
Abstract
Under conditions of spatial and/or temporal variability in predation risk, prey organisms often rely on acquired predator recognition to balance the trade-offs between energy intake and risk avoidance. The question of ‘for how long’ should prey retain this learned information is poorly understood. Here, we test the hypothesis that the growth rate experienced by prey should influence the length of the ‘memory window’. In a series of laboratory experiments, we manipulated growth rate of juvenile rainbow trout and conditioned them to recognize a novel predator cue. We subsequently tested for learned recognition either 24 h or 8 days post-conditioning. Our results suggest that trout with high versus low growth rates did not differ in their response to learned predator cues when tested 24 h post-conditioning. However, trout on a high growth rate exhibited no response to the predator cues after 8 days (i.e. did not retain the recognition of the predator odour), whereas trout on a lower growth rate retained a strong recognition of the predator. Trout that differed in their growth rate only after conditioning did not differ in their patterns of retention, demonstrating growth rate after learning does not influence retention. Trout of different initial sizes fed a similar diet (percent body mass per day) showed no difference in retention of the predator cue. Together, these data suggest that growth rate at the time of conditioning determines the ‘memory window’ of trout. The implications for threat-sensitive predator avoidance models are described.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The ability of prey to recognize potential predators is key to individual survival. However, simply responding to any local threat may not represent an optimal strategy (Lima and Dill 1990), as predation pressure is known to be spatially and temporally variable (Griffin 2004; Lima and Steury 2005; Ferrari et al. 2009). A wide variety of taxonomically diverse prey species rely, therefore, on associative learning (acquired predator recognition) to assess the risk associated with potential predators (Brown 2003; Griffin 2004). Learned, versus ‘innate’, predator recognition allows prey to make dynamic adjustments to predation threats and to balance the conflicting pressures of predator avoidance and energy intake (i.e. threat-sensitive learning; Ferrari et al. 2005; Ferrari and Chivers 2006; Gonzalo et al. 2010). Within aquatic ecosystems, such predator recognition learning is often facilitated through the pairing of damage released chemical alarm cues (Chivers and Smith 1998) with the sight or smell of a novel predator (Brown 2003). Acquired predator recognition has been shown to increase probability of survival during staged encounters with live predators (Mirza and Chivers 2000; Darwish et al. 2005; Eiben and Persons 2007; Shier and Owings 2007).
Given that predation is indeed variable, a relevant, yet poorly understood question is how long should prey exhibit (i.e. retain) an overt response to acquired information? Following a single pairing of an alarm cue and a novel predator odour, hatchery-reared rainbow trout (Oncorhynchus mykiss) retain an overt learned response for up to 21 days (Brown and Smith 1998), though the response begins to decline rapidly after 10 days (Mirza and Chivers 2000). Likewise, Iberian green frog tadpoles (Pelophylax perezi) retain a learned response for up to 9 days following a single conditioning event (Gonzalo et al. 2009). Presumably, prey should only respond to learned predator cues as long as they represent an actual threat (Kotler 1992; Gonzalo et al. 2009).
Within the context of foraging decisions, several models have addressed the issue of retention of learned responses (McNamara and Houston 1987; Mangel 1990; Hirvonen et al. 1999). These models generally predict that there should exist a ‘memory retrieval’ window (Shettleworth and Plowright 1992; Grubb and Pravosudov 1994; Dunlap et al. 2009) that allows for a flexible response pattern. Under relatively constant environmental conditions, information regarding foraging decisions should be retained for a longer period (i.e. remain within the memory window), whereas under highly variable environmental conditions, older learned foraging information would be of lower value, and hence be ‘forgotten’ (i.e. fall outside this window) more quickly (Kotler 1992; Kraemer and Golding 1997; Kerr and Feldman 2003). Thus, such models predict that learned information should only be retained as long as it is relevant (Pravosudov and Clayton 2002; Brydges et al. 2008). Learned information that is no longer relevant is forgotten (i.e. no longer capable of eliciting a behavioural response).
Recently, Ferrari et al. (2010) have developed an analogous model for the learned response to predation threats. They suggest that a suite of extrinsic (i.e. predator community diversity and predator encounter rates) and intrinsic (i.e. morphological adaptations to predators and life history stage) factors should influence the duration of an individual prey's memory window. One such factor is growth. As prey grow, they may escape gape limits of potential predators (Nilsson and Brönmark 2000; Feary et al. 2009), develop increased escape abilities (Bishop and Brown 1992) or reduce the rate of encounter with predators by shifting habitat use (Werner et al. 1983). Thus, as prey grow, the value of learned information regarding predator identity should decrease. Accordingly, we could predict that higher growth rates would lead to a more rapid decline in the value of this information, thus faster-growing prey should ‘forget’ sooner than slower-growing prey. As such, growth may be predicted to influence the retention of overt behavioural responses to learned predator recognition. Alternatively, size may play a similar role in the shaping of the response to learned predator cues.
In a series of laboratory trials, we tested the hypothesis that growth rate will shape the ‘memory window’ of juvenile rainbow trout conditioned to recognize a novel predator odour. We predicted that trout fed a high versus low food ration (hence high vs. low growth rate) would exhibit similar responses when tested for learned recognition 24 h post-conditioning due to the high degree of relevance of recently acquired information. However, when tested 8 days post-conditioning, we predict that trout fed the high food ration should show a reduced (or absent) response to the acquired cue when compared to those fed the low food ration. In addition, we conducted companion experiments to test if growth post-conditioning (experiments 2 and 3) or absolute size (experiment 4) had an effect on the retention of learned predator recognition.
General methods
Test fish
Juvenile rainbow trout were obtained from a commercial supplier (Pisciculture Arpents des Vert, Ste Edwidge-de-Clifton, Quebec) and transported to the laboratory. Trout were the offspring of brood stock reared in the hatchery for between two to four generations, crossed with wild-caught adults. Prior to testing, trout were housed in 390 l recirculating holding tanks (∼18°C, pH ∼ 7.0) under a 14:10 L:D cycle and fed ad libitum daily with commercial trout chow (Corey Mills). Adult pumpkinseed (Lepomis cyanellus), used as predator odour donors, were collected from Canal Lachine, Montreal, Quebec. Pumpkinseeds were held under in similar tanks and fed a diet of brine shrimp and commercial cichlid pellets daily. Under laboratory conditions, juvenile rainbow trout typically retain the learned response to a novel predator odour for up to 3 weeks (Brown and Smith 1998), though the response begins to wane after approximately 10 to 14 days (Mirza and Chivers 2000). Thus, we chose to test either 24 h or 8 days post-conditioning to ensure that we would be able to detect a learned response if present.
Stimulus production
We generated trout alarm cues from 15 juvenile trout (mean ± SD fork length = 5.60 ± 0.41 cm). Donor trout were killed via cervical dislocation (in accordance with Concordia University Animal Research Ethics Committee protocol number AREC-2008-BROW), and skin fillets were removed from either side of the donors and immediately placed into chilled distilled water. Skin fillets were then homogenized, filtered through polyester filter floss and diluted to the desired volume with the addition of distilled water. We collected a total of 108 cm2 of skin (in a final volume of 1,080 ml of distilled water). Chemical alarm cues were frozen in 20 ml aliquots at −20°C until needed. As a control, we also froze 20 ml samples of distilled water.
In order to collect the odour of a novel predator, we placed four pumpkinseeds (11.20–14.5 cm S.L.) into unfiltered 37-L glass aquaria, filled with 15 l of dechlorinated tap water for 3 days. Pumpkinseeds were not fed during this period. For at least 1 week prior to odour collection, we fed pumpkinseed only brine shrimp to ensure that the resulting predator odour did not contain any diet-related cues that may have been recognized by the test fish. After the 3-day period, the water from each donor tank was pooled, filtered and frozen in 50 ml aliquots at −20°C until needed.
Conditioning phase
For each of the four experiments (see below), we placed groups of ten juvenile trout into each of four ‘conditioning’ tanks. Conditioning tanks consisted of a series of 60-l glass aquaria, each equipped with a power filter and a gravel substrate (∼18°C, pH ∼ 7.0). Trout were placed in the tanks either 8 days (experiments 1, 3 and 4) or 1 day (experiment 2) prior to conditioning. To condition trout to recognize a novel predator odour, we introduced 20 ml of predator odour paired with 20 ml of trout alarm cue. As a control, we ‘pseudo-conditioned’ equal numbers of trout by introducing 20 ml of distilled water paired with 20 ml of predator odour. In all cases, we turned the power filter off approximately 10 min prior to conditioning and left it off for 1 h following conditioning. Conditioning occurred between 10:00 and 11:00. The day of conditioning (or pseudo-conditioning controls) is considered day 1 of the experiment.
Recognition phase
Approximately 6 h after conditioning, we transferred four or five individual trout from each of the four conditioning tanks (i.e. each of the treatment combinations) to test aquaria for recognition testing on day 2. The remaining trout were fed on the appropriate diet until day 8 and then transferred to individual test tanks for recognition testing on day 9. Test tanks consisted of a series of 37-l glass aquaria, each equipped with a single airstone and a gravel substrate. Test tanks were not filtered. In addition, we attached a 2-m length of plastic tubing to the back wall of the test tanks to allow for the introduction experimental cues without disturbing test fish.
Recognition trials consisted of a 5-min pre-stimulus and a 5-min post-stimulus observation period. Prior to the pre-stimulus observation, we withdrew and discarded 60 ml of tank water through the plastic tubing. We then withdrew and retained an additional 60 ml of water. Immediately following the pre-stimulus observation period, we injected 10 ml of predator odour and slowly flushed it into the tank with the retained water. During both the pre- and post-stimulus observation periods, we recorded the time spent moving and the frequency of foraging attempts. We fed test fish approximately 30 min prior to testing, to ensure activity, with a standard amount of powdered trout chow (∼0.1 g of powdered chow). Sufficient food was presented such that there were enough food particles remaining during the behavioural observations to quantify foraging without the need for additional food (Vavrek et al. 2008). A feeding attempt was defined as a pecking movement towards a food particle, either on the substrate or in the water column. A reduction in movement and foraging rate are consistent with increased predator avoidance behaviour in juvenile salmonids (Martel and Dill 1993; Brown and Smith 1997; Leduc et al. 2009). In all cases, the observers were blind to the treatment.
Experiment 1: different food rations before and after conditioning
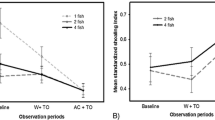
In order to test the effects of growth on the strength of conditioning and expression of learned recognition, we fed groups of trout either 1% or 5% of mean group body mass per day for a period of 7 days prior to conditioning. For trout tested on day 9, we continued to feed the same relative diet, adjusted for the reduced number of fish remaining in the conditioning tanks. We chose these diets because 1% represents a minimum growth food ration and 5% represents an abundance of food, allowing for high relative growth (Alsop and Wood 1997; Kamunde and Wood 2003; Brown et al. 2009a). Following testing on day 2 or day 9, individuals were blotted on sterile paper and measured (mass to the nearest 0.001 g and standard length to the nearest 0.5 mm). Each conditioning block yielded four test fish per treatment combination. We tested four complete blocks (n = 4 per conditioning stimulus, for a total of 16 trout per treatment combination (N = 128)). Mean (±SD) fork length was 3.25 ± 0.31 cm (see Fig. 1a for mean mass).

Mean (±SE) mass of juvenile rainbow trout tested in each of the four experiments. Experiment 1 (a): trout were fed either 1% or 5% mean body mass per day for the duration of the study. Experiment 2 (b): trout were fed 1% or 5% from the time of conditioning (day 1) with no pre-conditioning feeding phase. Experiment 3 (c): trout were fed 1% or 5% for 1 week prior to conditioning and then fed only 1% until testing on day 9. Experiment 4 (d): trout were fed similar (1%) diets, but differed in initial mass
We calculated the change in time spent moving and foraging attempts (post-stimulus − pre-stimulus) and used these difference scores as dependent variables in all analyses. We compared pre-stimulus time moving and foraging rates between the 1% and 5% food ration treatments for trout tested on day 2 (MANOVA: F 2,61 = 0.78, P = 0.46) and day 9 (MANOVA: F 2,61 = 1.97, P = 0.15) and found no difference in baseline activity, justifying the use of difference scores as dependent variables. We tested for the overall effects of conditioning stimulus (alarm cue versus distilled water), diet (1% versus 5%) and recognition test day (day 2 versus day 9) using univariate GLM ANOVAs. To account for any effect of ‘conditioning block’, we included this as a random variable. Given the significant overall interaction between the three independent variables (see below), we tested for the effects of conditioning stimulus and diet for day 2 and day 9 separately. Data met the assumptions for parametric analysis (i.e. were normally distributed and homoscedastic).
Experiment 2: different food rations after conditioning
In order to test for the possibility that the observed results of experiment 1 (see below) were due to growth after conditioning, rather than differences in growth prior to conditioning, we conducted this experiment. We placed groups of ten trout into the conditioning tanks 24 h prior to conditioning, with no prior differences in food ration. Following conditioning, trout were fed either 1% or 5% diets as in experiment 1. Sample sizes were the same as experiment 1. Mean (±SD) fork length at time of testing was 3.42 ± 0.32 cm (Fig. 1b for mean mass). Data met the assumptions of parametric tests and were analyzed as above. As in experiment 1, we found no difference in baseline activity levels (MANOVA: F 2,61 = 0.94, P = 0.39 and F 2,61 = 0.26, P = 0.77, days 2 and 9, respectively).
Experiment 3: different food rations prior to conditioning
As a further control experiment, we fed groups of trout a diet of 1% or 5% mean body mass for a period of 7 days prior to conditioning. Following conditioning, trout were either tested on day 2 or fed a common diet of 1% mean body mass per day for a further 8 days. We tested a total of four blocks (n = 5 per block, N = 160). Mean (±SD) fork length at time of testing was 3.15 ± 0.28 cm (Fig. 1c for mean mass). Data met the assumptions of parametric tests and were analyzed as above. As in experiment 1, there was no difference in baseline activity (MANOVA: F 2,77 = 2.37, P = 0.10 and F 2,77 = 1.52, P = 0.23, days 2 and 9, respectively).
Experiment 4: different absolute size, same food rations
In order to test the possibility that absolute size, rather than growth rate per se, is driving our observed results, we tested juvenile rainbow trout of differing size under similar conditions to experiment 1. We tested small (3.60 ± 0.34 cm) and large (5.71 ± 0.22 cm) trout (Fig. 1d for mean mass). We fed trout in the conditioning tanks a diet of 1% mean body mass per day for a period of 7 days prior to conditioning. Trout were conditioned as in experiment 1, and those to be tested on day 9, we continued the 1% mean body mass per day. We tested a total of four blocks (n = 4 per block per treatment combination, N = 128). Data met the assumptions of parametric tests and were analyzed as above. Though there was a trend towards smaller trout being more active than larger trout, baseline activity was not different for trout tested on day 2 (MANOVA: F 2,61 = 3.05, P = 0.052) nor day 9 (F 2,61 = 2.97, P = 0.058).
Results
Experiment 1
Our initial analysis revealed no effect of the blocking variable for either the change in time spent moving or foraging attempts (F 3,117 = 1.51, P = 0.217 and F 3,117 = 1.34, P = 0.26, respectively). We found significant three-way interactions between conditioning stimulus, diet and recognition test day for both change in time moving (F 4,117 = 3.92, P = 0.005) and foraging response variables (F 1,117 = 2.51, P = 0.046; Fig. 2). When testing for the effects of conditioning stimulus and diet for day 2 recognition trials alone, we found significant effects of conditioning stimulus for both change in time spent moving (F 1,57 = 36.42, P < 0.001) and change in foraging attempts (F 1,57 = 18.53, P < 0.001), but no effect of diet for either behavioural measure (change in time moving: F 1,57 = 0.11, P = 0.74; change in foraging attempts: F 1,57 = 0.15, P = 0.70). Moreover, there was no significant two-way interaction for either change in time moving (F 1,57 = 3.47, P = 0.51) or change in foraging attempts (F 1,57 = 0.65, P = 0.42; Fig. 2). Trout fed on both high and low food diets exhibited similar learned responses to the predator odour.

Mean (±SE) change in time spent moving (a) and foraging attempts (b) for trout tested in experiment 1. Dark bars denote trout initially conditioned with alarm cue + predator odour, and open bars denote trout initially conditioned with distilled water + predator odour. Trout fed on high food (5%) or low food (1%) diets throughout the course of the experiment were tested for recognition of the predator odour 24 h post-conditioning (day 2) or 8 days post-conditioning (day 9). N = 16 for each treatment combination
However, we found a considerably different response pattern when testing for expression of learned predator odours on day 9. We found significant conditioning stimulus × diet interactions for change in time moving (F 1,57 = 6.05, P = 0.017) and change in foraging attempts (F 1,57 = 4.42, P = 0.04), demonstrating that only the trout fed the low food regime (1% mean body mass per day), and not the high food regime, exhibited an overt antipredator response on day 9 (Fig. 2).
Experiment 2
When trout did not experience different feeding regimes prior to conditioning, we found no evidence that diet post-conditioning (1% vs. 5%) had an effect on the expression of acquired predator recognition. Trout fed either the high versus low food diet following conditioning exhibited similar responses to the learned predator odour on both day 2 and day 9 of recognition testing. We found a significant effect of conditioning stimulus for both change in time spent moving (F 1,117 = 22.86, P < 0.001) and change in foraging attempts (F 1,117 = 21.94, P < 0.001; Fig. 3), but no effect of diet (change in time moving: F 1,117 = 0.05, P = 0.82; change in foraging attempts, F 1,117 = 0.81, P = 0.37) or recognition test day (change in time moving: F 1,117 = 0.004, P = 0.95; change in foraging attempts: F 1,117 = 0.07, P = 0.78). Moreover, there was no three-way interaction (change in time moving: F 4,117 = 0.16, P = 0.96; change in foraging attempts: F 4,117 = 0.45, P = 0.78). As with experiment 1, there was no effect of the blocking variable for either behavioural measure (change in time moving: F 3,117 = 0.65, P = 0.56; change in foraging attempts: F 3,117 = 1.79, P = 0.15).

Mean (±SE) change in time spent moving (a) and foraging attempts (b) for trout tested in experiment 2. Dark bars denote trout initially conditioned with alarm cue + predator odour, and open bars denote trout initially conditioned with distilled water + predator odour. Trout were fed high food (5%) or low food (1%) diets from day 1 (conditioning) onwards and tested for recognition of the predator odour 24 h post-conditioning (day 2) or 8 days post-conditioning (day 9). N = 16 for each treatment combination
Experiment 3
When trout were fed different feeding regimes for 1 week prior to conditioning and then switched to the same proportional regime (1% mean body mass per day), the results were strikingly similar to those of experiment 1. For the overall analyses, we found significant interactions among conditioning stimulus, diet and recognition test day for both change in time moving (F 4,149 = 2.47, P = 0.047) and the change in foraging attempts (F 4,149 = 2.52, P = 0.044; Fig. 4). When we separated the dataset based on recognition test day (as in experiment 1), we found no evidence of an interaction between conditioning stimulus and diet on the recognition of predator cues on day 2 (change in time moving: F 1,73 = 0.43, P = 0.51; change in foraging attempts: F 1,73 = 0.014, P = 0.90), but we did find a significant interaction for both change in time moving (F 1,73 = 4.03, P = 0.048) and change in foraging attempts (F 1,73 = 5.90, P = 0.018; Fig. 4) on day 9. As in experiment 1, there was no overall effect of the blocking variable for either change in time moving (F 3,149 = 0.64, P = 0.59) or change in foraging attempts (F 3,149 = 0.41, P = 0.74). This finding further supports the hypothesis that growth rate at the time of conditioning influences expression of learned information.

Mean (±SE) change in time spent moving (a) and foraging attempts (b) for trout tested in experiment 3. Dark bars denote trout initially conditioned with alarm cue + predator odour, and open bars denote trout initially conditioned with distilled water + predator odour. Trout were fed high food (5%) or low food (1%) diets for 1 week prior to conditioning; after which, all trout to be tested on day 9 were fed the low food ration. Trout were tested for the recognition of the predator odour 24 h post-conditioning (day 2) or 8 days post-conditioning (day 9). N = 20 for each treatment combination
Experiment 4
When trout of different initial sizes were fed proportionally the same diet (1% mean body mass per day), we found a significant effect of conditioning stimulus on the learned recognition of the novel predator odour (change in time moving: F 1,117 = 21.16, P < 0.001; change in foraging attempts: F 1,117 = 38.16, P < 0.001; Fig. 5). There were no significant effects of size class (change in time moving: F 1,117 = 0.06, P = 0.80; change in foraging attempts: F 1,117 = 0.08, P = 0.77), day of recognition testing (change in time moving: F 1,117 = 0.25, P = 0.62; change in foraging attempts: F 1,117 = 0.08, P = 0.77) nor an interaction among the three (change in time moving: F 4,117 = 0.14, P = 0.97; change in foraging attempts: F 4,117 = 0.15, P = 0.98; Fig. 5). As above, there was no significant effect of the blocking variable (change in time moving: F 3,117 = 2.20, P = 0.09; change in foraging attempts: F 3,117 = 2.15, P = 0.10). These results suggest that absolute size, at least within this size range, cannot explain the results of experiment 1.

Mean (±SE) change in time spent moving (a) and foraging attempts (b) for trout tested in experiment 4. Dark bars denote trout initially conditioned with alarm cue + predator odour, and open bars denote trout initially conditioned with distilled water + predator odour. Trout differed in initial mass and were fed a similar diet (1%) throughout the course of the experiment. Trout were tested for the recognition of the predator odour 24 h post-conditioning (day 2) or 8 days post-conditioning (day 9). N = 16 for each treatment combination
Discussion
Recent models concerning the retention of ecologically relevant learning predict that the ‘value’ of information should decrease over time and that a suite of factors, including growth rate, will influence this rate of information loss (McNamara and Houston 1987; Mangel 1990; Hirvonen et al. 1999; Ferrari et al. 2010). Our current study provides support for the prediction that an individual's rate of growth at the time it acquires new information will influence its retention. The results of experiment 1 suggest that growth rate at the time of conditioning influences the ‘memory window’ for juvenile rainbow trout. Trout fed the higher food ration (hence higher growth rate) did not differ from those fed the lower food ration when tested on day 2, demonstrating that growth rate does not influence the strength of the learned response. However, when tested on day 9, only those fed the lower food ration exhibited expression of the response to the predator odour; trout fed the high food ration did not differ from the pseudo-conditioned controls. Alternatively, we might have predicted that energy costs associated with retention may be a significant factor shaping the observed memory window (i.e. Fitting et al. 2008). If this were the case, however, we would expect trout on the lower growth trajectory to show reduced retention. While it is tempting to state that our trout fed the high food ration ‘forgot’ sooner than those fed the low food ration, we can not differentiate between the neurological loss of recognition (Speed 2000; Wixted 2004) and the possibility that trout recognized the predator odour but fail to respond due to a behavioural decision. Indeed, the absence of an overt response does not mean that the memory of learned information is not present (Kraemer and Golding 1997; Skow and Jakob 2006). Regardless of the underlying mechanism, differences in the expression of acquired predator recognition will likely have considerable functional consequences.
Ferrari et al. (unpublished data) have found similar results with woodfrog tadpoles (Rana sylvatica). Tadpoles with higher growth rates exhibited reduced retention compared to tadpoles with lower growth rates. However, their study suggests that growth rate at the time of conditioning and following conditioning both contribute to shape retention. This differs from our current study since the results of experiments 2 and 3 suggest that growth after the initial conditioning phase does not have any detectable effect on the expression of an overt response to the predator cue. Alternatively, absolute size and not growth rate per se may influence the expression of predator recognition. At the time of conditioning (experiment 1), trout on the high food ration were considerably larger than those on the low food ration (0.85 ± 0.03 versus 0.66 ± 0.03 g). This, however, is an unlikely explanation given the results of experiment 4. When fed a similar diet (percent body mass per day), large and small trout exhibited similar retention patterns. Moreover, it could be argued that low body condition is linked to reduced retention. If this were the case, then we would have expected to see the trout on the lower growth trajectory (presumably lower condition) to have lower retention. Taken together, these results provide strong evidence that growth rate at the time of conditioning fixes the memory window of learned predator recognition.
Regardless of the underlying mechanism, it is clear that factors that increase the value of learned information should extend the ‘memory window’, while factors that reduce its relative value should result in the loss of response. The model of a flexible ‘memory window’ predicts that learned or acquired information should be retained (remain within the window) only as long as it is relevant (Cuthill et al. 1990; Mackney and Hughes 1995; Kraemer and Golding 1997). Moreover, factors that reinforce the relevance of information, either positively or negatively, will influence how long they remain within the window (Kerr and Feldman 2003; Dunlap et al. 2009). For example, retention of learned foraging information such as food caches increases as the predictability of finding food decreases (Cuthill et al. 1990; Prasvosudov and Clayton 2001).
Under variable habitat conditions, responding to learned information should be considered adaptive, as it would increase flexibility in an individual's behavioural repertoire (Kerr and Feldman 2003; Griffin 2004; Lima and Steury 2005). Recent models (White 2001; Dunlap et al. 2009) suggest that as an individual's ‘cost of living’ varies, the duration that learned information remains relevant (i.e. remains within the memory window) should likewise vary. Factors such as variability in abundance or predictability in finding suitable food sources and/or ambient predation pressure should be expected to influence an individual's ‘cost of living’. As the cost of living increases, the memory window for acquired information should become longer as learned information remains relevant (Kotler 1992; Kerr and Feldman 2003; Dunlap et al. 2009). Conversely, reduced cost of living should result in extended memory windows (Dunlap et al 2009).
Presumably, factors such as reduced food availability or predictability and increased predation pressure should result in increased pressure on individuals to balance threat-sensitive trade-offs (Helfman 1989; Brown et al. 2006; 2009b; Roitberg et al. 2010). As such, our observed differential response to learned predator recognition may result in behavioural response patterns consistent with threat-sensitive trade-offs. When applied to the question of retention of learned predator recognition, conditions leading to high growth rates (high availability and/or predictability of food) likely translate to a relatively low cost of living (Kerr and Feldman 2003; but see Biro and Stamps 2008; Adriaenssens and Johnsson 2011). At the same time, high growth should lead to a reduced individual risk of predation due to greater ability to escape predator (Bishop and Brown 1992), reduced risk of gape-limited predators (Nilsson and Brönmark 2000; Feary et al. 2009) and/or a higher energy reserve (Reinhardt and Healy 1999). As such, the retention period should be expected to decrease (i.e. prey ‘forget’ sooner). Conversely, low growth rates and the increased risk of predation associated with smaller body sizes and/or reduced energy stores (Reinhardt and Healy 1999) may result in elevated costs of living for prey individuals, leading to increased retention of acquired information (Dunlap et al. 2009). Overall, any devaluation of learned information may manifest as differing behavioural strategies, with faster-growing trout exhibiting a risk prone response pattern, while slower growing trout showing a more risk aversive pattern.
Recent research shows that threat-sensitive trade-offs may shape the intensity of learning in prey organisms (Ferrari et al. 2005; Ferrari and Chivers 2006; Zhao et al. 2006; Gonzalo et al. 2010). These studies demonstrate that the intensity of the learned response to a novel predator cue is proportional to the strength of the initial conditioning. Such a learning mechanism would allow prey to respond to acquired cues with a response intensity proportional to the initial level of perceived risk. Our current study builds on these results, suggesting that threat-sensitive trade-offs after the initial conditioning event may also shape the response intensity. In each of the current studies, the initial strength of the experimental conditioning phases (alarm cue + predator odour) was the same. Thus, it is not surprising that we found no difference between the diet (size) treatments when trout were tested on day 2. Rather, the observed retention effects (experiments 1 and 3) on day 9 likely represent the product of threat-sensitive trade-offs at the time of recognition testing and not due to differences in the initial conditioning events.
The applied goal of predator recognition training, at least for salmonids, has always been to condition hatchery-reared fish prior to stocking in hopes of increasing post-stocking survival (Berejikian et al. 1999; Brown and Smith 1998; Brown and Laland 2001). There is some support that this methodology works to increase survival (Berejikian et al. 1999; Mirza and Chivers 2000). However, recent studies by Wisenden et al. (2004) and Hawkins et al. (2007) suggest that conditioned hatchery-reared fish do not exhibit any evidence of learned recognition of predator cues when tested under fully natural conditions. Our current results suggest that one possible explanation for these findings is that hatchery fish, which presumably would be fed on a high growth rate diet, are simply not showing the expression of learned information due to diet-related threat-sensitive trade-offs. An intriguing possibility would be to test the effects of dietary restriction prior to conditioning and subsequent release. Future studies should address this possibility.
References
Adriaenssens B, Johnsson JI (2011) Shy trout grow faster: exploring links between personality and fitness-related traits in the wild. Behav Ecol. doi:10.1093/beheco/arq185
Alsop DH, Wood CM (1997) The interactive effects of feeding and exercise on oxygen consumption, swimming performance and protein usage in juvenile rainbow trout (Oncorhynchus mykiss). J Exp Biol 200:2337–2346
Berejikian BA, Smith RJF, Tezak PE, Schroder SL, Knusden CM (1999) Chemical alarm signals and complex hatchery rearing habitats affect antipredator and survival of chinook salmon (Oncorhynchus tshawytscha) juveniles. Can J Fish Aquat Sci 56:830–838
Biro PA, Stamps JA (2008) Are animal personality traits linked to life-history productivity? Trends Ecol Evol 23:361–368
Bishop TD, Brown JA (1992) Threat-sensitive foraging by larval threespine sticklebacks (Gasterosteus aculeatus). Behav Ecol Sociobiol 31:133–138
Brown GE (2003) Learning about danger: chemical alarm cues and local risk assessment in prey fishes. Fish Fish 4:227–234
Brown C, Laland KN (2001) Social learning and life skills training for hatchery reared fish. J Fish Biol 59:471–493
Brown GE, Smith RJF (1997) Conspecific skin extracts elicit antipredator responses in juvenile rainbow trout (Oncorhynchus mykiss). Can J Zool 75:1916–1922
Brown GE, Smith RJF (1998) Acquired predator recognition in juvenile rainbow trout (Oncorhynchus mykiss): conditioning hatchery-reared fish to recognize chemical cues of a predator. Can J Fish Aquat Sci 55:611–617
Brown GE, Bongiorno T, DiCapua DM, Ivan LI, Roh E (2006) Effects of group size on the threat-sensitive response to varying concentrations of chemical alarm cues by juvenile convict cichlids. Can J Zool 84:1–8
Brown GE, Macnaughton CJ, Elvidge CK, Ramnarine I, Godin J-GJ (2009a) Provenance and threat-sensitive predator avoidance patterns in wild-caught Trinidadian guppies. Behav Ecol Sociobiol 63:699–706
Brown GE, Harvey MC, Leduc AOHC, Ferrari MCO, Chivers DP (2009b) Social context, competitive interactions and the dynamic nature of antipredator responses of juvenile rainbow trout. J Fish Biol 79:552–562
Brydges NM, Heathcote RJP, Braithwaite VA (2008) Habitat stability and predation pressure influence learning and memory in populations of three-spined sticklebacks. Anim Behav 75:935–942
Chivers DP, Smith RJF (1998) Chemical alarm signaling in aquatic predator-prey systems: a review and prospectus. Écoscience 5:338–352
Cuthill IC, Kacelnik A, Krebs JR, Haccou P, Iwasa Y (1990) Starlings exploiting patches: the effect of recent experience on foraging decisions. Anim Behav 40:625–640
Darwish TL, Mirza RS, Leduc AOHC, Brown GE (2005) Acquired recognition of novel predator odour cocktails by juvenile glowlight tetras. Anim Behav 70:83–89
Dunlap AS, McLinn CM, MacCormick HA, Scott ME, Kerr B (2009) Why some memories do not last a lifetime: dynamic long-term retrieval in changing environments. Behav Ecol 20:1096–1105
Eiben B, Persons M (2007) The effect of prior exposure to predator cues on chemically-mediated defensive behavior and survival in the wolf spider Rabidosa rabida (Aranae: Lycosidae). Behaviour 144:889–906
Feary DA, McCormick MI, Jones GP (2009) Growth of reef fishes in response to live coral cover. J Exp Mar Biol Ecol 373:45–49
Ferrari MCO, Chivers DP (2006) Learning threat-sensitive predator avoidance: how do fathead minnows incorporate conflicting information? Anim Behav 71:19–26
Ferrari MCO, Trowell JJ, Brown GE, Chivers DP (2005) The role of learning in the development of threat-sensitive predator avoidance by fathead minnows. Anim Behav 70:777–784
Ferrari MCO, Sih A, Chivers DP (2009) The paradox of risk allocation: a review and prospectus. Anim Behav 78:579–585
Ferrari MCO, Brown GE, Bortolotti GR, Chivers DP (2010) Linking predator risk and uncertainty to adaptive forgetting: a theoretical framework and empirical test using tadpoles. Proc R Soc Lond B 277:2205–2210
Fitting S, Booze RM, Gilbert CA, Mactutus CF (2008) Effects of chronic adult dietary restriction on spatial learning in the aged F344 x BN hybrid F1 rat. Physiol Behav 93:560–569
Gonzalo A, López P, Martín J (2009) Learning, memory and apparent forgetting of chemical cues from new predators by Iberian green frog tadpoles. Anim Cogn 12:745–750
Gonzalo A, López P, Martin J (2010) Risk level of chemical cues determines retention of recognition of new predators in Iberian green frog tadpoles. Behav Ecol Sociobiol 64:1117–1123
Grubb TC Jr, Pravosudov VV (1994) Toward a general theory of energy management in wintering birds. J Avian Biol 25:255–260
Griffin AS (2004) Social learning about predators: a review and prospectus. Learn Behav 32:131–140
Hawkins LA, Armstrong JD, Magurran AE (2007) A test of how predator conditioning influence survival of hatchery-reared Atlantic salmon, Salmo salar, in restocking programmes. Fish Manage Ecol 14:291–293
Helfman GS (1989) Threat-sensitive predator avoidance in damselfish-trumpetfish interactions. Behav Ecol Sociobiol 24:47–58
Hirvonen H, Ranta E, Rita H, Peuhkuri N (1999) Significance of memory properties in prey choice decisions. Ecol Model 115:177–189
Kamunde C, Wood CM (2003) The influence of ration size on copper homeostasis during sublethal dietary copper exposure in juvenile rainbow trout, Oncorhynchus mykiss. Aquat Toxicol 62:235–254
Kerr B, Feldman MW (2003) Carving the cognitive niche: optimal learning strategies in homogeneous and heterogeneous environments. J Theor Biol 220:169–188
Kotler BP (1992) Behavioural resource depression and decaying perceived risk of predation in two species of coexisting gerbils. Behav Ecol Sociobiol 30:239–244
Kraemer PJ, Golding JM (1997) Adaptive forgetting in animals. Psych Bull Rev 4:480–491
Leduc AOHC, Roh E, Brown GE (2009) Effects of acid rainfall on juvenile Atlantic salmon (Salmo salar) antipredator behaviour: loss of chemical alarm function and potential survival consequences during predation. Mar Freshwater Res 60:1223–1230
Lima SL, Dill LM (1990) Behavioral decisions made under the risk of predation: a review and prospectus. Can J Zool 68:619–640
Lima SL, Steury TD (2005) Perception of predation risk: the foundation of nonlethal predator-prey interactions. In: Castellanos I, Barbosa P (eds) Ecology of predator-prey interactions. Oxford University Press, Oxford, pp 166–188
Mackney PA, Hughes RN (1995) Foraging behaviour and memory window in sticklebacks. Behaviour 132:1241–1253
Mangel M (1990) Dynamic information in uncertain and changing worlds. J Theor Biol 146:317–332
Martel G, Dill LM (1993) Feeding and aggressive behaviours in juvenile coho salmon (Oncorhynchus kisutch) under chemically-mediated risk of predation. Behav Ecol Sociobiol 32:365–370
McNamara JM, Houston AI (1987) Memory and the efficient use of information. J Theor Biol 125:385–395
Mirza RS, Chivers DP (2000) Predator-recognition training enhances survival of brook trout: evidence from laboratory and field-enclosure studies. Can J Zool 78:2198–2208
Nilsson PA, Brönmark C (2000) Prey vulnerability to a gape-limited predator: behavioural and morphological impacts on northern pike piscivory. Oikos 88:539–546
Pravosudov VV, Clayton NS (2001) Effects of demanding foraging conditions on cache retrieval accuracy in food-caching mountain chickadees (Poecile gambeli). Proc R Soc Lond B 268:363–368
Pravosudov VV, Clayton NS (2002) A test of the adaptive specialization hypothesis: population differences in caching, memory and the hippocampus in black-capped chickadees (Poecile atricapilla). Behav Neurosci 116:515–522
Reinhardt UG, Healey MC (1999) Season- and size-dependent risk taking in juvenile coho salmon: experimental evaluation of asset protection. Anim Behav 57:923–933
Roitberg BD, Zimmermann K, Hoffmeister TS (2010) Dynamic response to danger in a parasitoid wasp. Behav Ecol Sociobiol 64:627–637
Shettleworth SJ, Plowright CMS (1992) How pigeons estimate rates of prey encounter. J Exp Psych: Anim Behav Proc 18:219–235
Shier DM, Owings DH (2007) Effects of social learning on predator training and postrelease survival in juvenile black-tailed prairie dogs, Cynomys ludovicianus. Anim Behav 73:567–577
Skow CD, Jakob EM (2006) Jumping spiders attend to context during learned avoidance of aposematic prey. Behav Ecol 17:34–40
Speed MP (2000) Warning signals, receiver psychology and predator memory. Anim Behav 60:269–278
Vavrek MA, Elvidge CK, DeCaire R, Belland B, Jackson CD, Brown GE (2008) Disturbance cues in freshwater prey fishes: do juvenile convict cichlids and rainbow trout respond to ammonium as an ‘early warning’ signal? Chemoecol 18:255–261
Werner EE, Gilliam JF, Hall DJ, Mittlebach GG (1983) An experimental test of the effects of predation risk on habitat use in fish. Ecology 64:1540–1548
Wisenden BD, Klitzke J, Nelson R, Friedl D, Jacobson PC (2004) Predator-recognition training of hatchery-reared walleye (Stizostedion vitreum) and a field test of a training method using yellow perch (Perca favescens). Can J Fish Aquat Sci 61:2144–2150
White KG (2001) Forgetting functions. Anim Learn Behav 29:193–207
Wixted JT (2004) The psychology and neuroscience of forgetting. Ann Rev Psychol 55:235–269
Zhao X, Ferrari MCO, Chivers DP (2006) Threat-sensitive learning of predator odours by a prey fish. Behaviour 143:1103–1121
Acknowledgements
We wish to thank Drs. James Grant and Robert Weladji for comments on earlier versions of this manuscript and Matthew Murphy and Asra Toobie for assistance in the laboratory. All work reported herein was conducted in accordance with Concordia University Animal Research Ethics Protocol number AC-2008-BROW. Financial support was provided by Concordia University and the Natural Sciences and Engineering Research Council of Canada to GEB and the University of Saskatchewan and NSERC to DPC and an NSERC Postdoctoral Fellowship to MCOF.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. Krause
Rights and permissions
About this article
Cite this article
Brown, G.E., Ferrari, M.C.O., Malka, P.H. et al. Growth rate and retention of learned predator cues by juvenile rainbow trout: faster-growing fish forget sooner. Behav Ecol Sociobiol 65, 1267–1276 (2011). https://doi.org/10.1007/s00265-011-1140-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-011-1140-3