
Abstract
Dispersal and migratory behaviours are often important determinants of gene flow in wild species, and we have studied their role using ringing-recapture data in the Portuguese population of Miniopterus schreibersii, a cave-dwelling bat that forms large maternity colonies. Juvenile dispersal, usually a major agent of gene flow, appears to be negligible, as young females never settled to give birth in foreign colonies. Likewise, there was virtually no dispersal of adult females to foreign maternity colonies. This strong philopatry virtually eliminated female-mediated gene flow, but we found a great potential for male-mediated gene flow among colonies, as regional migrations temporarily joined both sexes from different colonies in the same roosts, during the mating season. In fact, females from some colonies were more likely to mate with males from foreign colonies than from their own, thus potentially bringing home genes of foreign males. In spite of this abundant gene flow, we found a pattern of isolation by distance and even strong barriers to gene flow, which is explained by the fact that migrations were usually too short to allow direct flow among distant colonies. We concluded that potential gene flow is influenced by the distance between colonies and the availability of mating roosts between them. In addition, we found this flow to be asymmetrical, with a dominant direction from the largest to the smallest colonies. Our ringing-recapture estimates of potential gene flow based on dispersal and migratory behaviour are compatible with the genetic structure of the population for both nuclear and mitochondrial DNA. Our conclusions have implications for the conservation of bats with a spatial behaviour similar to that of M. schreibersii. Colonies should be managed individually because strict female philopatry not only promotes their isolation but also minimises the contribution of immigration in rescuing declining colonies. Furthermore, the results underline the importance of preserving mating roosts to maintain gene flow among colonies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Gene flow, the transfer of alleles between populations, is a key factor in the structuring of the populations of a species (McCracken 1987). High rates of gene flow among populations tend to combine their gene pools, resulting in geographically homogeneous species (Frankham et al. 2005). In contrast, low rates allow populations to differentiate as a consequence of drift and local adaptations, resulting in geographically structured species. Genetic analyses have demonstrated that even species that are phenotypically homogeneous can have genetically structured populations (Frankham et al. 2005). To understand how this structuring arises and is maintained, it is important to determine the behavioural factors that control gene flow among populations.
Several aspects of behaviour, such as migration and philopatry, have the potential to strongly influence patterns of gene flow. Although all bats are very mobile, they range from highly migratory to sedentary species, and because migratory behaviour is an important enabler of gene flow, one would expect relatively low levels of genetic subdivision in the most migratory species (Fleming and Eby 2003). Supporting this prediction, genetic studies have found little geographic differentiation in long-distance migrants such as Leptonycteris curasoae (Wilkinson and Fleming 1996), Nyctalus noctula (Petit and Mayer 1999) and Pteropus scapulatus (Sinclair et al. 1996), and in regional migrants such as Myotis lucifugus (Fenton and Thomas 1985), Myotis myotis (Castella et al. 2000) and Carollia perspicillata (Fleming 1988). In contrast, high levels of genetic subdivision have been reported in sedentary species, such as Macroderma gigas (Worthington Wilmer et al. 1994), Myotis bechsteinii (Kerth et al. 2000) and Plecotus auritus (Burland et al. 1999). However, there are also bats, such as Miniopterus natalensis, which have a strong population structure (Miller-Butterworth et al. 2003) despite of migrating extensively. Consequently, there must be behavioural factors that are responsible for the preservation of this structure in spite of the massive long-distance transportation of genes undertaken by migrating animals.
In the case of bats, philopatry seems to play a very important role in the preservation of geographic genetic structure (e.g. Worthington Wilmer et al. 1994; Veith et al. 2004). If bats return to their natal colonies, rather than disperse to new or foreign colonies, then this reduces gene flow and favours the maintenance of structure. A number of migratory bat species have been found to have a highly philopatric behaviour, i.e. low levels of dispersal, such as Myotis grisescens (Tuttle 1976) and M. natalensis (Miller-Butterworth et al. 2003). Consequently, both migration and philopatry are likely to play important, but very different, roles in the determination of gene flow and population structuring in many bat species.
Intensive ringing-recapture studies are a good way to gain information about the dispersal and migratory behaviours of cave-dwelling bats, but to our knowledge none has objectively included a quantitative assessment of the potential role of each of these behaviours on gene flow. In this paper, we use the results of a large scale ringing project of a colonial bat Miniopterus schreibersii in Portugal to assess three potential sources of gene flow. Two of them are related to philopatry: dispersal of juveniles to foreign colonies and colony switching by adults. The third, mating between individuals of different colonies, is mostly dependent on the patterns of regional migrations during the mating season. Finally, we relate our results to the genetic structure of the Portuguese populations of this species, which has recently been studied (Ramos Pereira et al. 2009).
Methods
Following the latest taxonomic revisions (Tian et al. 2004; Miller-Butterworth et al. 2005), we consider that the range of M. schreibersii is restricted to the Mediterranean region, and we refer to the southern African form as M. natalensis.
M. schreibersii occurs throughout the Portuguese mainland and roosts almost exclusively underground, especially in caves and abandoned mine galleries. It is a gregarious species, forming large colonies throughout most of the year (Rodrigues 1999). Twelve maternity colonies and 15 important hibernation roosts (i.e. used regularly by more than 100 individuals) are known in Portugal. Since the country has been thoroughly surveyed since 1987, it is unlikely that large colonies, particularly nurseries, remain unknown.
Over 36,000 bats were marked with 4.2-mm smoothed metal lipped rings (made initially by Lambournes and later by Porzana, UK), designed to minimise the risk of damage to the wing. This is the ring size recommended by Resolutions 4.6 and 5.5 of EUROBATS. The impact of ringing on bats in this study was monitored and was apparently minor (Rodrigues and Palmeirim 2008), but the same low incidence of problems cannot be assumed for other bat species or ring types.
The ringing programme had two phases: from 1987 to 1992, about 27,000 bats were ringed throughout the whole country, mainly to map migration routes and obtain information to establish a network of protected sites. From 1993 to 2005, ringing was limited to a few roosts to obtain long-term data on the population dynamics of their colonies. A total of 36,269 bats were ringed in the two phases (16,394 adult females, 13,424 adult males, 2,997 juvenile females and 3,454 juvenile males). We obtained a total of 8,007 recaptures of ringed bats (3,947 adult females, 3,186 adult males, 419 juvenile females and 455 juvenile males). Colleagues working in Spain reported an additional 120 recaptures in that country.
We required a minimum of two recaptures to validate a connection between two roosts, thus minimising the effects of unusual behaviour and of possible errors reading the rings.
Bats were normally captured while emerging from roosts using harp traps (Palmeirim and Rodrigues 1993). When that was not possible, they were captured inside the roosts by hand or hand net. To minimise disturbance, bats were not trapped in maternity roosts until after all the young-of-the-year were already flying. The numbers of bats assigned to each roost are the result of direct daytime counts or visual estimates. When the animals formed compact clusters, their number was estimated using the approximate surface area covered by the clusters and a reference density of 2,000 individuals/m2 (Rodrigues and Palmeirim 2008).
Any maternity colony other than the one where a bat was born is referred to as a ‘foreign colony’. We assumed that the females found in a maternity colony during the maternity season had given birth there, because we have previously determined that they very seldom change roost during this period (Rodrigues and Palmeirim 2008). We considered two age classes for both females and males: adults (at least 1 year old) and young-of-the-year (henceforth referred to as ‘juveniles’). Age was determined by the ossification of the carpal joints, development of nipples and testes (e.g. Dwyer 1963a; Baagøe 1977), and the patterns of moult (Dwyer 1963b). The annual cycle was divided into four periods: hibernation (mid-December to end of February), spring (beginning of March to end of May), maternity (beginning of June to end of July) and autumn (end of July to mid-December).
M. schreibersii is known to make seasonal migratory movements of usually a few dozen kilometres, but up to several hundred kilometres (Rodrigues 1999). These movements bring together, during the mating season, males and females from different maternity colonies. We considered as mating roosts those occupied during this season and where we observed copulating animals. Using the recaptures of ringed bats, we determined which mating roosts were used by the animals from each colony and quantified their relative importance for each colony. We assumed that in mating roosts harbouring bats from different maternity colonies, the probability that two bats mate is random and independent of their origin. While there are no behavioural observations confirming this assumption, there are several genetic studies demonstrating that bats often mate with foreign animals, for example in M. bechsteinii (Kerth et al. 2003), P. auritus (Veith et al. 2004) and N. noctula (Petit and Mayer 2000). In each of those mating roosts, we then estimated the probability of a bat from a particular colony mating with animals from each of the colonies represented in the roost. For example, if 70% of the bats in a mating roost were from colony A, 20% from B and 10% from C, the probability of a bat from colony A mating with an animal from colonies A, B and C was 70%, 20% and 10%, respectively. Pooling this information for all the mating roosts used by animals from colony A, we estimated their mating potential with animals from each of the studied colonies.
Mating in M. schreibersii occurs mainly in autumn (Serra-Cobo 1989), but since these bats normally remain in the mating roost through the winter (Rodrigues and Palmeirim 2008), we pooled the recaptures from both seasons to make more robust estimates of the usage of each mating roost by bats from different colonies.
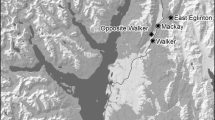
We analysed mating potential using only data from the first phase of the ringing programme to avoid biases resulting from the very uneven sampling during the second phase. We analysed only seven maternity colonies (Miranda do Douro, Tomar, Alcanena, Marvão, Sesimbra I, Moura I and Loulé I; Fig. 1) because data for the remaining five Portuguese colonies were considered insufficient.

Location of studied roosts (black dots represent maternity roosts and white dots mating roosts) and of limestone areas (shaded)
We tested for differences in philopatry between the two sexes by comparing the proportion of males and females that were recaptured in the maternity roost where they were first ringed (Newcombe 1998).
We conducted Mantel tests (due to the non-independence of the data) between the matrix of mating potential among colonies calculated using the results of the ringing study, and: (1) the matrix of geographical distance and (2) the matrix of nuclear DNA differentiation, presented in Ramos Pereira et al. (2009). These authors amplified five microsatellite loci (Miller-Butterworth et al. 2002) and mitochondrial D-loop (Miller-Butterworth et al. 2003) from tissues obtained from 407 bats of 11 Portuguese maternity colonies of M. schreibersii (including the seven colonies analysed in this study). In addition, we conducted a partial Mantel test to check if nuclear genetic differentiation was explained by mating potential even after accounting for geographical distance. As the original matrix of mating potential is not symmetrical, in order to run the multiple Mantel tests, we used the matrix of average mating potential among colonies, which is symmetric. This matrix was obtained by transforming the original matrix of mating potential into the sum of two symmetric matrices: a first matrix which is proportional to the average mating potential among colonies and a second matrix which is proportional to the difference in mating opportunities between colonies (Harvey 1994).
Results
Juvenile dispersal
No cases of juvenile dispersal were registered. In fact, during the maternity season, all the recaptures of bats that had been marked in the roost where they were born (n = 283) took place in that same roost: 51 bats (31 females, 20 males) were recaptured there in their second year and 232 bats (136 females, 96 males) in later years. So all the females returned to give birth in the colony where they were born, demonstrating strict natal philopatry.
The finding that these young bats do not move to other maternity roosts is not a consequence of not knowing their location. In fact, juvenile M. schreibersii often made temporary visits to roosts of foreign nurseries. Of the 75 juveniles (37 females and 38 males) recaptured in roosts of maternity colonies outside the maternity season, 23% were in the roosts of foreign colonies (seven females and 10 males). However, these are just temporary users, as only bats found in foreign colonies during the maternity season can be considered true dispersers.
Adult dispersal
We observed very few cases of adult female dispersal, i.e. females that switched colonies during the course of their reproductive life. During the maternity season, 99.5% of recaptured adult females (n = 1,120) were in the same maternity roost where they had given birth before.
Most males do not spend the maternity season in the nurseries, and they do not have any known role in them. However, the males that are with the maternity colonies also show a high degree of attachment to them. The great majority (93.7%; n = 327) of the recaptures in such colonies were of males that had been ringed in them, but males do disperse to foreign colonies significantly more often than females (χ 2 = 38.7; p = 0.00003).
In spite of this attachment to their colonies during the maternity season, adult bats of both sexes know the location of foreign maternity roosts and regularly visit them. In fact, 28% of the females and males recaptured outside the maternity season (n = 629) were visiting roosts that harbour foreign maternity colonies.
Potential mating with individuals of foreign colonies
Table 1 shows the mating potential between the maternity roosts, estimated using the ringing-recapture data. It is interesting to note that while some colonies show maximum mating potential with their own members (e.g. Loulé I), in other colonies bats seem to have more opportunities to mate with animals from foreign colonies (e.g. females from Tomar seem to have twice as many opportunities to mate with males from Alcanena than with males from their own colony). Our data show that potential mating opportunities between maternity colonies decrease with the distance between them (Fig. 2, Mantel test; r = −0.55, p = 0.001).

Potential mating opportunities between maternity roosts vs. distance between them
In spite of the potential of gene flow through mating with individuals from different colonies, the population is divided into subpopulations, although the level of isolation among them is variable (Table 1). The northern subpopulation (Miranda do Douro) seems to be very isolated from the rest, and the colonies in the west centre (Tomar, Alcanena, Sesimbra I) are isolated from that in the south of the country (Loulé I). The situations of the colonies of Marvão and Moura I are less clear. Marvão has relatively few links to the west-centre subpopulation, and Moura I seems to be even more isolated.
The values of nuclear genetic differentiation (Table 2) are consistent with the estimates of mating potential between colonies, as those with greater mating potential are genetically more similar at the nuclear DNA level (Fig. 3, Mantel test; r = −0.49, p = 0.009). Even when taking into account the geographical distance, nuclear genetic differentiation is still explained by the mating potential (partial Mantel test; r = −0.45, p = 0.048). A correlation between mating potential and mitochondrial DNA differentiation is not to be expected because this DNA does not include any contribution from the males with which the females mate. Mitochondrial DNA differentiation between colonies is higher than that observed for nuclear DNA (Table 2), indicating that isolation by distance is relevant for the studied mitochondrial markers, presumably because of the very strong philopatry of females to their natal colonies (Ramos Pereira et al. 2009).

Potential mating opportunities between maternity roosts vs. nuclear genetic differentiation (F ST) between them
Discussion
Contribution of juvenile dispersal to gene flow appears to be low
Juvenile dispersal is usually the single largest (and often only) long-distance movement made by individual animals of many species (Dice and Howard 1951), and is generally accepted as the major agent of gene flow among populations (Wiklund 1996). It has been observed in several bat species that juveniles of both sexes of Lophostoma silvicolum (Dechmann et al. 2007) and Phyllostomus hastatus (McCracken 1987) dispersed before their first birthdays, and juvenile males of Desmodus rotundus dispersed before reaching sexual maturity (McCracken 1987). However, we did not observe any case of juvenile dispersal to foreign nurseries in this study, and since our sample was quite large we can establish that such events, if they occur, are rare. Low rates of juvenile dispersal have also been reported for some bat species, such as P. auritus (Burland et al. 2006) and M. lucifugus (Humphrey and Cope 1976), and for females of D. rotundus (McCracken 1987) and M. grisescens (Tuttle 1976). In spite of the long-distance movements that juveniles of M. schreibersii make in their first year of life, juvenile dispersal appears to be a minor agent of gene flow among populations, although more ringing data would be required to establish that with certainty.
Adult dispersal contribution to gene flow is also minor
Bats can live for many years and reproduce throughout most of their lives (e.g. Ransome 1995), and we captured ringed M. schreibersii females at least 10 years old that were lactating. Consequently, during the reproductive life of a female bat, there are plenty of opportunities for it to switch the maternity colony where it gives birth, and this switch would result in a female-mediated transfer of genes between colonies, with the addition of a new matriline to the receiving colony. However, in the case of M. schreibersii, we found that such events are extremely rare. Even the observed 0.5% of recaptures of females in foreign colonies during the maternity season may not correspond to real switches between colonies, as some of these females could be in the wrong colony because of unusual circumstances. In one instance, we found several adult females in a foreign maternity colony during the nursing season, but immediately after that we checked their original colony and discovered that its animals had abandoned the roost because almost all the juveniles had died for some unknown reason. Presumably, as their young died, they no longer had any reason to remain in the roost and so migrated prematurely. These females were not quantified as dispersers in our study, but there may have been other similar events that we did not detect and that may explain the presence of a few recaptures in foreign maternity colonies.
Males also showed a high level of philopatry, but they were somewhat more likely to disperse to roosts harbouring foreign nurseries than females. Presumably, such male dispersal events are of little consequence for gene flow because females do not mate with the males in the maternity roosts. Generally, females leave these roosts when lactation is finished.
Our findings confirmed the strictly philopatric behaviour of M. schreibersii during the maternity season, already described by Palmeirim and Rodrigues (1995) using a smaller dataset. Female philopatry to maternity roosts is common in temperate bats, but its strength varies among species; colony switching was not observed in an intensive ringing-recapture study of M. bechsteinii (Kerth et al. 2002) but was not rare in M. myotis (Zahn 1998).
The combination of natal philopatry in juvenile females and reproductive philopatry in adult females makes female-mediated gene transfer virtually non-existent, and this has an impact on the genetic structure of population at the mitochondrial DNA level, as discussed below.
In this study, we only measured the potential dispersal to sites already harbouring maternity colonies. However, some artificial roosts (e.g. mine galleries) have been colonised by M. schreibersii, so some amount of dispersal to new sites must occur. But because new colonies are very seldom formed, we assume that either the number of animals dispersing in this way is very small or their success getting established is extremely low.
Male-mediated gene flow is strong, thanks to seasonal migrations that allow them to mate with females of foreign colonies
Regional migrations thoroughly mix males and females from different maternity colonies during the mating season, and this creates opportunities for gene flow. In fact, our ringing-recapture data suggest that the impact of this mixture on the mating opportunities is such that females from some colonies were far more likely to mate with males from foreign colonies than with males from their own colony. Genetic paternity studies done on other temperate bats, such as M. myotis (Petri et al. 1997), P. auritus (Burland et al. 2001) and M. bechsteinii (Kerth and Morf 2004), found low numbers of juveniles fathered by within-colony males. Consequently, this suggests that regional migrations, rather than dispersal, are the main agent of gene flow between colonies of M. schreibersii.
Despite this facilitation of inter-colony mating by regional migrations, it is clear from the values in Table 1 that the frequency of inter-changes varies substantially among the studied colonies; 79% of the females from Tomar presumably mated with foreign males, but the connections between some other pairs of colonies was nil (which does not mean that they were completely isolated, as they may have mated with colonies in Spain, which we did not study). So what are the factors that are behind these major variations in potential gene flow?
It is clear that distance between maternity colonies plays an important role in the determination of the amount of potential gene flow among them. This influence of distance occurs because most regional migrations tend to be just a few dozen kilometres long, and in the study area many colonies are separated by greater distances (Rodrigues and Palmeirim 2008). Consequently, most migrations are only likely to bring together animals from the closest maternity colonies (such as the various colonies of central Portugal).
But distance alone does not fully explain the observed variations in potential gene flow. For example, the Tomar colony is closer to Marvão than to Sesimbra I, but its females were four times as likely to mate with males from the latter colony. Regional differences in the availability of potential mating roosts explain this discrepancy; the region between Tomar and Sesimbra I includes limestone areas with caves, whereas there are none between Tomar and Marvão. Rossiter et al. (2000) also suggested that inter-colony gene flow in Rhinolophus ferrumequinum is probably limited by both the distances travelled by bats during the mating period and the availability of suitable mating sites.
Another finding that stands out from Table 1 is that potential gene flow is not symmetrical. For example, a female from Tomar had a 45% chance of mating with a male from Alcanena, but the probability of the reverse was only 16%. This is probably due to the fact that Alcanena is a larger colony, so it has more males in the mating sites used by both colonies. It seems probable that in general the dominant direction of gene flow is from the larger to the smaller colonies.
In summary, regional migrations created opportunities for intensive gene flow, but this was limited by the distance between nurseries and by the availability of mating roosts between them. In addition, this potential flow was asymmetrical, with greater flow from the larger to the smaller colonies. This scenario probably applies to many other temperate zone cave-dwelling species, for example M. myotis (Zahn and Dippel 1997) and Myotis nattereri (Rivers et al. 2006), as they often make regional migrations that mix animals from different maternity colonies in mating roosts. However, as these migrations become longer, relative to the distance between roosts, isolation by distance should become less relevant and there should be an increasing tendency for unstructured populations, as those observed in some bat species like L. curasoae (Wilkinson and Fleming 1996) and N. noctula (Petit and Mayer 1999).
Patterns of spatial behaviour explain genetic structure of population
Our observations on philopatry and seasonal migrations of M. schreibersii and the resulting table of potential matting opportunities are congruent with the genetic information available for the same populations (Ramos Pereira et al. 2009). In the remainder of this paper, we discuss how the behavioural patterns here described may have shaped the genetic structure of this population.
Ramos Pereira et al. (2009) found clear inter-colony differences at the mitochondrial DNA level in a study based mostly on the same colonies that we studied (Table 2). As mitochondrial DNA can only be transferred by females, these differences are easily explained by the apparent lack of dispersal by both juvenile and adult females, which eliminated the opportunities for female-mediated gene flow.
However, the structure in nuclear DNA, which is dispersed by both males and females, is much less marked than that of the mitochondrial DNA. This relative homogeneity can be explained by the regional migratory patterns here described, which resulted in a high mating potential between males and females of different colonies. As a consequence, females often bring home the genes of foreign males, thus weakening inter-colony differences at the nuclear DNA level.
The clear correlation between our ringing-recapture-based estimates of mating opportunities among colonies and the nuclear DNA differentiation (even when considering the geographical distance) suggests that spatial behaviour is responsible for those distances. Isolation by distance apparently developed because, as the ringing-recapture results demonstrated, the regional migrations that make inter-colony gene flow possible were usually too short to make direct links between distant colonies. Isolation by distance has rarely been described in bats (Burland and Worthington Wilmer 2001), but has been found in Rhinolophus monoceros (Chen et al. 2006), P. auritus (Burland et al. 1999), Myotis septentrionalis (Arnold 2007) and in autosomal genes of M. bechsteinii (Kerth and Petit 2005).
The structuring in multiple subpopulations that was inferred from the genetic data (Ramos Pereira et al. 2009) was confirmed by our ringing-recapture data, as we found few opportunities for gene flow between those subpopulations. This may be a consequence of the rarity of potential mating roosts between them, which results in gaps of strong resistance to gene flow.
Our results have several implications for the conservation of this species, which is classified as ‘vulnerable’ in Portugal (Cabral et al. 2005). The confirmation with behavioural data of the existence of separate subpopulations underlines the need to treat them as independent management units (Moritz 1999) in order to preserve the full genetic variability of the species and any possible local adaptations. In addition, the confirmation of the strict philopatry to the maternity colonies shows that it is important to monitor and avoid declines in each of them, as one cannot count on a rescue effect of immigrants to help colonies recover. Finally, we have corroborated the importance of preserving mating roosts, already suggested by several authors (e.g. Kerth et al. 2003; Rivers et al. 2006), as they are key in the maintenance of the existing gene flow among colonies.
References
Arnold BD (2007) Population structure and sex-biased dispersal in the forest dwelling vespertilionid bat, Myotis septentrionalis. Am Midl Nat 157:374–384
Baagøe HJ (1977) Age determination in bats (Chiroptera). Vidensk Medd Dan Naturhist Foren 140:53–92
Burland TM, Barratt EM, Beaumont MA, Racey PA (1999) Population genetic structure and gene flow in a gleaning bat, Plecotus auritus. Proc R Soc Lond B 266:975–980
Burland TM, Worthington Wilmer J (2001) Seeing in the dark: molecular approaches to the study of bat populations. Biol Rev 76:389–409
Burland TM, Barratt EM, Nichols RA, Racey PA (2001) Mating patterns, relatedness and the basis of natal philopatry in the brown long-eared bat, Plecotus auritus. Mol Ecol 10:1309–1321
Burland TM, Entwistle AC, Racey PA (2006) Social and population structure in the brown long-eared bat Plecotus auritus. In: Zubaid A, MacCracken GF, Kunz TH (eds) Functional and evolutionary ecology of bats. Oxford University Press, New York, pp 185–198
Cabral MJ (coord.), Almeida J, Almeida PR, Dellinger T, Ferrand de Almeida N, Oliveira E, Palmeirim JM, Queiroz AI, Rogado L & Santos-Reis M (Eds) (2005) Livro vermelho dos vertebrados de Portugal. Instituto da Conservação da Natureza, Lisboa
Castella V, Ruedi M, Excoffier L, Ibanez C, Arlettaz R, Hausser J (2000) Is the Gibraltar Strait a barrier to gene flow for the bat Myotis myotis (Chiroptera: Vespertilionidae)? Mol Ecol 9:1761–1772
Chen S, Rossiter SJ, Faulkes CG, Jones G (2006) Population genetic structure and demographic history of the endemic Formosan lesser horseshoe bat (Rhinolophus monoceros). Mol Ecol 15:1643–1656
Dechmann DKN, Kalko EKV, Kerth G (2007) All-offspring dispersal in a tropical mammal with resource defense polygyny. Behav Ecol Sociobiol 61:1219–1228
Dice LR, Howard WE (1951) Distance of dispersal by prairie deermice from birthplaces to breeding sites. Contrib Lab Vert Biol Univ Mich 50:1–13
Dwyer PD (1963a) The breeding biology of Miniopterus schreibersi blepotis (Temmick) (Chiroptera) in north-eastern New South Wales. Aust J Zool 11:219–240
Dwyer PD (1963b) Seasonal changes in pelage of Miniopterus schreibersi blepotis (Chiroptera) in north-eastern New South Wales. Aust J Zool 11:290–300
Fenton MB, Thomas DW (1985) Migrations and dispersal in bats (Chiroptera). Contrib Mar Sci 27:409–424
Fleming TH (1988) The short-tailed fruit bat: a study in plant–animal interactions. University of Chicago Press, Chicago
Fleming TH, Eby P (2003) Ecology of bat migration. In: Kunz TH, Fenton MB (eds) Bat ecology. University of Chicago Press, Chicago, pp 156–208
Frankham R, Ballou JD, Briscoe DA (2005) Introduction to conservation genetics. Cambridge University Press, Cambridge.
Harvey LE (1994) Spatial patterns of inter-island plant and bird species movements in the Galapagos Islands. J R Soc N Z 24:45–62
Humphrey SR, Cope JB (1976) Population ecology of the little brown bat, Myotis lucifugus, in Indiana and north-central Kentucky. Am Soc Mamm Spec Publ 4:1–88
Kerth G, Morf L (2004) Behavioural and genetic data suggest that Bechstein’s bats predominantly mate outside the breeding habitat. Ethology 110:987–999
Kerth G, Petit E (2005) Colonization and dispersal in a social species, the Bechstein’s bat (Myotis bechsteinii). Mol Ecol 14:3943–3950
Kerth G, Mayer F, König B (2000) Mitochondrial DNA (mtDNA) reveals that female Bechstein’s bats live in closed societies. Mol Ecol 9:793–800
Kerth G, Safi K, König B (2002) Mean colony relatedness is a poor predictor of colony structure and female philopatry in the communally breeding Bechstein's bat (Myotis bechsteinii). Behav Ecol Sociobiol 52:203–210
Kerth G, Kiefer A, Trappmann C, Weishaar M (2003) High gene diversity at swarming sites suggest hot spots for gene flow in the endangered Bechstein's bat. Conservat Genet 4:491–499
McCracken GF (1987) Genetic structure of bat social groups. In: Fenton MB, Racey PA, Rayner JMV (eds) Recent advances in the study of bats. Cambridge University Press, Cambridge, pp 281–298
Miller-Butterworth CM, Jacobs DS, Harley EH (2002) Isolation and characterization of highly polymorphic microsatellite loci in Schreibers' long fingered bat, Miniopterus schereibersii (Chiroptera: Vespertilionidae). Mol Ecol Notes 2:139–141
Miller-Butterworth CM, Jacobs DS, Harley EH (2003) Strong population structure is correlated with morphology and ecology in a migratory bat. Nature 424:187–191
Miller-Butterworth CM, Eick G, Jacobs DS, Schoeman MC, Harley EH (2005) Genetic and phenotypic differences between South African long-fingered bats, with a global miniopterine phylogeny. J Mammal 86:1121–1135
Moritz C (1999) Conservation units and translocations: strategies for conserving evolutionary processes. Hereditas 130:217–228
Newcombe RG (1998) Two-sided confidence intervals for the single proportion: comparison of seven methods. Stat Med 17:857–872
Palmeirim JM, Rodrigues L (1993) The 2-minute harp trap for bats. Bat Res News 34:60–64
Palmeirim JM, Rodrigues L (1995) Dispersal and philopatry in colonial animals: the case of Miniopterus schreibersii. In: Racey PA, Swift SM (eds) Ecology, evolution and behaviour of bats. Oxford University Press, Oxford, pp 219–231
Petit E, Mayer F (1999) Male dispersal in the noctule bat (Nyctalus noctula): where are the limits? Proc R Soc Lond B 266:1717–1722
Petit E, Mayer F (2000) A population genetic analysis of migration: the case of the noctule bat (Nyctalus noctula). Mol Ecol 9:683–690
Petri B, Paabo S, Von Haeseler A, Tautz D (1997) Paternity assessment and population subdivision in a natural population of the larger mouse-eared bat Myotis myotis. Mol Ecol 6:235–242
Ramos Pereira MJ, Salgueiro P, Rodrigues L, Coelho MM, Palmeirim JM (2009) Population structure of a cave-dwelling bat, Miniopterus schreibersii: does it reflect history and social organization? J Hered 100:533–544
Ransome RD (1995) Earlier breeding shortens life in female greater horseshoe bats. Phil Trans R Soc Lond B 350:153–161
Rivers NM, Butlin RK, Altringham JD (2006) Autumn swarming behaviour of Natterer’s bats in the UK: population size, catchment area and dispersal. Biol Conserv 127:215–226
Rodrigues L (1999) Miniopterus schreibersii. In: Mitchell-Jones AJ, Amori G, Bogdanowicz W, Krystufek B, Reijnders PJH, Spintzenberger F, Stubbe M, Thissen JBM, Vohralik V, Zima J (eds) The atlas of European mammals. Academic, London, pp 154–155
Rodrigues L, Palmeirim JM (2008) Migratory behaviour of Miniopterus schreibersii (Chiroptera): when, where, and why do cave bats migrate in a Mediterranean region? J Zool 274:116–125
Rossiter SJ, Jones G, Ransome RD, Barratt EM (2000) Parentage, reproductive success and breeding behaviour in the greater horseshoe bat (Rhinolophus ferrumequinum). Proc R Soc Lond B 267:545–551
Serra-Cobo J (1989) Biological and ecological study of the Miniopterus schreibersii. PhD thesis. Barcelona, University of Barcelona
Sinclair EA, Webb NJ, Marchant AD, Tidemann CR (1996) Genetic variation in the little red flying-fox, Pteropus scapulatus (Chiroptera: Pteropodidae): implications for management. Biol Conserv 76:45–50
Tian L, Liang B, Maeda K, Metzner W, Zhang S (2004) Molecular studies on the classification of Miniopterus schreibersii (Chiroptera: Vespertilionidae) inferred from mitochondrial cytochrome b sequences. Folia Zool 53:303–311
Tuttle MD (1976) Population ecology of the gray bat (Myotis grisescens): philopatry, timing, and patterns of movement, weight loss during migration, and seasonal adaptive strategies. Occas Pap Mus Nat Hist Univ Kans 54:1–38
Veith M, Beer N, Kiefer A, Johannesen J, Seitz A (2004) The role of swarming sites for maintaining gene flow in the brown long-eared bat (Plecotus auritus). Heredity 93:342–349
Wiklund CG (1996) Determinants of dispersal in breeding merlins (Falco columbarius). Ecology 77:1920–1927
Wilkinson GS, Fleming TH (1996) Migration and evolution of lesser long-nosed bats Leptonycteris curasoae, inferred from mitochondrial DNA. Mol Ecol 5:329–339
Worthington Wilmer J, Moritz C, Hall L, Toop J (1994) Extreme population structuring in the threatened ghost bat, Macroderma gigas: evidence from mitochondrial DNA. Proc R Soc Lond B 257:193–198
Zahn A (1998) Individual migration between colonies of greater mouse-eared bats (Myotis myotis) in Upper Bavaria. Z Säugetierkd 63:321–328
Zahn A, Dippel B (1997) Male roosting habits, mating system and mating behaviour of Myotis myotis. J Zool 243:659–674
Acknowledgements
We would like to thank the many colleagues who provided help in the field, especially to Hugo Rebelo, Sofia Lourenço, Tiago Marques, Gabriel Mendes, Francisco Rasteiro, Nuno Vieira and Henrique Vicêncio. Several Spanish colleagues have been kindly reporting the ring numbers of bats that were found in their working areas. Peter Lina helped to find bibliography, and Luka Clarke made useful comments on the manuscript.
Bats were ringed under licence from the ‘Instituto da Conservação da Natureza e da Biodiversidade’.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by G. Jones
Rights and permissions
About this article
Cite this article
Rodrigues, L., Ramos Pereira, M.J., Rainho, A. et al. Behavioural determinants of gene flow in the bat Miniopterus schreibersii . Behav Ecol Sociobiol 64, 835–843 (2010). https://doi.org/10.1007/s00265-009-0900-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-009-0900-9