:N ratios (0–30 cm) were 44.8 ± 2.1 and 36.9 ± 1.4 for the light-grazing treatment and exclosure, and 50.9 ± 3.1 and 42.7 ± 2.6 for the heavy-grazing treatment and exclosure.
Soil Response: Bulk Density and Water Content
Soil bulk density of the 0 to 8-cm soil depth increment was significantly higher in the heavy grazing treatment (1.23 ± 0.05 Mg m−3) compared to its nongrazed exclosure (1.02 ± 0.02 Mg m−3), and in the light-grazing treatment (1.18 ± 0.03 Mg m−3) compared to its exclosure (1.09 ± 0.05 Mg m−3). Below the 8-cm soil depth, bulk density was not influenced by grazing and generally ranged from 1.4 to 1.5 Mg m−3. Soil clay content of all depth increments did not differ between each grazing treatment and its respective exclosure, but the clay content of the depth increments between 15 and 60 cm were significantly higher by 8 to 10% in the heavy-grazing treatment and exclosure than in the light-grazing treatment and its exclosure.
At the time of sampling, water content was significantly higher in each depth increment to 60 cm in the soil profile of the heavy-grazing treatment compared to its nongrazed exclosure (Table 3). In comparison, soil water content of the light-grazing treatment was comparable to that of its exclosure in the 0 to 45-cm depth, but was higher in the 45 to 90-cm depth of the grazed treatment. Annual precipitation was normal during the year before sampling, whereas annual precipitation the year of sampling was 45% above the 1939 to 1992 average, with an especially wet spring and early summer.
Soil Response: Carbon and Nitrogen
The light-grazing treatment was comparable to its exclosure in SOC mass throughout the 90-cm soil profile (Table 4). Similarly, total mass of organic N in the 0 to 90-cm profile was comparable in the light-grazing treatment (6.1 ± 0.4 Mg N ha−1) and exclosure (5.7 ± 0.1 Mg N ha−1). In comparison, higher SOC was observed in the 0 to 8, 15 to 30, and 45 to 60-cm depth increments of the heavy-grazing treatment compared to its exclosure (Table 4), with 16% higher SOC in the primary root zone (0–30 cm) and 12% higher SOC in the total soil profile (0–90 cm). Soil organic N was comparable in the heavy-grazing treatment (4.3 ± 0.1 Mg N ha−1) and its exclosure (4.2 ± 0.1 Mg N ha−1) in the 0 to 45-cm depth, but was lower in the 45 to 90-cm depth of the grazed treatment (1.9 ± 0.1 Mg N ha−1) compared to its exclosure (2.4 ± 0.1 Mg N ha−1)
Total mass of NH4-N (0–90 cm) was higher in the soil profiles of both grazed treatments compared to their exclosures (Table 5). This trend was significant in the depth increments between 4 and 90 cm in the light-grazing treatment, and between 4 and 45 cm in the heavy-grazing treatment. Total mass of NO3-N (0–90 cm) was significantly lower in the light grazing treatment compared to its exclosure, whereas NO3-N was significantly higher in the 30 to 60-cm depth of the heavy-grazing treatment compared to its exclosure. Concentrations of NH4-N ranged from 0.5 to 4.4 mg N/kg soil, whereas, concentrations of NO3-N ranged from 0.2 to 2.5 mg N/kg soil.
Significantly lower ratios of organic C:N were observed in the light-grazing treatment compared to its exclosure in depth increments 4 to 8, 15 to 30, and 60 to 90 cm, and in the average C:N ratio of the 0 to 90-cm soil profile (Table 5). In the heavy-grazing treatment, the average C:N ratio for the 0 to 90-cm soil profile was higher with grazing compared to its exclosure, and this trend was significant in the depth increments between 8 and 90 cm. The average immobilization ratio (the ratio of microbially respired CO2 to net mineralized N) for the 0 to 90-cm soil profile was significantly higher in the light-grazing treatment compared to its exclosure (Table 5), and this trend was significant in the 0 to 4, 8 to 15, and 60 to 90-cm-depth increments. In the heavy-grazing treatment, average immobilization ratio for the 0 to 90-cm soil profile also was significantly higher with the grazed compared to the nongrazed treatment, and this trend was significant in the 0 to 4, 8 to 15, and 45 to 60-cm depth increments. The proportion of total soil organic N in the 0 to 90-cm soil profile that was readily mineralized was lower in both grazing treatments compared to their exclosures (Table 5). In the heavy-grazing treatment, this proportion was significantly lower with grazing compared to its exclosure in all but the surface depth increment (0–4 cm). (Average ratios are weighted by the lengths of depth increments that are included in the average.)
Inorganic C mass was larger and depth to lime zone shallower in the soil profiles of the heavy grazing treatment and exclosure compared to the soil profiles of the light-grazing treatment and exclosure (Table 6). In the light-grazing treatment and exclosure, inorganic C represented less than 5% of total C in the surface 60 cm, and 32 to 38% of total C in the 60 to 90-cm-depth increment. Mass of SIC was significantly higher in the depth increments between 0 and 60 cm of the light grazing treatment compared to its exclosure (Table 6). In comparison, total mass of SIC in the soil profile (0–90 cm) was significantly higher in the heavy-grazing treatment compared to its exclosure, and this trend was significant in the 4 to 8-cm depth and in the depth increments between 30 and 90 cm (Table 6). Inorganic C represented 9, 46, and 56% of total C in the 30 to 45, 45 to 60, and 60 to 90-cm-depth increments, respectively, of the nongrazed exclosure within the heavy-grazing treatment, compared to 12, 30, 60, and 60% of total C in the 15 to 30, 30 to 45, 45 to 60, and 60 to 90-cm-depth increments of the heavy-grazing treatment. The 15 to 30-cm-depth increment of the heavy-grazing treatment was a transition zone of patchy and highly variable levels of SIC in the soil profile.
Discussion
In agreement with the findings of studies to characterize the plant community of the CPER (Ashby and others 1993, Milchunas and others 1989, 1990, and 1992, Hart 2001), we found that decades of season-long heavy grazing of the shortgrass steppe resulted in decreased cool season C3 grasses and half-shrubs and increased warm season C4 grasses (primarily B. gracilis), whereas decades of excluding livestock resulted in increased cool season C3 grasses and/or prickly pear cactus. Hart (2001) reported that small patches of O. polycantha observed in the nongrazed exclosures by Fisser and others (1969) increased in frequency and biomass with protection from trampling by livestock. Similarly, cool-season grasses increased in the exclosures with protection from selective grazing by livestock (Hart 2001). We had expected the plant communities of the two nongrazed exclosures to be similar. The lack of similarity (increased cool season grasses in one exclosure, and increased O. polycantha in the other) appears to be caused by the patchiness of subdominant plant species on the shortgrass steppe, and the small number of samples collected in this study being insufficient to capture the uneven distribution of these species.
Because plant root residues are the primary source of SOM, the mass and distribution of SOM depend heavily on the decomposability and distribution patterns of the root systems of the different plant species comprising the vegetation community (Dormaar 1992, Reeder and others 2001). On the shortgrass steppe, 55 to 60% of root biomass generally is concentrated in the 0 to 10-cm depth, and approximately 80% in the 0 to 30-cm depth of the soil profile (Bartos and Sims 1974, Sims and Singh 1978). The dominant species, B. gracilis, is shallow rooted and concentrates approximately 65 to 80% of its root mass in the 0 to 15-cm depth, and approximately 75 to 98% in the 0 to 30-cm depth of the soil profile, whereas more deeply rooted subdominant C3 grass species, such as P. smithii, have approximately one third of root mass below 30-cm depth (Weaver and Darland 1949). The light-grazing treatment and its non-grazed exclosure were comparable in total above- and belowground plant biomass, and these similarities are reflected in the lack of significant difference in amounts of SOC and organic N throughout the soil profile. In contrast, the plant community under prolonged heavy grazing was strongly dominated by the short-grass B. gracilis, and aboveground plant mass was lower and root mass in the 15 to 30-cm depth higher in the heavy-grazing treatment compared to its exclosure. We had expected the strong dominance of B. gracilis with heavy grazing to result in a larger root mass in the 0 to 15-cm depth compared to the nongrazed exclosure. Although this trend was observed, it was not statistically significant because of high variability among replicate samples in estimates of root mass. Mass of SOC, however, was 12% higher in the 0 to 8-cm-depth increment of the heavy-grazing treatment compared to its exclosure. Observed differences in plant biomass components between the heavy-grazing treatment and its exclosure are reflected in 16% higher SOC in the primary root zone (0–30 cm), and the 12% higher SOC in the total soil profile to 90 cm, of the heavy-grazing treatment (Table 4). Wider ratios of organic C:N (Table 5) with heavy grazing reflect an increase in SOC but not in organic N.
Most of the studies conducted on northern Great Plains grasslands that have reported increased SOC with grazing have occurred where grazing has resulted in a significant increase in B. gracilis at the expense of cool-season midgrasses and other species (Smoliak and others 1972, Frank and others 1995, Schuman and others 1999, Reeder and Schuman 2002). The increase in SOC observed with increased B. gracilis suggests that B. gracilis is a more efficient producer of SOC than are other species. With its high root:shoot ratio, B. gracilis transfers more of its photosynthate C to belowground biomass than other species common to the shortgrass steppe (Coupland and Van Dyne 1979, Detling and others 1979). Whether part of the increase in SOC with increased B. gracilis is the result of higher levels of C assimilation remains unclear. In a companion study, LeCain and others (2002) measured C exchange rates for 3 years along the same transects sampled for this study. They reported that, when averaged over the growing season, levels of C assimilation were not significantly different between grazed and nongrazed treatments, but when temperature and precipitation conditions were near-normal, C assimilation was higher in the heavy grazing treatment than in its exclosure during the first half of the growing season, and comparable during the last half of the growing season.
Our results suggest that long-term grazing may alter the quality as well as the quantity of SOM. The ratios of net mineralized N to total organic N were lower with both grazing treatments compared to nongrazed exclosures (Table 5), indicating that long-term grazing decreased the readily mineralizable fraction of SOM. Grazing results in losses of both C and N from the soil:plant system as animal tissue, and further losses of N by volatilization from animal wastes (Woodmansee 1979, Schimel and others 1986). Of the N consumed by cattle, approximately 17 to 21% is converted to animal tissues, 40 to 47% returns to the soil as feces and 23 to 27% as urine, and 12% is lost to the atmosphere from livestock as gaseous products of digestion (N2 and NH3) (Dean and others 1975). Depending on conditions, volatilization losses of NH3 from urine and feces can be as high as 84 to 90% for urine and 80% for feces (Woodmansee 1979). Although C is replenished through photosynthesis, annual inputs of N (by wet and dry deposition and symbiotic and nonsymbiotic N2 fixation) are small at the CPER and may not be sufficient to replenish losses, especially with high grazing intensities (Woodmansee 1979).
Higher immobilization ratios (the ratio of respired CO2 to net mineralized N) also were observed in both grazed treatments compared to nongrazed treatments, suggesting that soil microflora were N-limited in the grazed treatments. Defoliation appears to stimulate root exudates (Dyer and Bokhari 1976), and exudation of C-rich labile organic substrates stimulates microbial activity in the rhizosphere (Biodini and others 1988, Cheng and others 1994). Grazing-induced increases in microbial immobilization of N may limit plant-available N in the soil, and the observed higher C:N ratios of root mass in grazed compared to nongrazed treatments suggest that grazed plants may be unable to take up sufficient N from the soil to replenish the N transported from roots to shoots for regrowth after grazing. Although higher levels of NH4-N occurred in both grazed treatments compared to nongrazed treatments (which suggests higher rather than lower levels of plant-available N in the grazed treatments), the levels of mineral N were very low in all treatments (less than one-half of 1% of total soil N). Because the chemical characteristics and decomposability of roots change during the growing season (Dormaar and others 1981), and because the roots of each species in a grassland system take up nutrients from different regions in the soil at different times and with different intensities (Weaver 1958), data need to be collected throughout the season (rather than at one point in time) in order to accurately assess C and N cycling and N losses (Dormaar 1992).
Our results suggest that, besides altering the characteristics of SOM, grazing also stimulated precipitation of inorganic carbonates. Mass of SIC was slightly but significantly higher (0.7 Mg ha−1) in the 0 to 60-cm depth of the light-grazing treatment compared to its exclosure, whereas mass of SIC was 10.3 Mg ha−1 higher in the 45 to 90-cm depth of the heavy-grazing treatment compared to its exclosure. Whether the increase in SIC with grazing represents newly fixed C, or whether it represents redistribution of existing parent material SIC, is uncertain at this time. The C source for SIC sequestration is CO2 from the atmosphere and from root and soil microbial respiration. This CO2 combines with soil water to form carbonic acid and bicarbonate (HCO3–) in the soil solution, which precipitates as CaCO3 under conditions of decreased soil moisture, decreased pCO2, increased concentration of Ca+2 and HCO3− ions, or increased soil pH (Lal and Kimble 2000). The source of the Ca+2 ions that react with carbonic acid determines whether or not precipitation of carbonates results in a net increase in SIC. If the Ca+2 is derived from a new source, for example, enhanced weathering of calcium silicates or atmospheric additions of noncarbonate Ca+2, then the reaction of Ca+2 with carbonic acid results in sequestration of new SIC as “atmogenic” carbonate. If the Ca+2 ions are derived from preexisting sources, then formation of carbonates is a lateral transfer and reprecipitation of “lithogenic” carbonate, and there is no net sequestration of SIC (Monger and Martinez-Rios 2002).
We hypothesize that the large observed increase in SIC with heavy grazing is a combination of newly sequestered C and redistributed C from deeper in the soil profile. The higher SIC content with heavy grazing compared to its nongrazed exclosure occurred primarily in the 30 to 90-cm depth, below the primary root zone of B. gracilis. At the time of sampling, soil water content was higher throughout the soil profile to 90 cm with the heavy-grazing treatment compared to its adjacent nongrazed exclosure, and this same trend of higher soil water content with the heavy grazing treatment was frequently observed in the surface 30 cm of the soil profile over three growing seasons (LeCain and others 2002). The wetter soil profile of the heavy grazing treatment compared to its exclosure likely reflects the higher water use efficiency and lower levels of evapotranspiration of B. gracilis compared to other grasses of the shortgrass steppe (Pearcy and Ehlinger 1984, Morgan and others 1998). If grazing stimulates root exudation as suggested by Dyer and Bokhari (1976), then higher levels of organic acid root exudates should enhance the weathering of primary calcium-bearing minerals (feldspars and amphiboles) and release new Ca+2 into the soil solution of the primary root zone of the heavily grazed treatment (Mengle and Kirby 1982). Soil water movement through the primary root zone into the subsoil would transport this newly released Ca+2 and HCO3− formed from respired CO2, down into the subsoil beneath the rhizosphere, where lower pCO2 and higher pH facilitate the precipitation of CaCO3 (Lal and Kimble 2000). The higher water content in the lower portion of the soil profile of the heavy grazing treatment also facilitates dissolution of existing parent material carbonates and reprecipitation higher in the profile (30–90 cm) as the soil profile dries during the growing season. Future studies to evaluate C isotopes will help elucidate the sources of the increased SIC in the soil profile of the heavy-grazing treatment.
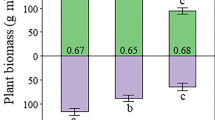
The grazing-induced change in SOC was small relative to the grazing-induced change we observed in SIC. For the long-term heavy-grazing treatment compared to its adjacent nongrazed exclosure, average annual increase in SOC was 0.13 Mg ha−1 yr−1, whereas average annual increase in SIC was 0.29 Mg ha−1 yr−1. Total soil C was 22.8 Mg ha−1 higher in the soil profile (0–90 cm) with heavy grazing compared to the nongrazed exclosure, of which 16.3 Mg ha−1 (68%) was increase in SIC, and 7.5 Mg ha−1 (32%) was increase in SOC (Figure 1). This study demonstrates the importance in semiarid and arid ecosystems of including inorganic carbon in assessments of the amount and distribution of C in the soil-plant system, and of the impacts of management on C sequestration.

Total soil C (0–90-cm depth) after 56 years of season-long light, heavy, or no grazing by livestock (t-test comparisons of heavy grazing and adjacent nongrazed exclosure: p = 0.002 for total C; p = 0.006 for inorganic C; p = 0.006 for organic C).
References
Ashby, M. M., R. H. Hart, and J. R. Forwood. 1993. Plant community and cattle responses to fifty years of grazing on shortgrass prairie. Rangeland Resources Research Unit, RRRU-1. U.S. Department of Agriculture, Agricultural Research Service, Cheyenne, Wyoming.
D. L. Bartos P. L. Sims (1974) ArticleTitleRoot dynamics of a shortgrass ecosystem. Journal of Range Management 27 33–36
M. Biodini D. A. Klein E. F. Redente (1988) ArticleTitleCarbon and nitrogen losses through root exudation by Agropyron cristatum, A. smithii and Bouteloua gracilis. Soil Biology and Biochemistry 20 477–482 Occurrence Handle10.1016/0038-0717(88)90061-2
Brady, N. C., and R. R. Weil. 1999. Soil organic matter. Pages 446–490 in The nature and properties of soils. Prentice Hall, Upper Saddle River, New Jersey.
I. C. Burke W. K. Lauenroth R. Riggle P. Brannen B. Madigan S. Beard (1999) ArticleTitleSpatial variability of soil properties in the shortgrass steppe: The relative importance of topography, grazing, microsite and plant species in controlling spatial patterns. Ecosystems 2 422–438 Occurrence Handle10.1007/s100219900091 Occurrence Handle1:CAS:528:DyaK1MXns12qtrY%3D
W. Cheng D. C. Coleman C. R. Carroll C. A. Hoffman (1994) ArticleTitleInvestigating short-term carbon flows in the rhizospheres of different plant species, using isotopic trapping. Agronomy Journal 86 782–788
R. T. Coupland G. M. Van Dyne (1979) Systems synthesis. R .T. Coupland (Eds) Grassland ecosystems of the world: analysis of grasslands and their uses. International biological programme 18. Cambridge University Press Cambridge, UK 97–106
R. Dean J. W. Ellis R. W. Rice R. E. Bement (1975) ArticleTitle. Journal of Applied Ecology 12 25–29
J. K. Detling M. I. Dyer D. T. Winn (1979) ArticleTitleNet photosynthesis, root respiration and regrowth of Bouteloua gracilis following simulated grazing. Oecologia 41 127–134
J. W. Doran T. B. Parkin (1995) Defining and assessing soil quality. J. W. Doran D. C. Coleman D. F. Bezdicek B. A. Stewart (Eds) Defining soil quality for a sustainable environment. SSSA special publication number 35. Soil Science Society of America Madison, Wisconsin 3–21
J. F. Dormaar (1992) Decomposition as a process in natural grasslands. R. T. Coupland (Eds) Natural grasslands: introduction and western hemisphere. Ecosystems of the World 8A. Elsevier Amsterdam 121–136
J. F. Dormaar S. Smoliak A. Johnston (1981) ArticleTitleSeasonal fluctuations of blue grama roots and chemical characteristics. Journal of Range Management 34 62–64
M. I. Dyer U. G. Bokhari (1976) ArticleTitlePlant-animal interactions: studies of the effects of grasshopper grazing on blue grama grass. Ecology 57 762–772
G. W. W. Elbersen G. R. Davies L. P.R. van Reeuwijk P. T. Janssen (2000) Quantification of the incorporation of atmospheric CO2 in calcic horizons. R. Lal J. M. Kimble H. Eswaran B. A. Stewart (Eds) Global climate change and pedogenic carbonates. Lewis Publishers Boca Raton, Florida 87–95
Fisser, H. G., J. E. Lester, G. C. Mitchell, and L. J. Bledsoe. 1969. Preliminary report of methodology and results for analyses of plant pattern project research on the Pawnee Site. US IBP Grassland Biome Technical Report 9.
R. W. Fitzpatrick R. H. Merry (2000) Pedogenic carbonate pools and climate change in Australia. R. Lal J. M. Kimble H. Eswaran B. A. Stewart (Eds) Global climate change and pedogenic carbonates. Lewis Publishers Boca Raton, Florida 105–120
R. F. Follett (2001) Organic carbon pools in grazing land soils. R. F. Follett J. M. Kimble R. Lal (Eds) The potential of U.S. grazing lands to sequester carbon and mitigate the greenhouse effect. Lewis Publishers Boca Raton, Florida 65–86
R. F. Follett J. M. Kimble R. Lal (2001) The potential of U.S. grazing lands to sequester soil carbon. R. F. Follett J. M. Kimble R. Lal (Eds) The potential of U.S. grazing lands to sequester carbon and mitigate the greenhouse effect. Lewis Publishers Boca Raton, Florida 401–430
A. B. Frank D. L. Tanaka L. Hofmann R. F. Follett (1995) ArticleTitleSoil carbon and nitrogen of Northern Great Plains grasslands as influenced by long-term grazing. Journal of Range Management 48 470–474
G. W. Gee J. W. Bauder (1986) Particle-size analysis. A. Klute (Eds) Methods of soil analysis. Part 1. Physical and mineralogical methods. Agronomy 9, part 1. American Society of Agronomy Madison, Wisconsin 383–412
Haferkamp, M. R., R. K. Heitschmidt, and M. D. MacNeil. Carbon dynamics in the mixed-grass prairie. Environmental Management (this issue).
R. H. Hart (2001) ArticleTitlePlant biodiversity on shortgrass steppe after 55 years of zero, light, moderate or heavy cattle grazing. Plant Ecology 155 111–118 Occurrence Handle10.1023/A:1013273400543
J. M. Kimble R. F. Follett R. Lal (2001) Introduction: The characteristics and extent of U.S. grazing lands. R. F. Follett J. M. Kimble R. Lal (Eds) The Potential of U.S. grazing lands to sequester carbon and mitigate the greenhouse effect. Lewis Publishers Boca Raton, Florida 3–19
Krueger, W. C., M. A. Sanderson, J. B. Cropper, M. Miller-Goodman, C. E. Kelley, R. D. Pieper, P. L. Shaver, and M. J. Trlica. 2002. Environmental impacts of livestock on U.S. grazing lands. Council for Agricultural Science and Technology (CAST) issue paper no. 22. November 2002.
R. Lal J. M. Kimble R. F. Follett C. V. Cole (1998) The potential of U.S. cropland to sequester carbon and mitigate the greenhouse effect. Ann Arbor Press Chelsea, MI 128
R. Lal J. M. Kimble (2000) Pedogenic carbonates and the global carbon cycle. R. Lal J. M. Kimble H. Eswaran B. A. Stewart (Eds) Global climate change and pedogenic carbonates. Lewis Publishers Boca Raton, Florida 1–14
D. R. LeCain J. A. Morgan G. E. Schuman J. D. Reeder R. H. Hart (2002) ArticleTitleCarbon exchange and species composition of grazed pastures and exclosures in the shortgrass steppe of Colorado. Agriculture Ecosystems and Environment 1870 1–15
K. Mengle E. A. Kirkby (1982) Principles of plant nutrition. International Potash Institute Worblaufen-Bern, Switzerland 655
D. G. Milchunas W. K. Lauenroth (1993) ArticleTitleQuantitative effects of grazing on vegetation and soils over a global range of environments. Ecological Monographs 63 327–366
D. G. Milchunas W. K. Lauenroth P. L. Chapman M. K. Kazempour (1989) ArticleTitleEffects of grazing, topography, and precipitation on the structure of a semiarid grassland. Vegetatio 80 11–23
D. G. Milchunas W. K. Lauenroth P. L. Chapman M. K. Kazempour (1990) ArticleTitleCommunity attributes along a perturbation gradient in a shortgrass steppe. Journal of Vegetation Science 1 375–384
D. G. Milchunas W. K. Lauenroth P. L. Chapman (1992) ArticleTitlePlant competition, abiotic, long-and short-term effects of large herbivores on demography of opportunisitic species in semiarid grassland. Oceologia 92 520–531
H. C. Monger J. J. Martinez-Rios (2002.) Inorganic carbon sequestration in grazing lands R. F. Follett J. M. Kimble R. Lal (Eds) The potential of U.S. grazing lands to sequester carbon and mitigate the greenhouse effect. Lewis Publishers Boca Raton, Florida 87–118
J. A. Morgan D. R. LeCain J. J. Read H. W. Hunt W. G. Knight (1998) ArticleTitlePhotosynthetic pathway and ontogeny affect water relations and the impact of CO2 on Bouteloua gracilis (C4) and Pascopyrum smithii (C3). Oecologia 114 483–492 Occurrence Handle10.1007/s004420050472
M. A. Naeth A. W. Bailey D. J. Pluth D. S. Chanasyk R. T. Hardin (1991) ArticleTitleGrazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta. Journal of Range Management 44 7–12
InstitutionalAuthorNameNational Research Council (NRC) (1994) Rangeland health: new methods to classify, inventory and monitor rangelands. National Academy Press Washington, D.C. 182
D. W. Nelson L. E. Sommers (1982) Total carbon, organic carbon, and organic matter. A. L. Page R. H. Miller D. R. Kenney (Eds) Methods of soil analysis, Part 1, 2nd ed., Agronomy Monograph No. 9. American Society of Agronomy Madison, Wisconsin 539–580
R. W. Pearcy J. Ehleringer (1984) ArticleTitleComparative ecophysiology of C3 and C4 plants. Plant, Cell and Environment 7 1–13
Reeder, J. D. 2002. “Overcoming spatial variation in measuring soil carbon stocks and sequestration potential of native rangelands in the western U.S.” In: Smith, CAS. (ed) Soil organic carbon in agriculture: Developing indicators for policy analyses. Proceedings of an OECD Expert Meeting. pp. 193–200
J. D. Reeder C. D. Franks D. G. Milchunas (2001) Root biomass and microbial processes. R. F. Follett J. M. Kimble R. Lal (Eds) The potential of U.S. grazing lands to sequester carbon and mitigate the greenhouse effect. ewis Publishers Boca Raton, Florida 139–166
J. D. Reeder G. E. Schuman (2002) ArticleTitleInfluence of livestock grazing on C sequestration in semi-arid mixed-grass and shortgrass rangelands. Environmental Pollution 116 457–463 Occurrence Handle10.1016/S0269-7491(01)00223-8 Occurrence Handle1:CAS:528:DC%2BD3MXovVertrc%3D Occurrence Handle11822725
J. D. Reeder G. E. Schuman R. A. Bowman (1998) ArticleTitleSoil C and N changes on conservation reserve program lands in the central Great Plains. Soil & Tillage Research 47 339–349
D. S. Schimel W. J. Parton F. J. Adamsen R. G. Woodmansee R. L. Senft M. A. Stillwell (1986) ArticleTitleThe role of cattle in the volatile loss of nitrogen from a shortgrass steppe. Biogeochemistry 2 39–52 Occurrence Handle1:CAS:528:DyaL28XksVWkur4%3D
G. E. Schuman J. D. Reeder J. T. Manley R. H. Hart W. A. Manley (1999) ArticleTitleImpact of grazing management on the carbon and nitrogen balance of a mixed-grass rangeland. Ecological Applications 9 65–71
G. E. Schuman J. E. Herrick H. H. Janzen (2001) The dynamics of soil carbon in rangelands. R. F. Follett J. M. Kimble R. Lal (Eds) The potential of U.S. grazing lands to sequester carbon and mitigate the greenhouse effect. Lewis Publishers Boca Raton, Florida 267–290
G. E. Schuman M. A. Stanley D. Knudsen (1973) ArticleTitleAutomated total nitrogen analysis of soil and plant samples. Soil Science Society of America Proceedings 37 480–481 Occurrence Handle1:CAS:528:DyaE3sXksFaiu74%3D
L. A. Sherrod G. Dunn G. A. Peterson R. L. Kolberg (2002) ArticleTitleInorganic carbon analysis by modified pressure-calcimeter method. Soil Science Society of America Journal 66 299–305 Occurrence Handle1:CAS:528:DC%2BD38XlslOquro%3D
M. Shoop S. Kanode M. Calvert (1989) ArticleTitleCentral plains experimental range: 50 years of research. Rangelands 11 112–117
P. L. Sims J. S. Singh (1978) ArticleTitleThe structure and function of ten western North American grasslands. Journal of Ecology 66 547–572
S. Smoliak J. F. Dormaar A. Johnston (1972) ArticleTitleLong-term grazing effects on Stipa-Bouteloua prairie soils. Journal of Range Management 25 246–250
T. M. Sobecki D. L. Moffitt J. Stone C. D. Franks A. G. Mendenhall (2001) A broad-scale perspective on the extent, distribution, and characteristics of U.S. grazing land. R. F. Follett J. M. Kimble R. Lal (Eds) The potential of U.S. grazing lands to sequester carbon and mitigate the greenhouse effect. Lewis Publishers Boca Raton, Florida 21–63
R. G. D. Steel J. H. Torrie (1980) Principles and procedures of statistics; a biometrical approach. 2nd ed. McGraw-Hill New York, New York
D. J. Tongway J. A. Ludwig (1994) ArticleTitleSmall-scale resource heterogeneity in semi-arid landscapes. Pacific Conservation Biology 1 201–208
J. E. Weaver (1958) ArticleTitleSummary and interpretation of underground development in natural grassland communities. Ecological Monographs 28 55–78
J. E. Weaver R. W. Darland (1949) ArticleTitleSoil-root relationships of certain native grasses in various soil types. Ecological Monographs 19 303–338
R. G. Woodmansee (1979) Factors influencing input and output of nitrogen in grasslands. N. R. French (Eds) Perspectives in grassland ecology. Ecological studies 32. Springer-Verlag New York, New York 117–134
C. M. Yonker D. S. Schimel E. Paroussis R. D. Heil (1988) ArticleTitlePatterns of organic carbon accumulation in a semiarid shortgrass steppe, Colorado. Soil Science Society of America Journal 52 478–483 Occurrence Handle1:CAS:528:DyaL1cXktVGlt7s%3D
Acknowledgements
The authors thank Ernie Taylor, Matt Mortensen, Pam Freeman, and Lucretia Sherrod for their assistance in the collection and analyses of soil and plant samples.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Reeder, J., Schuman, G., Morgan, J. et al. Response of Organic and Inorganic Carbon and Nitrogen to Long-Term Grazing of the Shortgrass Steppe . Environmental Management 33, 485–495 (2004). https://doi.org/10.1007/s00267-003-9106-5
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-003-9106-5