
Abstract
Courtship displays are often important in determining male mating success but can also be costly. Thus, instead of courting females indiscriminately, males might be expected to adjust their signalling effort strategically. Theory, however, predicts that such adjustments should depend on the rate with which males encounter females, a prediction that has been subject to very little empirical testing. Here, we investigate the effects of female encounter rate on male courtship intensity by manipulating the time interval between sequential presentations of large (high quality) and small (low quality) females in a fish, the Australian desert goby Chlamydogobius eremius. Males that were presented with a small female immediately after a large female reduced their courtship intensity significantly. However, males courted large and small females with equal intensity if the interval between the sequential presentations was longer. Our results suggest that mate encounter rate is an important factor shaping male reproductive decisions and, consequently, the evolutionary potential of sexual selection.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Males of many species perform conspicuous courtship displays in an attempt to increase their mating success. However, courtship can be costly in terms of, for example, time, energy and predation risk (e.g. Kotiaho et al. 1998; Woods et al. 2007; Hoefler et al. 2008). To reduce these costs, males may evolve strategies to adjust their level of courtship depending on the situation. For instance, males may court with lower intensity when females are likely to be aggressive, as in dwarf chameleons (Stuart-Fox and Whiting 2005), or prone to raiding the male's nest, as in three-spined sticklebacks (Belles-Isles et al. 1990). Importantly, males may also reduce the time and effort spent courting females of lower reproductive value as shown, for example, in fiddler crabs (Reading and Backwell 2007) and zebra finches (Jones et al. 2001). Theory predicts that such strategies can increase a male's reproductive success, but only when there is a good chance of rapidly finding alternative (higher-quality) females (Bonduriansky 2001). Thus, the rate with which males encounter females should be a crucial factor in determining whether males adjust their courtship effort with regard to female reproductive quality (Kokko and Johnstone 2002). Surprisingly few studies, however, have explicitly addressed whether mate encounter rate influences male courtship decisions (Wong et al. 2004).
The Australian desert goby, Chlamydogobius eremius, is a remarkable freshwater fish endemic to the arid regions of Central Australia. Despite the taxing environmental conditions of their desert habitat, males invest heavily in reproduction through nest defence, elaborate courtship displays and exclusive paternal care of the eggs (Allen et al. 2002). As in other gobies (Kvarnemo 1994; Pelabon et al. 2003), there is a strong positive length–fecundity relationship among female desert gobies (T. K. Lehtonen, L. Slingerland, P. A. Svensson and B. B. M. Wong, unpublished manuscript). Thus, by mating with larger females, males may receive a larger clutch, thereby increasing their reproductive success. Previously, Wong and Svensson (2009) showed that desert goby males adjust their courtship effort according to female size, and that this was true even when females were presented one at a time (i.e. sequentially). In that study, however, the interval between female encounters was kept constant, and it remains to be tested if and how the timing of female presentations affects male courtship tactics. This is an ecologically meaningful question because, in the wild, male desert gobies defend and guard nest sites and must rely on mate-searching females to visit their territories. Thus, nest-holding males have limited opportunities to directly affect the rate of mate encounters. However, males might be expected to respond to variation in female visits by adjusting their mating decisions according to the perceived encounter rate. The aim of the present study was to test whether the interval between female encounters influences how males adjust their courtship effort with regard to female quality.
Materials and methods
Collection and housing
Desert gobies were collected from waterholes and springs west of Lake Eyre in South Australia and transported back to the laboratory where they were housed in separate-sex aquaria, kept at a temperature of 24–26°C on a 12 hour light–dark cycle. The aquaria had a fine gravel substrate, rocks and plastic plants for cover, and the water was maintained at a salinity of 5‰. All fish were fed daily on a diet of commercially prepared pellets and frozen brine shrimp (Artemia spp.).
Experimental setup
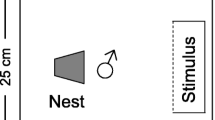
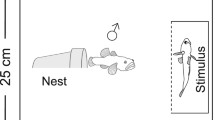
Sexually mature males, identified by their nuptial colouration, were placed in individual aquaria 1 week prior to the experiment. Each aquarium measured (length × width) 30 × 20 cm and was filled with water to a depth of 15 cm. Males were provided with a nest in the form of a 9-cm long PVC pipe (3-cm diameter) positioned horizontally in the middle of the aquarium on top of a layer of fine gravel. Each pipe was capped at one end with the opening facing the front of the tank and was anchored in place by securing it onto a piece of ceramic tile that was buried into the substrate. Males remained in these tanks for the duration of the experiment. At the start of the experiment, a large stimulus female was introduced into a clear container which was placed inside the front of the male’s tank. A sheet of black plastic prevented visual contact between the male and the female. After a 5-min acclimation period, the sheet was removed and we quantified male courtship behaviours directed towards the stimulus female. This was achieved by conducting spot samples every 10 s over a 10-min period. A male was recorded as 'associating' if facing the female while being within 5 cm of her compartment and 'courting' if also performing a fin display (flaring its dorsal and anal fins; Wong and Svensson 2009). After the first presentation, the container with the large female was removed. Males were then haphazardly assigned to one of two treatments (n = 10 per treatment). In the short interval treatment, a small female was introduced shortly after the large female (interval between presentations, 11 ± 0.8 min [mean ± SE]). In the long interval treatment, a small female was introduced the next day (interval between presentations, 24 ± 0.1 h). For both treatments, the same acclimation and sampling procedures were used as during the presentation of the large female. At the completion of the experiment, the total length of all fish was measured using image analysis of digital photographs. Large females were 55 ± 0.6 mm, whereas small females were 42 ± 0.7 mm. The length of females did not differ between treatments (t-tests comparing large females, small females and the size difference between the large and small female in each replicate, all t 18 < 0.13, P > 0.89). Males were 66 ± 1.1 mm, and their length did not differ between treatments (t-test, t 18 = 0.5, P = 0.65).
In addition to the aforementioned experiment, we reanalysed data from a previous study (Wong and Svensson 2009) to test whether the short interval, in itself, might potentially cause a change in male courtship intensity between presentations (for example, due to male exhaustion, handling stress or habituation). The sequential experiment in Wong and Svensson (2009) used an identical experimental design as the short interval treatment described here, except that half of the males were offered females in the reversed order (first small, then large). Re-analysing this data set therefore allowed us to test for changes in male courtship between the first and second presentation whilst controlling for the effects of female size. Our prediction was that males, on average, should court the second female with equal intensity as the first.
Statistical analyses
Data analysis was performed using R 2.8.0 (R Development Core Team 2008). Parameter estimates are presented as mean ±1 SE. Association and courtship were analysed using mixed models to account for the repeated measurements of male behaviours. The identity of each male was entered as a random factor, while treatment (long or short interval) and female size (large or small) were entered as fixed factors. Values were log + 1 transformed prior to analysis, whenever this improved the normality of residual errors. Models were fitted using maximum likelihood, and likelihood ratio (G 2) tests were used to calculate P-values of interaction terms (Quinn and Keough 2002).
Results
There was a significant interaction between female size and interval between female presentations, both for associations (G 2 = 5.1, P = 0.02) and for male courtship displays (G 2 = 5.8, P = 0.02; Fig. 1). Analysing the two treatments separately, there were significant reductions in courtship in the short interval treatment (paired t-tests, associations: t 9 = 4.2, P = 0.002, courtship: t 9 = 4.1, P = 0.002), but there were no changes in the long interval treatment (paired t-tests, associations: t 9 = 0.04, P = 0.97, courtship: t 9 = 0.16, P = 0.89; Fig. 1).

Mean ± SE time males spent a associating with and b displaying to females presented in sequence with either a short or long time interval. Shaded bars represent large females (first presentation); open bars represent small females (second presentation)
When re-analysing data from Wong and Svensson (2009), there was no reduction in overall male courtship intensity between the first and the second presentation (paired t-tests, associations: t 15 = 0.11, P = 0.91; fin displays: t 15 = 0.13, P = 0.90). Thus, removing the effect of female size, males courted at the same level during the first and the second presentation, despite a short time interval.
Discussion
Males dramatically reduced both association time and courtship intensity when they were presented with a small female shortly after encountering a large female. However, female size had no effect on these behaviours if the interval between presentations was longer. Theory predicts that mate encounter rate is crucial in determining whether male mating behaviours will be affected by female reproductive quality (Bonduriansky 2001; Kokko and Johnstone 2002). Our empirical results suggest, for the first time, that males adjust their courtship in regard to female fecundity only if the time interval between female encounters is short. Until now, mate encounter rate has only been shown to affect mating decisions in females, for example, in sticklebacks, where female interest in dull males increased as the time interval between male encounters grew longer (Bakker and Milinski 1991; Milinski and Bakker 1992).
In contrast to females, males are generally expected to maximise their reproductive success by increasing their number of sexual partners (Kokko and Johnstone 2002). Any delay in mating may therefore translate into lowered male reproductive success. Thus, even though a high-quality female may increase male reproductive payoffs, males should not necessarily forego the opportunity to attract lower-quality females (Rowland 1982; Verrell 1995). However, because the intensity of male courtship displays can be important for female choice, and courtship can be costly, males may direct their efforts strategically by courting particular females more than others (Stuart-Fox and Whiting 2005; Reading and Backwell 2007). Such strategies may be especially important in species where males are constrained by the number of clutches they can accommodate in their nest, a common scenario in nest-guarding gobies (Lindström 1992). Indeed, when given a choice, male desert gobies direct less courtship toward less fecund females (Wong and Svensson 2009). Our current results suggest that this discrimination disappears if we simulate a scenario where females are encountered infrequently. Temporal costs therefore seem important for how males will balance the trade-off between the quality and quantity of mating partners.
Although our results can be interpreted as an adaptive male strategy, an alternative explanation is also possible, namely, that a long time interval between female encounters simply obfuscated accurate size comparisons (MacLaren and Rowland 2006). However, regardless of the proximate reason for the behavioural differences between the two treatments, the actual outcome remains the same: when males encountered females infrequently, differences in female quality were of less importance for male courtship effort.
The re-analysed data from Wong and Svensson (2009) showed that mean male courtship intensity (i.e. when disregarding female size) did not change between the first and the second presentation of females, despite a short time interval. We can therefore rule out male exhaustion and/or habituation as potential causes for the observed reduction in courtship and association time in the short interval treatment.
In conclusion, our results suggest that mate encounter rate can influence male courtship tactics. Temporal aspects of mate encounters should therefore be considered both when designing mate choice experiments and when investigating how mate selection operates in nature.
References
Allen GR, Midgley SH, Allen M (2002) Field guide to the freshwater fishes of Australia. CSIRO, Collingwood
Bakker TCM, Milinski M (1991) Sequential female choice and the previous male effect in sticklebacks. Behav Ecol Sociobiol 29:205–210
Belles-Isles JC, Cloutier D, Fitzgerald GJ (1990) Female cannibalism and male courtship tactics in threespine sticklebacks. Behav Ecol Sociobiol 26:363–368
Bonduriansky R (2001) The evolution of male mate choice in insects: a synthesis of ideas and evidence. Biol Rev 76:305–339
Hoefler CD, Persons MH, Rypstra AL (2008) Evolutionarily costly courtship displays in a wolf spider: a test of viability indicator theory. Behav Ecol 19:974–979
Jones KM, Monaghan P, Nager RG (2001) Male mate choice and female fecundity in zebra finches. Anim Behav 62:1021–1026
Kokko H, Johnstone RA (2002) Why is mutual mate choice not the norm? Operational sex ratios, sex roles and the evolution of sexually dimorphic and monomorphic signalling. Philos Trans R Soc Lond B Biol Sci 357:319–330
Kotiaho JS, Alatalo RV, Mappes J, Nielsen MG, Parri S, Rivero A (1998) Energetic costs of size and sexual signalling in a wolf spider. Proc R Soc Lond B Biol Sci 265:2203–2209
Kvarnemo C (1994) Temperature differentially affects male and female reproductive rates in the sand goby—consequences for operational sex-ratio. Proc R Soc Lond B Biol Sci 256:151–156
Lindström K (1992) Female spawning patterns and male mating success in the sand goby Pomatoschistus minutus. Mar Biol 113:475–480
MacLaren RD, Rowland WJ (2006) Differences in female preference for male body size in Poecilia latipinna using simultaneous versus sequential stimulus presentation designs. Behaviour 143:273–292
Milinski M, Bakker TCM (1992) Costs influence sequential mate choice in sticklebacks, Gasterosteus aculeatus. Proc R Soc Lond B Biol Sci 250:229–233
Pelabon C, Borg AA, Bjelvenmark J, Forsgren E, Barber I, Amundsen T (2003) Do male two-spotted gobies prefer large fecund females? Behav Ecol 14:787–792
Quinn GP, Keough JK (2002) Experimental design and data analysis for biologists. Cambridge University Press, Cambridge
R Development Core Team (2008) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Reading KL, Backwell PRY (2007) Can beggars be choosers? Male mate choice in a fiddler crab. Anim Behav 74:867–872
Rowland WJ (1982) Mate choice by male sticklebacks, Gasterosteus aculeatus. Anim Behav 30:1093–1098
Stuart-Fox DM, Whiting MJ (2005) Male dwarf chameleons assess risk of courting large, aggressive females. Biol Lett 1:231–234
Verrell PA (1995) Males choose larger females as mates in the salamander Desmognathus santeelah. Ethology 99:162–171
Wong BBM, Svensson PA (2009) Strategic male signalling effort in a desert-dwelling fish. Behav Ecol Sociobiol 63:543–549
Wong BBM, Jennions MD, Keogh JS (2004) Sequential male mate choice in a fish, the Pacific blue-eye Pseudomugil signifer. Behav Ecol Sociobiol 56:253–256
Woods WA, Hendrickson H, Mason J, Lewis SM (2007) Energy and predation costs of firefly courtship signals. Am Nat 170:702–708
Acknowledgements
We thank R. Thompson for field assistance, R. San-Martin for logistical support and T. C. M. Bakker and two anonymous reviewers for insightful comments. This study was funded by grants from the Academy of Finland (TKL), the Finnish Cultural Foundation and the Australian Research Council and complies with all the relevant State and Commonwealth laws of Australia.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by T. Bakker
Rights and permissions
About this article
Cite this article
Svensson, P.A., Lehtonen, T.K. & Wong, B.B.M. The interval between sexual encounters affects male courtship tactics in a desert-dwelling fish. Behav Ecol Sociobiol 64, 1967–1970 (2010). https://doi.org/10.1007/s00265-010-1007-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-010-1007-z