
Abstract
Bird species are hypothesized to join mixed-species flocks (flocks hereon) either for direct foraging or anti-predation-related benefits. In this study, conducted in a tropical evergreen forest in the Western Ghats of India, we used intra-flock association patterns to generate a community-wide assessment of flocking benefits for different species. We assumed that individuals needed to be physically proximate to particular heterospecific individuals within flocks to obtain any direct foraging benefit (flushed prey, kleptoparasitism, copying foraging locations). Alternatively, for anti-predation benefits, physical proximity to particular heterospecifics is not required, i.e. just being in the flock vicinity can suffice. Therefore, we used choice of locations within flocks to infer whether individual species are obtaining direct foraging or anti-predation benefits. A small subset of the bird community (5/29 species), composed of all members of the sallying guild, showed non-random physical proximity to heterospecifics within flocks. All preferred associates were from non-sallying guilds, suggesting that the sallying species were likely obtaining direct foraging benefits either in the form of flushed or kleptoparasitized prey. The majority of the species (24/29) chose locations randomly with respect to heterospecifics within flocks and, thus, were likely obtaining antipredation benefits. In summary, our study indicates that direct foraging benefits are important for only a small proportion of species in flocks and that predation is likely to be the main driver of flocking for most participants. Our findings apart, our study provides methodological advances that might be useful in understanding asymmetric interactions in social groups of single and multiple species.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Associations between individuals of different species are known to occur in a wide variety of taxa including birds, fish, primates, dolphins, etc. (reviewed in Morse 1977; Diamond 1981). Such associations are thought to form for the same reasons as single-species foraging groups, namely for improved protection from predators or better foraging efficiency (Morse 1977; Diamond 1981; Krause and Ruxton 2002). However, some of the mechanisms through which these foraging and anti-predation benefits are obtained are unique to multispecies groups (Morse 1977; Greenberg 2000). While all groups can obtain anti-predation benefits through the dilution effect (Foster and Treherne 1981), many-eyes effect (Pulliam 1973), confusion effect (Neill and Cullen 1974), selfish herd effect (Hamilton 1971) and communal defense (Port et al. 2011), only mixed-species flocks allow individuals to group with other species that have better (Powell 1985) or complementary (Diamond 1981) anti-predatory abilities. Similarly, while copying (e.g. Krebs 1973; Waite and Grubb 1988) and kleptoparasitism (e.g. King and Rappole 2001; Satischandra et al. 2007) benefits can be obtained from both single-species and multispecies groups, feeding in multispecies groups gives access to resources made available by activities of other species (Peres 1993). A recent global analysis found that heterospecific foraging groups in terrestrial birds largely consist of phenotypically similar species (Sridhar et al. 2012), which suggests that the mechanisms that underlie the benefits in these groups are likely to be similar to those of single-species groups. Foraging and predation benefits need not be mutually exclusive explanations for heterospecific association; participation in these groups might allow organisms to reduce their vigilance and correspondingly increase time spent on foraging (Greenberg 2000; Sridhar et al. 2009). At the same time, it is possible that not all group members obtain benefits: certain individuals might be joined and followed by other individuals for the benefits they provide but might not be receiving any benefits in return (‘leaders’ or ‘nuclear species’, Sridhar et al. 2009). Group members might also incur costs in terms of aggression and competition (Greenberg 2000); participation in social groups therefore depends on the trade-off between costs of competition and/or aggression and the benefits accrued.
Mixed-species flocks in terrestrial bird communities (flocks hereon) are the most common and best-studied heterospecific association (Morse 1977; Greenberg 2000). Different approaches have been used to understand why flocks form, which are as follows: (1) experiments, typically in small temperate woodlots, manipulating the possible causative factors (food and predators) and examining changes in flocking levels (e.g. Berner and Grubb 1985; Székely et al. 1989); (2) comparative analyses relating flocking tendencies to species traits linked to vulnerability and to predation (e.g. Buskirk 1976; Thiollay and Jullien 1998; Sridhar et al. 2009); (3) natural experiments that compare foraging and vigilance rates of species, within and outside flocks (e.g. Sullivan 1984; Pomara et al. 2003; Hino 1998); and (4) observational and experimental studies examining the benefits that flock leaders or nuclear species provide to other flock participants (Greig-Smith 1981; Dolby and Grubb 1999, 2000). These different lines of evidence have indicated that both direct foraging and anti-predation benefits are operational in flocks.
The approaches described above, however, have two limitations. First, experimental and ‘natural experiment’ approaches do not allow us to distinguish whether foraging benefits, if any, are obtained directly or reflect indirect benefits mediated through reduced vigilance. Distinguishing between the two is important in order to determine the relative importance of food or predation as the main driver of the evolution and dynamics of flocks. Second, through these approaches, community-wide assessments of the relative importance of direct foraging and predation-related benefits are not feasible. This is especially important, given that studies have suggested that, within the same flock system, some species might obtain foraging, while others obtain anti-predatory benefits (e.g. Hino 1998; Kotagama and Goodale 2004).
In this study, we used intra-flock association patterns to understand the benefits of flock participation in a tropical evergreen forest. Specifically, our approach allows us to determine whether a species is obtaining direct foraging benefits or not in a flock. The rationale of our approach is as follows: Direct foraging benefits, through copying, flushed insects or kleptoparasitism, can only be obtained when birds are physically in close proximity to the specific individuals that provide the benefits (McLean 1984; King and Rappole 2001; Satischandra et al. 2007). These benefits likely require visual monitoring and cues, for which proximity is necessary, particularly in dense tropical habitats (e.g. McLean 1984; Kotagama and Goodale 2004). On the other hand, most mechanisms underlying predation-related benefits are operational at the whole group level. Therefore, birds obtaining predation-related benefits need not be physically close to specific heterospecific individuals within flocks, i.e. just being within the group should suffice. Even if predation-related benefits are obtained from specific flock associates, in the form of eavesdropping on alarm calls of more vigilant species (e.g. Goodale and Kotagama 2005a), close physical proximity is not required because vocal cues can be obtained from much greater distances than visual cues (Goodale and Kotagama 2008). Therefore, if a species chooses locations close to heterospecific individuals within a flock, it is likely to be for direct foraging benefits (also see Hutto 1994).
Direct foraging benefits in flocks can be of different types including flushed insects, copied foraging locations and kleptoparasitism. By relating species’ traits, especially foraging behaviour, to intra-flock association patterns, we can identify the type of direct foraging benefits involved. If the benefit is flushed prey, we expect sallying (catching insects in air with short flights) species to show strong tendencies to be physically proximate to non-sallying heterospecifics (McLean 1984; Satischandra et al. 2007). On the other hand, benefits in the form of copying foraging locations can plausibly be obtained by species from any foraging guild but only from heterospecifics of one’s own foraging guild (Krebs 1973; Waite and Grubb 1988). Benefits in the form of kleptoparasitism can also be obtained by species of all foraging guilds, but, unlike copying benefits, can potentially be obtained from members of both one’s own and other foraging guilds (Greenberg 2000). Apart from foraging behaviour, other species’ traits might also provide clues to the nature of the foraging benefit obtained. Studies have shown that solitary and socially dominant species obtain copying and kleptoparasitizing benefits from intraspecifically gregarious and socially subordinate species, respectively (Morse 1980; Berner and Grubb 1985; Waite and Grubb 1988).
Using the approach described above, we carried out a community-wide assessment of flocking benefits in a tropical forest site in the Western Ghats, India. First, we quantified tendencies of individual species to be physically proximate to heterospecifics within flocks. We then related these tendencies to relevant species traits. Finally, we identified preferred associates, for the subset of species that showed strong non-random within-flock proximity to heterospecifics. Our findings apart, this study develops new sampling methods that will be useful in understanding asymmetric interactions in social groups of multiple as well as single species.
Methods
Study area
The study was carried out in a tropical evergreen forest site within Anshi National Park (15.00978 N, 74.38722 E), Western Ghats, India. Sampling was restricted to January–March in 2010 and 2011; this period coincides with the non-breeding season of most evergreen forest bird species, when flocking activity is highest. Data was collected along 12 pre-existing trails (lengths 1.5–4.5 km) in a 26-km2 focal study area around the Anshi nature camp and Anshi village. The minimum distance between any two trails was at least 500 m. Each trail was surveyed once every 3–4 weeks in both years, giving a total of seven walks per trail.
Recording flock composition
Each trail survey was carried out between 0830 and 1500 hours during which an observer (HS) walked at a slow pace (ca. 1 km/h) searching for flocks. We defined flocks as roving associations of two or more species, staying together for at least 5 min (following Stotz 1993). The 5-min cutoff was used to reduce the possibility of including chance associations of independently moving heterospecifics. We began data collection on each flock only after the initial 5-min period. All data were recorded by the observer speaking into a Sony IC recorder (model ICD-UX71F); this allowed us to record observations and observe birds in the flock simultaneously. An individual bird was designated as being part of the flock if it was within 10 m of at least one other heterospecific individual. This 10-m cutoff was employed as a ‘chain rule’ (Whitehead 2008): if individuals A and B were within 10 m of each other and individuals B and C were within 10 m of each other, A, B and C were considered part of the same flock even if A and C were more than 10 m apart. It is possible that benefits from association are obtained even beyond 10 m; however, because of poor visibility, this distance cutoff is commonly used for flocks in forested habitats (e.g. Hutto 1994). We only included data on roving associations of insectivores and excluded frugivorous flocks; the latter are likely to be the result of species independently aggregating at clumped resources (Stotz 1993; Greenberg 2000) and not outcomes of species interactions. Once a flock was detected, we followed it either until it disintegrated or it became difficult to make observations (observation time = 21 ± 0.9 SE min, N = 360 flocks). We recorded the identities of all species present within each flock, as well as the number of individuals per species, wherever possible. When we were unable to obtain complete counts of individuals, we assigned species to group size classes (0–5, 5–10, 10–15, 15–20, >20 individuals). Though species differed in their detectability, this is unlikely to affect our data because, given the lengths of our observation times, all species present in a flock are likely to have been seen or heard at least once.
Within-flock measures on individual birds
General approach
Given the poor visibility in our study site and that flocks were constantly on the move, a random sampling approach was not practical to obtain adequate data for the individual-based measures described below (within-flock association, foraging behaviour and foraging height). Therefore, the general approach we used was to obtain as many observations of each species in a flock as possible (including potentially multiple observations of the same individual). Given the non-independence of multiple observations of a species within a flock, we calculated a single composite value (average or proportion) per species per flock for each measure.
Physical proximity to heterospecifics
To be certain that patterns in within-flock proximity reflect decisions made by focal individuals, we recorded association only after the individual of interest flew to a new location within the flock. Our preliminary observations revealed that all species, irrespective of foraging behaviour, showed similar patterns of movement within flocks: short hops or flights while searching for prey and feeding, and longer flights when moving to a new foraging location within a flock. We also found that, irrespective of species identity, flights >3 m in length always represented instances of birds moving to a new within-flock location. Therefore, we decided to include all flights >3 m to record associations. It is however possible that, for some species, flights <3 m are also instances of choice of new locations; but we excluded these to use a standardized cutoff across all species. When we observed such an instance, we carried out a focal neighbourhood scan (FNS) to record all flock participants present within a 5-m radius of the focal bird’s new perch. We chose a 5-m cutoff because earlier studies have indicated that birds obtaining direct foraging benefits in flocks generally locate themselves at short distances from benefactors (McLean 1984; Styring and Ickes 2001; Kotagama and Goodale 2004). Moreover, given the dense nature of the evergreen forest habitat, it is reasonable to assume that birds will need to be at distances <5 m to be able to visually monitor other birds for foraging opportunities. We tried to obtain at least one FNS for every species in a flock. Flight distances and cutoff point for FNSs (5 m) were estimated visually. We did not include flights that involved the focal individual: (1) chasing or being chased by another bird; (2) responding to the detection of a predator; (3) flying out of the flock vicinity; and (4) making a foraging attempt. Given that flights were unpredictable events, we continuously scanned the flock looking for such instances by flock participants.
In the 2011 season, we also examined if the choice of within-flock locations was different from random by doing a paired random neighbourhood scan (RNS) for every FNS carried out as described above. We basically asked: if the focal bird had chosen to fly the same distance and to the same height but in a different direction, how many associates would it have had? To do this, we picked a point that was matched for height and distance from original perch but in a randomly determined direction and counted flock participants within a 5-m radius of this point. To determine the direction of the RNS, we added 0, 45, 90, 135, 180, 225, 270 or 315 to original flight direction in degrees in a clockwise fashion; the number to add for a particular RNS was chosen by drawing lots, i.e. writing these numbers on a piece of paper each and picking one at random. If there was no suitable perching substrate at the chosen RNS point, we used the nearest available substrate to the RNS point instead. Given that the height and distance for each RNS was dependent on the paired FNS, RNSs were always carried out after the paired FNS, i.e. we were unable to randomize the order of the FNSs and RNSs. However, we do not think this introduces any systematic bias in our results. Moreover, since the method employed was uniform across our points of comparison (species), even if there was bias, it would have affected all species equally.
Species traits
Body mass (gram) values were obtained from Dunning (2008), while the other traits were calculated from field data. We carried out focal animal sampling of arbitrarily chosen individuals in flocks to quantify the relative proportions of different foraging behaviours. Foraging behaviour categories (modified from Remsen and Robinson 1990) included the following: (1) glean (prey picked from upper and lower leaf surfaces through active searching); (2) search (prey obtained by probing live and dead leaf clusters and dead wood); (3) bark-probe (prey obtained by pecking and probing tree trunks, branches and twigs); (4) sally-glean (prey obtained from leaf surfaces and bark with flights from stationary positions); (5) sally (prey caught in the air with a flight); and (6) kleptoparasitize (prey snatched or stolen from other flock participants). Focal individuals were observed until they were lost from view. We excluded the first 10 s of observation of each focal animal sample to remove any biases caused by differences in detectability of different behaviours. Given that kleptoparasitizing was a rare behaviour, we kept records of all instances seen outside focal sampling as well. We also recorded the heights of flock participants, in classes of 0–1, 1–2, 2–4, 4–8, 8–16, 16–32 and >32 m.
Analysis
General approach
Our sampling design of repeatedly walking the same trails raises the issue of non-independence of flocks. In fact, our sampling revealed ‘hotspots’ of flock activity, i.e. locations along trails where flocks were regularly seen. However, it is appropriate to treat these flocks as independent data points for two reasons. First, our field observations showed that flock formation and disintegration happens on a minutes-to-hours timescale. Therefore, even flocks seen at the same location on different days are outcomes of independent formation events, often comprising different species compositions (Sridhar et al. 2013). Nevertheless, given that there was likely to be pseudoreplication at the individual level and that the conditions that promote flocking (resources or predators) are likely to be similar in the same location at different times, we also reran all our analyses after pooling data on flocks from the same locations, considering these as hotspots. For this hotspot analysis, all flocks within 200 m of each other were designated as belonging to the same hotspot. The distance between any two ‘hotspots’ was at least 500 m. Our ‘flockwise’ and ‘hotspotwise’ results were qualitatively similar, and therefore, we present only the former in the main text. Corresponding ‘hotspotwise’ results are provided in Online Resource 1. We also ran all analyses for each individual trail separately and found the results to be qualitatively similar across trails and compared to the overall results (Online Resource 2).
Within-flock physical proximity to heterospecifics
For each species, we calculated the average proportion of FNSs per flock that included heterospecifics. Given that only a small percentage of FNSs (<0.5 %) contained multiple heterospecific individuals, we restricted our analysis to only presence–absence of heterospecifics. We also calculated for each species within a flock, the proportion of RNSs that included heterospecifics. We then used McNemar’s test to examine whether, for a species, the proportion of FNSs that included heterospecifics was significantly different from the proportion of RNSs that included heterospecifics. We also used the same approach described above to characterize intra-flock association of species with conspecifics (Online Resource 3).
Relationship between species traits and within-flock physical proximity to heterospecifics
We examined the relationship between average proportion of FNSs that included heterospecifics and species traits including body mass, average intraspecific group size, average foraging height and foraging behaviour guild separately. We use body mass as a surrogate of social dominance because larger species tend to dominate smaller species (Shelley et al. 2004; but see Hino 2005). Foraging height was included in the analysis to ensure that detectability of associates in FNSs for a particular species was not influenced by its foraging height, i.e. that detectability of associates does not decrease higher in the canopy. To calculate foraging height averages we used the midpoints of height classes. To calculate average intraspecific group size we used the midpoints of group size classes. Foraging behaviour guilds included gleaners, searchers, bark-probers, sally-gleaners and sallyers; we excluded kleptoparasitizing because it contributed negligible proportions to the diets of all species. Each species was assigned to a guild based on the foraging behaviour it employed most commonly. There was little ambiguity in the assignment of foraging behaviour guilds because all species employed a particular behaviour in more than 50 % of the observations. Given the non-normality of the data, we used randomization equivalents of conventional parametric tests to examine the strength of the relationships. Randomized Pearson’s correlations were used for body mass, foraging height and intraspecific group size tests, while randomized ANOVA was used for foraging guild tests. All analyses were implemented in the software ECOSIM (Gotelli and Entsminger 2001) and based on 1,000 iterations.
Preferred associates
We used permutation tests to examine whether certain species were disproportionately represented as associates in FNSs. This analysis was performed only for focal species with higher than expected proportions of heterospecifics in FNSs, as determined through comparison with paired RNSs. For each focal species separately, we first calculated the observed number of times each associate species was present in its neighbourhood scans (O). We then calculated the expected number of times each associate species would occur in its FNSs, if the focal species showed no preference. For this, we constructed an available species pool consisting of the summed composition of all the flocks in which the focal species participated. We then randomly drew FNSs from this pool, equal to the observed FNSs for that species, and counted the number of times each associate species occurred in them. We created 1,000 such sets of simulated FNSs and calculated the average number of times each associate species was present in simulated FNSs (μ). The association strength of a focal species with a particular associate was calculated as:
Where σ = standard deviation of number of simulated FNSs in which species occurred (following Srinivasan et al. 2010). We considered association strength values greater than 2 as indicative of preferential association; association strengths greater than 2 were generally significant at p < 0.05.
Results
Flock composition
We sampled 370 flocks including a total of 59 bird species. The flocks included, on average, 8.6 ± 0.2 (SE) species and 28.7 ± 0.7 (SE) individuals. Seven species occurred in 50–75 % of the flocks, 5 species occurred in 20–50 % of flocks, 14 species occurred in 10–20 % of flocks and 33 species occurred in <10 % of flocks (Online Resource 4).
Within-flock physical proximity to heterospecifics
We recorded 2,385 FNSs across 370 flocks totally but only included species for which we obtained at least one FNS each in at least five flocks in the following analyses. Species clustered into two groups based on association with heterospecifics (Table 1): a small proportion of species (5/29) had heterospecifics in >80 % of FNSs, while the rest of the species (24/29) had heterospecifics in ≤30 % of FNSs. Further, comparisons with paired RNSs showed that only the five species that had heterospecifics in >80 % of FNSs also had a significantly higher proportion of FNSs that included heterospecifics than expected by chance (McNemar’s test, p < 0.05; Table 1). For all other species, proportion of FNSs that included heterospecifics was not different from random expectation (Table 1).
Across all species, the average proportion of FNSs that included conspecifics ranged between 0.0 and 0.3. As expected, species that were more intraspecifically gregarious (average intraspecific group size > 2) had higher proportion of FNSs that included conspecifics (Online Resource 3). We found three species to have significantly higher proportion of FNSs that included conspecifics than expected by chance (Online Resource 3; McNemar’s test, p < 0.05).
Relationship between species traits and association patterns
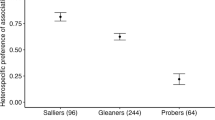
The average proportion of FNSs that included heterospecifics was significantly different across foraging guilds (F ratio = 146.48, randomization p < 0.001; Fig. 1; Table 1). The difference was mainly due to the sallyers guild having a much higher average proportion of FNSs that included heterospecifics (0.88) compared with other foraging guilds (range in average = 0.03–0.14). The average proportion of FNSs that included heterospecifics was not related to the body mass of species (r = −0.01, randomization p = 0.49, n = 29; Table 1) or foraging height (r = −0.09, randomization p = 0.35, n = 29; Table 1), but showed a significant but weak negative relationship with intraspecific group size (r = −0.29, randomization p = 0.003, n = 29; Table 1).

Average proportion of focal neighbourhood scans that included heterospecifics, for different foraging guilds in mixed-species bird flocks in Anshi National Park, India. Data labels indicate the number of species in each foraging guild
To confirm that the difference in association pattern between sallyers and non-sallyers was not due to differences in the types of flocks of these two groups, we also carried out within-flock comparisons. We compared, in a pairwise fashion for each flock, the difference in proportion of FNSs that included heterospecifics for sallyers versus non-sallyers. Across all flocks, we found that sallyers had significantly higher proportion of FNSs that included heterospecifics than non-sallyers (sallyers > non-sallyers: 229 flocks; non-sallyers > sallyers: 2 flocks; sallyers = non-sallyers: 13 flocks; McNemar’s test p < 0.001).
Preferred associates
All five sallying species, which had higher than expected proportions of FNSs that included heterospecifics, showed significant preferential association with at least one other species; moreover, all preferred associates were from foraging guilds other than sallying (Table 2 and Online Resource 5).
Discussion
Our study used intra-flock association patterns to understand the benefits that different species obtain from mixed-species flocks, based on the following logic: to obtain direct foraging benefits, flock participants need to be physically proximate to particular heterospecific individuals within flocks (McLean 1984; King and Rappole 2001; Satischandra et al. 2007). On the other hand, to obtain predation-related benefits, physical proximity to specific individuals is not required because the mechanisms underlying predation-related benefits are usually operational at the whole group level (also see Hutto 1994).
Our study found that only a small subset of the total species (5/29), i.e. all members of the sallying foraging guild, associated closely with heterospecifics within flocks. The majority of species (24/29) chose locations randomly with respect to heterospecifics. Here, we discuss what our results mean for understanding the benefits of mixed-species flocking and also the validity of the approach.
A few studies have attempted to quantify intra-flock association patterns (e.g. Hutto 1994; Latta and Wunderle 1996; Kotagama and Goodale 2004). The approach in these studies was to randomly pick focal individuals and identify its close neighbours. Association patterns determined using this approach are not easily interpretable because they can result from three different mechanisms: (1) attraction of focal individuals to neighbours; (2) attraction of neighbours to focal individuals; and (3) focal individuals and neighbours moving independently but being physically proximate just by chance. In our study, by only identifying associations after focal individuals flew to new locations, we ensure that patterns are outcomes of decisions made by focal individuals. Moreover, by comparing association patterns to what one would expect if focal individuals flew randomly, we also verify that these associations are a result of active choice. In other words, the association patterns we describe are both directional and non-random.
Pattern of association of sallying species
Our study revealed a stark difference in within-flock association patterns between sallyers and non-sallyers. All five sallying species showed high levels of non-random association with heterospecifics within flocks, while for all other bird species, association with heterospecifics was low and not different from random. We also detected a weak but significant negative relationship between intraspecific group size and proportion of FNSs that included heterospecifics, i.e. greater the intraspecific group size, lower was the proportion of FNSs that included heterospecifics. This relationship is likely to be a secondary consequence of two other patterns: (1) all sallying species were usually intraspecifically solitary in flocks (Table 1) and (2) intraspecifically gregarious species tended to choose locations close to conspecifics (Online Resource 3). Therefore, it is reasonable to infer that sallying behaviour serves as a singular demarcator of differences in association patterns within flocks.
Why do sallying species show strong within-flock association with heterospecifics? Comparisons with paired RNSs in flocks clearly show that the associations result from active choice. Given that all sallying species are highly vigilant themselves (Munn 1986; Goodale and Kotagama 2005a), the physical proximity to heterospecifics is unlikely to be for vigilance. Therefore, it is likely that the association is for direct foraging benefits. Given that preferred associates of all sallying species belonged to non-sallying foraging guilds (Table 2), it is likely that benefits obtained are either flushed prey or kleptoparasitism (Swynnerton 1915; McLean 1984; Satischandra et al. 2007; Greenberg 2000). Foraging benefits gained by copying are unlikely to be obtained from individuals of other foraging guilds (Krebs 1973; Waite and Grubb 1988). While we cannot definitively say how much of the prey obtained by sallyers was the result of flushing, our field observations and earlier studies indicate that it is likely to form a substantial portion of sallyers’ diets (McLean 1984; Herremans and Herremans-Tonnoeyr 1997; King and Rappole 2001; Styring and Ickes 2001; Veena and Lokesha 1993; Satischandra et al. 2007). Though we observed very few instances of kleptoparasitism during focal animal sampling, ad hoc observations revealed many attempts by the three drongo species (Dicrurus aeneus, 11 attempts on 5 species; Dicrurus leucophaeus, 31 attempts on 10 species; Dicrurus paradiseus, 41 attempts on 14 species). In all cases, the ‘victims’ came from non-sallying foraging guilds, a pattern similar to what has been reported in earlier studies on drongo foraging in flocks (King and Rappole 2001; Styring and Ickes 2001; Veena and Lokesha 1993; Satischandra et al. 2007). Moreover, the ‘victims’, most often, were the same species which were preferred associates in the FNSs. We observed no instances of kleptoparasitism by the other two sallying species namely Hypothymis azurea and Terpsiphone paradisi.
An interesting future line of research might be to examine interactions among sallying species in flocks, especially drongos. Though our results show that each drongo species had a unique set of preferred associate species (Table 2), there was also some overlap (e.g. Pericrocotus flammeus was a preferred associate of both D. aeneus and D. leucophaeus). It would be interesting to examine how competition for associates is mediated or avoided when different drongo species that share preferred associates co-occur in the same flock. The non-drongo sallyers, H. azurea and T. paradisi, showed no overlap in preferred associates with any of the drongos and were instead almost obligately associated with a single species, Phylloscopus occipitalis. However, despite sharing the same singular preferred associate, we found no evidence of competition between H. azurea and T. paradisi—we observed many instances of both species associating with P. occipitalis within the same flock to feed on insects flushed by the latter. This could be because, given the large numbers of individuals of P. occipitalis in a flock on average (15.0 ± 0.50 SE), insects flushed by P. occipitalis might not be a limiting resource.
Pattern of association of non-sallying species
The majority of species in the bird community, including all members of non-sallying foraging guilds, showed low levels of association with heterospecifics within flocks, not different from random expectation (although the power for some of these tests was low (Table 1), the general trend is as described). It is therefore likely that none of these species obtain copying, flushed prey or kleptoparasitism-related benefits.
What benefits might these species obtain by participating in flocks? Earlier studies have suggested that certain direct-foraging benefits could be obtained from group level mechanisms. Diamond (1987) proposed that foraging in mixed-species flocks allows birds to overcome the territorial defenses of conspecifics. Foraging in mixed-species flocks might also help birds monitor the foraging of other participants and avoid foraging in already exploited areas (Morse 1970; Austin and Smith 1972). However, neither of these mechanisms is likely to be operational in our study system because of the wide variation in foraging micro-habitats used by participants within the same flocks; flocks, on average, contained 3.7 (±0.06 SE) unique foraging guilds and each guild is specialized on a different foraging microhabitat. It is unlikely that group level foraging benefits described above are plausible across species using different foraging microhabitats. Another potential group level foraging benefit in flocks is the finding of clumped insect prey (Greig-Smith 1978). However, our field observations showed that flocks are constantly on the move and range over large distances (often >100 m) for feeding; therefore, flocking is unlikely to be a mechanism to find clumped food for non-sallyers, at least in our study site.
It is likely, therefore, that species that do not associate closely with heterospecifics within flocks are obtaining anti-predation benefits. Most of the mechanisms proposed for predation-related benefits in groups, including dilution effect (Foster and Treherne 1981), many-eyes effect (Pulliam 1973), confusion effect (Neill and Cullen 1974) and communal defense (Port et al. 2011), are operational at the whole group level. Therefore, these mechanisms do not predict non-random choice of locations of species within flocks. The selfish herd effect (Hamilton 1971) does predict differential susceptibility depending on position within flock, but not at the level of individual associations. Moreover, predation-related grouping benefits can be profitably obtained from species of different foraging guilds because they are likely to share the same predators. In our study site, the most likely predators of flock participants are likely to be two to three species of small forest raptors.
It is, however, possible that anti-predation benefits are also obtained through mechanisms that do not operate at the group level. Studies have shown that certain kinds of species, e.g. intraspecifically gregarious birds or sentinels, are often the quickest to alarm call in response to predators (Goodale and Kotagama 2005b). Therefore, species might choose to associate non-randomly with such species within flocks. However, given that these benefits are based on vocal cues, they can be obtained at much larger distances than that required for direct foraging benefits, i.e. >5 m (Goodale and Kotagama 2008). In other words, species which show random association with heterospecifics at the 5-m scale might nevertheless be associating non-randomly with heterospecifics at distances greater than 5 m to obtain anti-predatory benefits. In sum, irrespective of the specific mechanisms involved, species that are not physically proximate to heterospecifics at the 5-m scale are likely to be obtaining anti-predatory benefits in flocks. However, it is important to note that our inference that most species in flocks obtain anti-predatory benefits only follows as a corollary of our finding regarding direct foraging benefits. Direct tests are required to establish the anti-predatory function of flocks.
Among non-sallying species, we also found a tendency for non-random association with conspecifics, especially among the more intraspecifically gregarious species. It is possible that such species are obtaining direct foraging benefits from conspecifics and predation-related benefits from the larger mixed-species group.
We also confirmed that the differences between sallyers and non-sallyers were not a reflection of these two species types participating in different types of flock. Our within-flock comparisons indicated that even within the same flocks, sallyers showed significantly greater proximity to heterospecifics compared to non-sallyers. In other words, the same individual flocks include a few species obtaining direct foraging benefits and the remaining obtaining predation-related benefits.
Caveats to the approach
A drawback of our approach was that we only measured associates within fixed 5 m radii of focal individuals. While we acknowledge that a better characterization of association patterns would be obtained from actual measurement of distances to associates (e.g. Kotagama and Goodale 2004), we were unable to do so in our study site because of detectability issues. Preliminary observations showed that while we were relatively certain of detecting 100 % of individuals present within 5 m of focal birds, the probability of missing individuals increased beyond this distance. Moreover, this probability was different for different associate species depending on their individual detectability. Given these issues, using actual measurements of distances would have introduced systematic biases in our association patterns, and hence, we decided to use a fixed radius approach instead. Our findings are therefore contingent on the specific spatial scale of our observations. The crucial question is whether the 5-m radius is biologically meaningful. Earlier studies have found that direct foraging benefits (copying, feeding on flushed insects or kleptoparasitism) are usually obtained at distances in the range of a few metres in flocks (McLean 1984; Styring and Ickes 2001; Kotagama and Goodale 2004). This is true even in relatively open habitats (McLean 1984; Hutto 1994); hence, it is even more likely in our dense tropical evergreen forest site. Although we do not know the actual range of visibilities of birds in these forests, it is reasonable to expect that the visual monitoring of associates required to obtain direct foraging benefits is likely to be possible only at short distances.
Conclusion
In sum, our results suggest that at the whole community level, only a small proportion of species obtain direct foraging benefits in flocks, and as a corollary, the majority of species are likely to be getting predation-related benefits. This was the case even at the individual flock level because, on average, only one in four species in any given flock was a sallyer. In other words, each individual flock was typically a combination of a few species obtaining direct foraging benefits and many species obtaining predation-related benefits. Given that direct foraging or anti-predatory benefits are the only likely benefits in flocks, it is reasonable to infer that mixed-species flocks in our study site are mainly a predation-related phenomenon. Moreover, a low percentage of sallyer species among flock participants is a pattern seen in flock systems across the world (range in percent sallyer species across 32 studies—0–33 %; Online Resource 6); therefore, our inferences about the relative importance of direct foraging and anti-predation benefits in flocks are likely to apply to flocks in general. Additional support for the role of flocks as anti-predatory groupings is provided by studies that show that species in flocks readily respond to heterospecific alarm calls (Goodale and Kotagama 2005b) and that areas without predators tended to have less flocking (e.g. Beauchamp 2004). Additionally, studies have also shown that kleptoparasitic species such as drongos are ‘tolerated’ in flocks only because they provide anti-predatory benefits in return (Ridley and Raihani 2007).
Recently, it has also been shown that the proportion of sallyers and gleaners might reflect the relative proportions of foraging resources for these two groups in an area (Srinivasan and Quader 2012). Therefore, it is possible that sallyer representation in flocks, and consequently the importance of direct foraging benefits in flocks, could be higher in areas where underlying resource distributions are different. Moreover, even if sallyer numerical representation in flocks is fairly low, their functional importance can be disproportionately high because studies have shown sallying species to often be ‘nuclear species’ or leaders of flocks (Srinivasan et al. 2010). As a consequence, the component of direct foraging benefits in flocks, though beneficial only to a small number of participants, can have disproportionately large effects as a driver of flock evolution and dynamics.
Finally, our results shed light on fundamental differences in mechanisms underlying direct foraging and predation-related benefits in flocks. A recent global analysis of flock composition revealed that flocks are largely composed of phenotypically similar species (Sridhar et al. 2012). Taken together with our finding that majority of flock participants obtain predation-related benefits, these results suggest that mechanisms underlying predation-related benefits are dependent on species being similar. Direct foraging benefits, on the other hand, as shown by our study, are obtained through mechanisms, i.e. flushed prey and kleptoparasitism, which are dependent on the benefactors being ecologically different from the beneficiaries. Additional support for this inference comes from studies that have shown bird species to even join members of completely different lineages, such as mammals, to obtain direct foraging benefits (e.g. Boinsky and Scott 1988; Oommen and Shanker 2009). Taken together, these findings suggest that while mechanisms through which anti-predatory benefits are obtained in tropical flocks might be similar to single-species groups, direct foraging benefits are obtained through mechanisms only possible in a multispecies condition.
Our findings apart, this study also demonstrates the utility of a new method to measure associations within social groups. By measuring associations based on choice of new locations within flocks by focal individuals and by comparing choice of locations to what is expected by chance, we were able to examine if associations were asymmetric and non-random. We believe this can be a useful alternative to conventional ‘nearest neighbour’ approaches (e.g. Hutto 1994; Latta and Wunderle 1996; Kotagama and Goodale 2004; Whitehead 2008) in a variety of contexts. First, our method can be directly extended to flocks in other geographical areas. Our results are somewhat in contrast to studies on flocks from temperate areas that have indicated a more important role for other forms of direct foraging benefits such as copying benefits exchanged between core members of flocks (Krebs 1973; Waite and Grubb 1988). At the same time, predation-related benefits in flocks have also been documented in temperate communities (e.g. Székely et al. 1989). Therefore, it will be useful to use this approach to understand the relative importance of foraging and predation-related benefits in temperate flocks. Second, our approach can also be extended to understand the nature of benefits in mixed-species groups in other taxa. Studies have indicated that, similar to sallyers in bird flocks, participants in mixed-species groups in other taxa feed too on prey flushed by other group members (e.g. fishes: Aronson and Sanderson 1987; primates: Peres 1992). Finally, going beyond mixed-species groups, our method might also find application in analogous situations in single-species groups, namely producer–scrounger relationships (e.g. Barnard and Sibly 1981). More generally, the methodological advances we propose will be useful in measuring intra-group associations and quantifying individual importance of group members in any social group context in which there is likely to be asymmetry in interactions.
References
Aronson RB, Sanderson SL (1987) Benefits of heterospecific foraging by the Caribbean wrasse Halichoeres garnoti (Pisces: Labridae). Environ Biol Fish 18:303–308
Austin GT, Smith EL (1972) Winter foraging ecology of mixed insectivorous bird flocks in oak woodland in southern Arizona. Condor 74:17–24
Barnard CJ, Sibly RM (1981) Producers and scroungers: a general model and its application to captive flocks of house sparrows. Anim Behav 29:543–550
Beauchamp G (2004) Reduced flocking of birds on islands with relaxed predation. Proc R Soc Lond B 271:1039–1042
Berner TO, Grubb TC Jr (1985) An experimental analysis of mixed-species flocking in birds of deciduous woodland. Ecology 66:1229–1236
Boinsky S, Scott PE (1988) Association of birds with monkeys in Costa Rica. Biotropica 20:136–143
Buskirk WH (1976) Social systems in a tropical forest avifauna. Am Nat 110:293–310
Diamond JM (1981) Mixed-species foraging groups. Nature 292:408–409
Diamond JM (1987) Flocks of brown and black New Guinean birds: a bicoloured mixed-species foraging association. Emu 87:201–211
Dolby AS, Grubb TC Jr (1999) Functional roles in mixed-species flocks: a field manipulation. Auk 116:557–559
Dolby AS, Grubb TC Jr (2000) Social context affects risk taking by a satellite species in a mixed-species foraging group. Behav Ecol 11:110–114
Dunning JD Jr (2008) CRC handbook of avian body masses. CRC, Boca Raton
Foster WA, Treherne JE (1981) Evidence for the dilution effect in the selfish herd from fish predation on a marine insect. Nature 293:466–467
Goodale E, Kotagama SW (2005a) Testing the roles of species in mixed-species bird flocks of a Sri Lankan rain forest. J Trop Ecol 21:669–676
Goodale E, Kotagama SW (2005b) Alarm calling in Sri Lankan mixed-species bird flocks. Auk 122:108–120
Goodale E, Kotagama SW (2008) Response to conspecific and heterospecific alarm calls in mixed-species bird flocks of a Sri Lankan rainforest. Behav Ecol 19:887–894
Gotelli NJ, Entsminger GL (2001) EcoSim: null models software for ecology, version 1.0. http://garyentsminger.com/ecosim/index.htm
Greenberg R (2000) Birds of many feathers: the formation and structure of mixed-species flocks of forest birds. In: Boinski S, Gerber PA (eds) On the move: how and why animals travel in groups. The University of Chicago Press, Chicago, pp 521–559
Greig-Smith PW (1978) Imitative foraging in mixed-species flocks of Seychelles birds. Ibis 120:233–235
Greig-Smith PW (1981) The role of alarm responses in the formation of mixed-species flocks of heathland birds. Behav Ecol Sociobiol 8:7–10
Hamilton WD (1971) Geometry for the selfish herd. J Theor Biol 31:295–311
Herremans M, Herremans-Tonnoeyr D (1997) Social foraging in the Forktailed Drongo Dicrurus forficatus: beater effect or kleptoparasitism? Bird Behav 12:41–45
Hino T (1998) Mutualistic and commensal organization of avian mixed-species foraging flocks in a forest of western Madagascar. J Avian Biol 29:17–24
Hino T (2005) Resident males of small species dominate immigrants of large species in heterospecific, winter bird flocks. Ornithol Sci 4:89–94
Hutto RL (1994) The composition and social organization of mixed-species flocks in a tropical deciduous forest in western Mexico. Condor 96:105–118
King DI, Rappole JH (2001) Kleptoparasitism of laughingthrushes Garrulax by Greater racket-tailed drongos Dicrurus paradiseus in Myanmar. Forktail 17:121–122
Kotagama SW, Goodale E (2004) The composition and spatial organization of mixed-species flocks in a Sri Lankan rainforest. Forktail 20:63–70
Krause J, Ruxton GD (2002) Living in groups. Oxford University Press, Oxford
Krebs JR (1973) Social learning and the significance of mixed-species flocks of chickadees (Parus spp.). Can J Zool 51:1275–1288
Latta SC, Wunderle JM (1996) The composition and foraging ecology of mixed-species flocks in pine forests of Hispaniola. Condor 98:595–607
McLean IG (1984) Feeding association between fantails and saddlebacks: who benefits? New Zeal J Ecol 7:165–168
Morse DH (1970) Ecological aspects of some mixed-species foraging flocks of birds. Ecol Monogr 40:119–168
Morse DH (1977) Feeding behavior and predator avoidance in heterospecific groups. Bioscience 27:332–339
Morse DH (1980) Behavioural mechanisms in ecology. Harvard University Press, Cambridge
Munn CA (1986) Birds that cry ‘wolf’. Nature 391:143–145
Neill SRSJ, Cullen JM (1974) Experiments on whether schooling by their prey affects the hunting behavior of cephalopods and fish predators. J Zool 172:549–569
Oommen M, Shanker K (2009) Shrewd alliances: mixed foraging associations between treeshrews, greater racket-tailed drongos and sparrowhawks on Great Nicobar Island, India. Biol Lett 6:304–307
Peres CA (1992) Prey capture benefits in a mixed-species group of Amazonian tamarins Saguinus fuscicollis and S. mystax. Behav Ecol Sociobiol 31:339–347
Peres CA (1993) Anti-predator benefits in a mixed species group of Amazonian tamarins. Folia Primatol 61:61–76
Pomara LY, Cooper RJ, Petit LJ (2003) Mixed-species flocking and foraging behavior of four Neotropical warblers in Panamanian shade coffee fields and forests. Auk 120:1000–1012
Port M, Kappeler PM, Johnstone RA (2011) Communal defense of territories and the evolution of sociality. Am Nat 178:787–800
Powell GVN (1985) Sociobiology and adaptive significance of interspecific foraging flocks in the Neotropics. Ornithol Monogr 36:713–732
Pulliam HR (1973) On the advantages of flocking. J Theor Biol 38:419–422
Remsen JV Jr, Robinson SK (1990) A classification scheme for birds in terrestrial foraging habitats. Stud Avian Biol 13:144–160
Ridley AR, Raihani NJ (2007) Facultative response to a kleptoparasite by the cooperatively breeding pied babbler. Behav Ecol 18:324–330
Satischandra SHK, Kudavidanage EP, Kotagama SW, Goodale E (2007) The benefits of joining mixed-species flocks for greater racket-tailed drongos Dicrurus paradiseus. Forktail 23:145–148
Shelley EL, Tanaka M, Ratnathicam AR, Blumstein DT (2004) Can Lanchester’s laws help explain inter-specific dominance in birds? Condor 106:395–400
Sridhar H, Beauchamp G, Shanker K (2009) Why do birds participate in mixed-species foraging flocks? A large-scale synthesis. Anim Behav 78:337–347
Sridhar H, Srinivasan U, Askins RA, Canales-Delgadillo JC, Chen CC et al (2012) Positive relationships between association strength and phenotypic similarity structure the assembly of mixed-species bird flocks worldwide. Am Nat 180:777–790
Sridhar H, Jordán F, Shanker K (2013) Species importance in a heterospecific foraging association network. Oikos 122:1325–1334
Srinivasan U, Quader S (2012) To eat and not be eaten: modeling resources and safety in multi-species animal groups. PLoS ONE 7:e42071
Srinivasan U, Raza RH, Quader S (2010) The nuclear question: rethinking species importance in multi-species animal groups. J Anim Ecol 79:948–954
Stotz DF (1993) Geographic variation in species composition of mixed species flocks in lowland humid forests in Brazil. Pap Avul Zool 38:61–75
Styring AR, Ickes K (2001) Woodpecker participation in mixed species flocks in peninsular Malaysia. Wilson Bull 113:342–345
Sullivan KA (1984) Information exploitation by downy woodpeckers in mixed-species flocks. Behaviour 91:294–311
Swynnerton CFM (1915) Mixed bird-parties. Ibis 57:346–354
Székely T, Szép T, Juhász T (1989) Mixed species flocking of tits (Parus spp.): a field experiment. Oecologia 78:490–495
Thiollay J-M, Jullien M (1998) Flocking behaviour of foraging birds in a neotropical rain forest and the antipredator defence hypothesis. Ibis 140:382–394
Veena T, Lokesha R (1993) Association of drongos with myna flocks: are drongos benefited? J Biosci 18:111–119
Waite TA, Grubb TC Jr (1988) Copying of foraging locations in mixed-species flocks of temperate-deciduous woodland birds: an experimental study. Condor 90:132–140
Whitehead H (2008) Analyzing animal societies. The University of Chicago Press, Chicago
Acknowledgments
We thank the Ministry of Environment and Forests, Government of India and the International Foundation for Science (grant no. D/4910-1) for supporting this study and the Karnataka Forest Department for providing permits for field work. We thank Guy Beauchamp, Kavita Isvaran, and Maria Thaker for discussions regarding sampling design and analysis and comments on earlier manuscript drafts. HS thanks Prakash, Nagesh, Narayan, Chandrakant, and Sadanand for their help with field work.
Ethical standards
This study complies with the current laws of India, where the study was carried out.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by P. A. Bednekoff
Electronic supplementary material
Below is the link to the electronic supplementary material.
Online Resource 1
(PDF 81.7 kb)
Online Resource 2
(PDF 109 kb)
Online Resource 3
(PDF 28.2 kb)
Online Resource 4
(PDF 22.7 kb)
Online Resource 5
(PDF 31.0 kb)
Online Resource 6
(PDF 39.2 kb)
Rights and permissions
About this article
Cite this article
Sridhar, H., Shanker, K. Using intra-flock association patterns to understand why birds participate in mixed-species foraging flocks in terrestrial habitats. Behav Ecol Sociobiol 68, 185–196 (2014). https://doi.org/10.1007/s00265-013-1633-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-013-1633-3