
Abstract
Large eyespots on the wings of butterflies and moths have been ascribed generally intimidating qualities by creating a frightening image of a bird or mammal much larger than the insect bearing the eyespots. However, evidence for this anti-predator adaptation has been largely anecdotal and only recently were peacock butterflies, Inachis io, shown to effectively thwart attacks from blue tits, Parus caeruleus. Here, we test whether large eyespots on lepidopterans are generally effective in preventing attacks from small passerines and whether the size of insect or bird can influence the outcome of interactions. We staged experiments between the larger eyed hawkmoths, Smerinthus ocellatus, and the smaller peacock butterflies, I. io, and the larger great tits, Parus major, and the smaller blue tits, P. caeruleus. Survival differed substantially between the insect species with 21 of 24 peacock butterflies, but only 6 of 27 eyed hawkmoths, surviving attacks from the birds. Thus, surprisingly, the smaller prey survived to a higher extent, suggesting that factors other than insect size may be important. However, great tits were less easily intimidated by the insects’ eyespots and deimatic behaviour and consumed 16 of 26, but the blue tits only 8 of 25, of the butterflies and hawkmoths. Our results demonstrate that eyespots per se do not guarantee survival and that these two insects bearing equally large eyespots are not equally well protected against predation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The co-evolutionary arms race between predator and prey has led to spectacular morphological and behavioural adaptations in both parties (Dawkins and Krebs 1979). In prey, examples of morphological adaptations range from cryptic colouration (e.g. Cott 1940; Endler 1981; Ruxton et al. 2004), decreasing the risk of being discovered in the first place, to sharp spines and horns, structures useful for defence and retaliation (Edmunds 1974). Behavioural adaptations are equally important in prey and range from choosing a matching background, thereby decreasing the risk of being detected (Endler 1984), to ritualised displays of weapons or armour with the purpose of deterring a predator from attacking (Edmunds 1974; Ruxton et al. 2004).
Edmunds (1974) coined the term deimatic behaviours to include all forms of displays, postures and sounds performed by prey with the apparent purpose of intimidating an attacking predator. A classical example of a structure typically associated with startle displays is eyespots, conspicuous circular colour markings on various animals that resemble the eyes of a vertebrate (Poulton 1890; Cott 1940). Eyespots are most frequently found in butterflies, moths and other insects such as mantids and grasshoppers (Orthoptera) but are also found in other animal groups, for example fish and some frogs (Blest 1957; Edmunds 1974).
The most commonly suggested function of these eyespots is that prey may trick a predator into believing that one of the predator’s own enemies has suddenly entered the scene. Although anecdotal stories of the intimidating qualities of eyespots are abundant, little empirical evidence exists (cf. Ruxton et al. 2004; Ruxton 2005; Stevens 2005). For example, the peacock butterfly, Inachis io, has interested researchers for at least a century as a potential species which uses startle signals to increase its chance of survival (Swinton 1876; Blest 1957; Møhl and Miller 1976). However, experimental proof of the intimidating qualities of the peacock butterfly’s eyespots was only recently published (Vallin et al. 2005).
Earlier studies using abstract models of eyespots have shown that qualities like shape, symmetry, number and orientation of spots influence their signalling strength to, for example, chickens and passerines (Blest 1957; Scaife 1976a,b; Jones 1980; Forsman and Merilaita 1999). Under natural conditions and with naturally occurring eyespots, however, few experimental studies exist. Hence, circular patterns may be effective as signals but whether conspicuous eyespots found on insects generally are intimidating to predators or the peacock butterfly is a singular and exceptional phenomenon remains to be investigated.
Another classical “eye-spot species” in the literature is the eyed hawkmoth, Smerinthus ocellatus. However, its reputation of successfully scaring an attacking predator seems to rest on little more than anecdotal evidence (cf. Cott 1940; Tinbergen 1958; Edmunds 1974; Rothschild 1985) and the only record that we have found comes from a short popular science account by Tinbergen (1958).
Although both peacock butterflies and eyed hawkmoths have similar eyespots that are kept out of sight when at rest, the eyed hawkmoth is a bigger insect than the peacock and their deimatic behaviours differ. When attacked, the peacock flicks its wings open, suddenly exposing its eyespots, and continues to close and flick open its wings and also seems to track and follow the bird predator as long as it is being under attack (Blest 1957). However, the eyed hawkmoth exposes its eyespots on the hindwings by simply protracting its forewings and thereafter performs a kind of rocking movement to and fro by bending its legs while keeping the eyespots fully exposed (Blest 1957; Tinbergen 1958; personal observations).
In this study our main objective was to test whether large “intimidating eyespots” on the wings of butterflies and hawkmoths are generally effective in thwarting attacks from small birds. Our second objective was to test whether the size ratio between predator and prey influenced the outcome of the interactions. To address these issues, we staged trials where living larger, eyed hawkmoths and smaller peacock butterflies were subjected to attacks from two insectivorous passerines, the smaller blue tit, Parus caeruleus, and the larger great tit, Parus major.
Materials and methods
Larvae from wild-caught females of peacock butterflies and eyed hawkmoths were raised on their host plants, stinging nettles, Urtica dioica, and sallow, Salix caprea, respectively. After eclosion, the peacock butterflies were released in a butterfly cage in which they were free to feed on 25% sucrose-solution for 2 weeks to fill their energy deposits. Each butterfly was then transferred to a small plastic cup where they sat on a piece of gauze netting stretched over the cup. In this way, the peacock butterflies were kept in hibernation in a cool (6°C) storage room until the time of the experiment. Eyed hawkmoths do not feed after eclosion and thus were immediately transferred to the storage room where they were kept in plastic cups in the same way as the peacock butterflies.
The experiments were carried out at Tovetorp Zoological research station, situated approximately 90 km southeast of Stockholm in southern Sweden. Blue tits, P. caeruleus, and great tits, P. major, were used as predators in the experiments. Both species are small opportunistic birds that raise their young exclusively on insects, which implies good insect-catching skills. Birds were trapped outside their breeding season close to the research station using mist nets and trap cages (permit Swedish Bird Ringing Centre 619:M03). Thus, because we used wild birds we do not have any knowledge of their previous experience. Housing of birds and experimental setup was reviewed and approved by the regional ethical committee (permit Linköpings djurförsöksetiska nämnd 49-01). Birds were kept individually in indoor cages measuring 80 × 60 × 40 cm, where they had ad libitum access to water, sun flower seeds, suet and mealworms (Tenebrio molitor). Each cage had four perches for the bird to rest on. Any individual bird or insect took part in one trial only. After a finished trial, birds were ringed and released at the site of their capture. No bird was held in captivity for more than a week and all the birds maintained their condition during captivity and were healthy upon release.
Trials were carried out in a room measuring 2.3 × 2.4 × 1.9 m. The room was lit by six day-light fluorescent tubes and had one-way mirrors (80 × 50 cm) on two of the walls, thus allowing the experimenter to observe the animals inside without noticeably disturbing them. A longitudinally cut log of sallow, S. caprea, was placed on the floor so that one of its cut-off ends was in contact with one of the walls. On top of the log, a rough plank (80 × 20 cm) was raised against the wall so that it was more or less in a vertical position. Before the onset of a trial, randomly, either a peacock butterfly or a hawkmoth was transferred, still sitting on its net, to the plank by pinning the net to the plank using map pins. Because of this arrangement, the insect was always positioned close to the floor and thus a bird would not be reflected in either of the one-way mirrors during its approach. To mimic their resting position in nature, the peacocks were placed on the plank with their heads facing down, towards the log and floor and the hawkmoths with their heads up. A small mark on the plank assured that the insects were positioned at the same place and, thus, systematically presented to the birds throughout all trials. A mealworm, T. molitor, was pinned on each side of the hawkmoth or butterfly to encourage the birds to approach the insect. The distance between each mealworm and the butterfly or hawkmoth was 4 cm. The temperature in the room was maintained at 9.9 ± 0.1°C (mean ± SE) to reduce the risk of the insects taking flight during the trials. This is a temperature that both insects will encounter during the cool Nordic mornings and evenings, the two periods during a day when predation by birds is most intense. Trials began with a bird being released into the experimental room through a hatch in the door. We noted the time to the first attack on either of the two mealworms and the elapsed time to the first attack on the butterfly or hawkmoth. A trial lasted for 30 min but was terminated earlier when an insect was seized and subsequently consumed by a bird. We also noted the number of interactions between a bird and an insect that took place during a trial. An interaction was defined to begin with the bird’s attack on the insect and ended when the butterfly or hawkmoth closed its wings again after the bird had retreated as a consequence of the intimidating display by the insect. Thus, attacks that ended with the insect being seized were not counted as interactions. Birds were always allowed to finish eating a seized butterfly or hawkmoth to get a crude idea of the insect’s palatability. All trials were recorded using a digital video camera (Sony DCR-VX1000E). The recordings made it possible to analyse the interactions on a more detailed level. When using direct observations it was, for example, not always possible to decide if an intentional movement from the bird towards the insect made physical contact or not.
For size comparison of species, the length of the left forewing of seven dead peacock butterflies and eight eyed hawkmoths was measured, using a plastic ruler with a millimetre scale. Additionally, we weighed five male and 15 female peacock butterflies that were caught in the wild between 20 April and 7 May at Riala located 50 km northeast of Stockholm, and we also weighed 10 male and 10 female pupae of the eyed hawkmoth obtained from Worldwide Butterflies. Furthermore, we measured the diameter of the hind-wing eyespots of these individuals to reveal potential interspecific differences.
Statistical analysis
All statistical tests are two-tailed and were conducted with Statistica for Windows 5.5 (StatSoft), except for the generalised linear models, which were executed in R 2.1.1 for Windows (R Foundation for Statistical Computing). All values given are mean ± SE. To compensate for multiple comparisons in the frequency tests of number of interactions, sequential Bonferroni corrections (Dunn–Šidák method) were used to achieve the associated critical p values (Sokal and Rohlf 1995).
Results
The forewing of the eyed hawkmoth (mean±SE, 32.4 ± 1.2 mm, N = 8) is longer compared with the forewing of the peacock (28.3 ± 1.6 mm, N = 7) (Mann–Whitney U test, U = 1.5, p = 0.002). The mean weight of eyed hawkmoth pupae was 3.15 ± 0.1 g, whereas that of adult peacock butterflies was 0.15 ± 0.03 g, which means that eyed hawkmoths weighed some ten times more than peacock butterflies, allowing for the fact that about half of a butterfly or hawkmoth’s weight is lost at emergence from the pupa (Wiklund and Karlsson 1984; Fischer et al. 2004). The size of the hindwing eyespots on the eyed hawkmoths (6.8 ± 0.7 mm, N = 8) did not differ from the peacocks’ (6.4 ± 0.5 mm, N = 7) (Mann–Whitney U test, U = 21, p = 0.42). The mean weight of blue tits was 11.73 ± 0.09 g, N = 25, and that of great tits 19.08 ± 0.19 g, N = 26.
All birds (N = 51) eventually attacked the insect and the insects invariably responded to the attack by displaying their eyespots. A total of 13 birds ignored the mealworms and instead attacked the butterfly or the hawkmoth first. There was no difference between treatments (great tit vs eyed hawkmoth, great tit vs peacock, blue tit vs eyed hawkmoth, blue tit vs peacock) regarding the time to when the first mealworm was collected (one-way ANOVA, F 3,34 = 0.84, p = 0.48) or time to the first attack on the butterfly or hawkmoth (one-way ANOVA, F 3,47 = 1.76, p = 0.17).
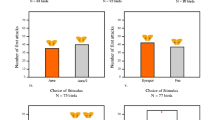
During trials, blue tits had more interactions with peacocks (6.8 ± 1.2) compared with hawkmoths (1.2 ± 0.3) (Mann–Whitney U test, N blue tit vs peacock = 12, N blue tit vs hawkmoth = 13, U = 9.0, p < 0.001; Fig. 1). The same pattern was found for great tits with peacocks (4.6 ± 1.0) and hawkmoths (0.6 ± 0.2) (Mann–Whitney U test, N great tit vs peacock = 12, N great tit vs hawkmoth = 14, U = 10.0, p < 0.001). When comparing the number of interactions between peacocks and blue tits (6.8 ± 1.2) and great tits (4.6 ± 1.0), no difference was found (Mann–Whitney U test, N blue tit vs peacock = 12, N great tit vs peacock = 12, U = 48.0, p = 0.17). Finally, no difference was found between the number of interactions between eyed hawkmoths and blue tits (1.2 ± 0.3) and great tits (0.6 ± 0.2) (Mann–Whitney U test, N blue tit vs hawkmoth = 13, N great tit vs hawkmoth = 14, U = 67.5, p = 0.25).

The number of interactions between the birds and the insects in the different treatments. An interaction was defined to begin with the bird’s attack on the insect and to end when the butterfly or hawkmoth closed its wings again after the bird had retreated as a consequence of the intimidating display by the insect. Thus, attacks that ended with the insect being seized were not counted as interactions. Data are mean ± one SE (whiskers). Numbers above whiskers are N values
A generalised linear model [with logit link function, survival as the binomially distributed response variable, predator (great tit and blue tit) and prey (eyed hawkmoth and peacock butterfly) as crossed factors and type II sums of squares] revealed that more peacock butterflies survived compared with eyed hawkmoths (\( \chi ^{2}_{1} = 27.3 \), p < 0.001; see Table 1 for frequencies), and furthermore there was an effect of predator (\( \chi ^{2}_{1} = 7.9 \), p < 0.005). No interaction between predator and prey was detected (\( \chi ^{2}_{1} = 0.72 \), p = 0.40). All seized peacock butterflies and hawkmoths were, without exception, killed, de-winged and eaten, suggesting that these insects were at least palatable to the birds in this experiment.
Looking at the insects’ first display in the trials, peacocks were more effective, compared with hawkmoths, in making the present bird retreat [generalised linear model with logit link function, retreat as the binomially distributed response variable, predator (great tit and blue tit) and prey (eyed hawkmoth and peacock butterfly) as crossed factors and type II sums of squares; \( \chi ^{2}_{1} = 10.8 \), p < 0.001; see Table 2 for frequencies]. There was no effect of predator (\( \chi ^{2}_{1} = 0.34 \), p = 0.56) and no interaction (\( \chi ^{2}_{1} = 1.14 \), p = 0.29). Thus, although great tits eventually killed and consumed 13 out of the 14 eyed hawkmoths, the birds initially retreated and flew away from the insect in eight trials; hence, seven of these eight great tits returned and killed the hawkmoth within the 30-min time window allotted for the trials. The peacock butterflies were more effective in scaring great tits and only 2 of the 11 birds that retreated from the butterfly after the first attack later returned to kill and eat the insect. In a similar manner, three out of eight blue tits that initially retreated from the eyed hawkmoths later returned and killed and consumed the insect, whereas none of the 12 blue tits that initially retreated from the peacock ever killed a single butterfly, in spite of returning for an average of approximately six times to interact with the peacock (Fig. 1).
Discussion
More peacock butterflies, compared with eyed hawkmoths, survived their encounters with great tits. The same pattern was found when the two insect species were subjected to attacks by blue tits. Thus, both bird species killed more hawkmoths than peacocks. This may seem counterintuitive as both species have eyespots of similar size and, furthermore, the hawkmoth is a bigger prey item. Hence, at first sight there seems to be a potential for the eyed hawkmoth to be a more intimidating prey but, clearly, despite its larger size, the hawkmoths fared worse in repeated interactions with blue tits and great tits. Although larger size did not confer any advantage upon insect survival in our experiment, size did seem to have an effect on bird behaviour; the larger great tits killed a higher proportion of eyed hawkmoths and peacock butterflies than the smaller blue tits.
It may be worth noting that every butterfly and hawkmoth that was seized by a bird was both killed and eaten. Hence, both insects were edible and the difference in survival between the two insects cannot be accounted for by a difference in palatability.
It could be argued that the differences in survival depend, to some extent, on the bird’s perception of the insect before it is attacked and that the peacock butterfly may be more intimidating in its resting posture. This seems not to be the case, however, as there was no difference in either time to when the first mealworm or the insect was attacked. Thus, the cause of the observed differences of survival should instead be found between the initial attack and the end of the trial.
Scaife (1976a) showed that, to chickens, the intimidating effect of eye-like structures is context dependent. An obvious dissimilarity in context concerns the behaviours associated with the exposure of the eyespots in the two species. The peacock butterfly, when resting, keeps all four wings pressed together above its body, perpendicular to the surface. In this posture, only the evenly dark-coloured ventral side of the wings is shown, which, in combination with the irregular shape of the wing margins, makes the butterfly appear like a dead leaf (Brakefield et al. 1992). When disturbed, it rapidly opens its wings, making them parallel to the surface and thereby exposing the strong colours and the conspicuous eyespots on the dorsal surface of the wings. The butterfly then continues to rapidly flick its wings, thus adding movement to the eyespot stimuli (Swinton 1876 in Blest 1957), and also turns around to follow an attacking bird that hops around the butterfly, giving the human observer the impression that it attempts to attack its attacker (personal observation). Eyed hawkmoths, on the other hand, always have their wings parallel to the surface. When a hawkmoth is disturbed, it protracts its cryptic forewings and the bright colours and the conspicuous eyespots on the hind wings are suddenly exposed (Edmunds 1974). The hawkmoths also add movement to the display but, contrary to the peacock, it keeps its wings still once the eyespots have been exposed. Movement of the eyespots is instead created by the hawkmoth alternately bending the forelegs and hindlegs, which creates a kind of rocking movement, although the hawkmoth remains in the same place all the time retaining its grip on the resting surface with its tarsal claws. Comparing the deimatic behaviour of the two insects, that of the peacock appears much more intimidating to the human observer, as well as, judging from our results, to small passerine birds.
Another circumstance that may give the peacock an advantage is that the size difference between a cryptic and a displaying peacock butterfly is relatively larger compared with the hawkmoth. Thus, it is possible that the bird is more intimidated by the initial display of the peacock butterfly and that this effect will influence the butterfly’s chances of surviving the rest of the trial. Looking at the number of interactions, however, we found that both blue tits and great tits had more interactions with peacocks during trials compared with eyed hawkmoths. Thus, it was not the case that the peacocks were left alone after the first interaction; on the contrary, they were attacked repeatedly, the record being one blue tit attacking a peacock 14 times during the 30 min constituting a trial. The observed difference in number of interactions between the two insect species is, in part, a consequence of fewer hawkmoths surviving the whole trial but, nevertheless, indicates that the peacocks’ defence was more effective by repeatedly scaring off attacking birds.
More hawkmoths and peacocks were killed by great tits than by blue tits. Tinbergen (1958) earlier suggested that an eyed hawkmoth might be too big a food item for a small passerine, a suggestion that, at face value, was shown not to hold true in our experiments. However, we did notice a size effect in the sense that the bigger bird was a more ferocious predator, a result which is in line with those of Exnerová et al. (2003), who demonstrated that an aposematic insect was less well protected against larger, compared to smaller, insectivorous birds. Our finding that the smaller prey, the peacock butterfly, was better protected against predation demonstrates that size is not everything and suggests that what counts is the combined effect of size and intimidating quality of the deimatic display.
Few experimental studies still exist on the efficiency of large conspicuous eyespots in insects to protect their bearer against predation. Vallin et al. (2005) showed that the combination of eyespots and the associated display behaviour of the peacock butterfly indeed constitute an effective defence against predation by small insectivorous birds, whereas its close relative, the small tortoiseshell butterfly Aglais urticae, which exhibits the wing-flicking deimatic behaviour but lacks eyespots, is much less efficient in thwarting attacks from blue tits and is usually killed and eaten when attacked (Vallin et al. 2006). The results in this present study are in agreement with Vallin et al. (2005) by again showing that the peacock’s display is efficient in thwarting the attacks of blue tits but also provides evidence that the display is quite effective against a somewhat larger passerine predator, the great tit.
The eyed hawkmoth has for a long time been suggested to be able to effectively defend itself against bird predation (cf. Cott 1940; Tinbergen 1958; Edmunds 1974; Rothschild 1985). However, apart from Tinbergen’s (1958) short popular account, empirical data supporting this idea appear to be lacking and the idea of the eyed hawkmoth’s ability to intimidate an attacking predator seems to have been taken more or less for granted. For example, Edmunds (1974), in his book on defence in animals, says: “One can see how the perfect ‘eye’ of the eyed hawkmoth have evolved from a simple startle display with each modification being more effective than the last at intimidating predators” (p. 168); but no data are given. Indeed, the adaptive value of any conspicuous eyespots found in nature seems so intuitive that Cott (1940) in his classic book on adaptive colouration in animals, when discussing “Such terrifying masks, in which the essential feature is a pair of gleaming eye-spots”, writes: “the coloration is intimately associated with an instinctive display reaction, whose display towards, and effect on, enemies indicate sufficiently clearly its intimidating function” (pp. 387–388).
In our experiments, 5 of the 13 eyed hawkmoths survived in the trials with blue tits, but only 1 of 14 in the trials with great tits, which seems to raise doubts about the adaptive value of the eyespots and anti-predator behaviour. However, before deeming the eyespots and deimatic behaviour inefficient, it is important to realise that the conditions under which the experiments were carried out are likely to underestimate their function because the trials were carried out in a relatively small room from which neither bird nor insect could escape, and so encouraged numerous repeated interactions in which the outcome was often eventually fatal for the insect. However, in a natural setting, it is feasible that a bird that retreated from the eyed hawkmoth during the first interaction would not have returned, in which case, the survival of the eyed hawkmoths would have increased from 1 to 8 out of the 14 that were attacked by great tits, and from 5 to 8 out of the 13 that were attacked by blue tits (Table 2). Regardless of the actual proportion of hawkmoths that survived, it is likely that intimidating eyespots and deimatic behaviour are likely to confer at least some fitness advantage in nature by giving a harmless prey a chance to escape after discovery. Furthermore, birds are not the only threat to these insects, and it is conceivable that other predators may be more intimidated by the hawkmoth’s display.
In conclusion, our results clearly show that the eyed hawkmoths, despite being much larger and having equally large eyespots, were less successful in thwarting attacks of blue tits and great tits compared with the peacock butterflies. Our results also suggest that the size of bird seems to have an effect, with great tits being more inclined than blue tits to attack a butterfly or hawkmoth displaying large eyespots. Finally, our results raise doubts about the efficiency of the anti-predator defence of the eyed hawkmoth and demonstrate that eyespots per se do not guarantee survival and that species with equally large eyespots are not equally well protected against predation.
References
Blest AD (1957) The functions of eyespots in Lepidoptera. Behaviour 11:209–256
Brakefield PM, Shreeve TM, Thomas JM (1992) Avoidance, concealment and defence. In: Dennis RLH (ed) The ecology of butterflies in Britain. Oxford University Press, Oxford, pp 93–119
Cott H (1940) Adaptive coloration in animals. Methuen, London
Dawkins R, Krebs JR (1979) Arms races between and within species. Proc R Soc Lond B 205:489–511
Edmunds M (1974) Defence in animals: a survey of animal defences. Longman, Burnt Mill
Endler JA (1981) An overview of the relationships between mimicry and crypsis. Biol J Linn Soc 16:25–31
Endler JA (1984) Progressive background matching in moths, and a quantitative measure of crypsis. Biol J Linn Soc 22:187–231
Exnerová A, Landová E, Štys P, Fuchs R, Prokopová M, Cehláriková P (2003) Reactions of passerine birds to aposematic and non-aposematic firebugs (Pyrrhocoris apterus; Heteroptera). Biol J Linn Soc 78:517–525
Fischer K, Zeilstra I, Hetz SK, Fiedler K (2004) Physiological costs of growing fast: does accelerated growth reduce pay-off in adult fitness? Evol Ecol 18:343–353
Forsman A, Merilaita S (1999) Fearful symmetry: pattern size and asymmetry affects aposematic signal efficacy. Evol Ecol 13:131–140
Jones RB (1980) Reactions of male domestic chicks to two-dimensional eye-like shapes. Anim Behav 28:212–218
Møhl B, Miller LA (1976) Ultrasonic clicks produced by the peacock butterfly: a possible bat-repellent mechanism. J Exp Biol 64:639–644
Poulton EB (1890) The colours of animals. Kegan Paul, London
Rothschild M (1985) British aposematic Lepidoptera. In: Head J, Emmets AM (eds) The moths and butterflies of Great Britain and Ireland. II. Harley, Essex, pp 9–62
Ruxton GD (2005) Intimidating butterflies. Trends Ecol Evol 20:276–278
Ruxton GD, Sherratt TN, Speed MP (2004) Avoiding attack: the evolutionary ecology of crypsis, warning signals and mimicry. Oxford University Press, Oxford
Scaife M (1976a) The response to eye-like shapes by birds. I. The effect and context: a predator and a strange bird. Anim Behav 24:195–199
Scaife M (1976b) The response to eye-like shapes by birds. II. The importance of staring, pairedness and shape. Anim Behav 24:200–206
Sokal RR, Rohlf FJ (1995) Biometry: the principles and practice of statistics in biological research, 3rd edn. Freeman, New York
Stevens M (2005) The role of eyespots as anti-predator mechanisms, principally demonstrated in the Lepidoptera. Biol Rev 80:1–16
Swinton AH (1876) On stridulation in the genus Vanessa. Entomol Mon Mag 13:169–172
Tinbergen N (1958) Curious naturalists. Country Life, London
Vallin A, Jakobsson S, Lind J, Wiklund C (2005) Prey survival by predator intimidation: an experimental study of peacock butterfly defence against blue tits. Proc R Soc Lond B 272:1203–1207
Vallin A, Jakobsson S, Lind J, Wiklund C (2006) Variation in anti-predator behaviour in three closely related butterfly species—experiments with blue tits as predators on peacocks, small tortoiseshells and commas. Behav Ecol Sociobiol 59:455–459
Wiklund C, Karlsson B (1984) Egg size variation in satyrid butterflies: adaptive vs. historical, “Bauplan”, and mechanistic explanations. Oikos 43:391–400
Acknowledgements and animal welfare
We thank Olof Leimar for statistical advice. This study was financially supported by the Swedish Research Council (to C.W.). Birds were captured with permission from the Swedish Bird Ringing Centre (permit 619:M03). Housing of animals and experimental setup was reviewed and approved by the regional ethical committee of animal experiments (permit Linköpings djurförsöksetiska nämnd 49-01). The experiments described herein comply with the current laws of Sweden.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by D. Gwynne
Rights and permissions
About this article
Cite this article
Vallin, A., Jakobsson, S. & Wiklund, C. “An eye for an eye?”—on the generality of the intimidating quality of eyespots in a butterfly and a hawkmoth. Behav Ecol Sociobiol 61, 1419–1424 (2007). https://doi.org/10.1007/s00265-007-0374-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-007-0374-6