
Abstract
In general, reproductive output in long-lived bird species increases in older compared to younger individuals. Therefore, experienced mates should be advantageous for first-time breeders. To examine requirements and consequences of experienced pair mates we investigated the first pair bonds of common terns, Sterna hirundo, recruiting to their natal colony. We found that male recruits were usually the same age as their mates, whereas female recruits were usually the younger member of the pair. In order to acquire experienced mates, it was necessary for males to arrive early in the year of recruitment. Mates with 2 or more years of breeding experience were only attainable by male recruits characterised by greater body mass and age. Female recruits arrived more than 1 week later than their experienced mates and significantly later in the season than male recruits paired with experienced females. In general, females first bred at a younger age than males, and neither the female recruits’ body mass nor their age was related to the level of experience of their first mate. These sex-specific differences in obtaining an experienced mate did not result in different levels of reproductive success between the sexes. Male and female recruits with mates with 2 or more years’ breeding experience benefited from having experienced mates: they had greater reproductive success. First-time breeders paired with mates with only 1 year of breeding experience did not differ from pairs where both members were breeding for the first time in terms of reproductive success, but clutches were larger. Our results illustrate not only different prerequisites for males and females, but also males’ need for experienced mates. Delayed male breeding (postponing breeding for another year), supposed to be a negative trait, and high body mass, supposed to be a trait of superior individual quality, can be combined in some individuals, improving reproductive success and showing that breeding common terns use a range of tactics to begin reproduction.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In many bird species, reproductive output increases with age, especially in the early and middle stages of life (Saether 1990; Martin 1995; for seabirds, Ryder 1981; Wooller et al. 1992). However, because of the intrinsic relationship between them, it is often impossible to differentiate between the influence of accumulated breeding experience, and of increased reproductive effort with age (Pärt 1995). Many authors have used the terms “age” and “experience” synonymously, because they have not known at what age individuals included in their analyses began reproducing (review in Fowler 1995). Also, it is known that in long-lived animals, an extended period of adjustment is needed before pairs ‘function’ efficiently. Thus, breeding success can depend more on the longevity of the pair bond than on either the members’ age or experience alone (e.g. Bradley et al. 1995; Black et al. 1996; Rees et al. 1996; Bried et al. 2003). For example, in the Cassin’s auklet, Ptychoramphus aleuticus, Pyle et al. (2001) showed that experience with the same mate improved reproductive success (cf. Coulson and Thomas 1983; Ollason and Dunnet 1988; Bradley et al. 1990).
However, only experienced birds have the option of re-pairing with their mates of the previous season. So it is appropriate to consider mate choice in two ways (as suggested by Coulson and Thomas 1983 for the kittiwake, Rissa tridactyla): mate choice in first-time breeders, and in individuals which have bred before and therefore have the possibility of retaining their mate from the previous year. Furthermore, although most studies of pair bonding in seabirds have shown high proportions of pairs in which members are of similar age (Reid 1988), it is well known that great age differences can occur, especially in pairs including young birds (Coulson 1980; Bradley et al. 1995; Nisbet et al. 2002). However, not much attention has been paid to these different-age pairings, probably due to a lack of adequate data. Thus, little is known about why and how such pairs form.
In this paper, we study the requirements and consequences of primary mate choice in the common tern, Sterna hirundo, an iteroparous, colonial breeding and mainly monogamous seabird (González-Solís et al. 2001; Becker and Ludwigs 2004). We address the question: what traits characterise first-time breeders able to obtain experienced mates under natural conditions in a non-experimental situation?
Data were collected during a long-term study of this species, in which a transponder identification technique has been used to identify and collect information on every breeding bird recruited into a colony since 1994 (Becker and Wendeln 1997; Becker et al. 2001). Because of the information available about attendance of each bird at the colony before (Dittmann and Becker 2003), during and after the first breeding attempt (Ludwigs and Becker 2002), we are able to exclude the possible influences of pair bond longevity, and also to determine the precise amount of breeding experience of each recruit’s mates. Hence, our first objective was to show the ages of birds paired with first-time breeders, and the proportions of experienced birds paired with males and females breeding for the first time.
In the common tern, body mass is a parameter indicating individual quality because it is constant between years in individual terns, and positively correlated with breeding success (Wendeln and Becker 1996). The dates at which individuals arrive at the natal colony are important in determining whether they can recruit into the colony (Ludwigs and Becker 2002), hold their mate and breeding status (Gonzáles-Solís et al. 1999) or produce replacement clutches (Wendeln et al. 2000). Our second objective was to determine the influence of arrival date and body mass on the probability of obtaining a specific mate.
Thirdly, we suggest that experienced breeders may improve reproductive performance of first-time breeders with which they are paired, since age-related reproductive performance is increased by experience and by improving behavioural skills (Forslund and Pärt 1995). To separate the effects of males or females with previous breeding experience, we compare the influences of previous breeding experience in pairs consisting of a first-time breeder paired with an experienced breeder, and pairs consisting of two first-time breeders.
Methods
Colony site and registration of terns
The data were collected during an ongoing long-term study of the common tern in the harbour area of Wilhelmshaven (Lower Saxony, German North Sea coast, 53°27′N, 08°07′E). The colony has been monitored since 1984, while since 1992, subcutaneously implanted transponders (microchips or “smart tags”; ID 100, Trovan) have been used to mark all fledged chicks (for details see Becker and Wendeln 1997; Becker et al. 2001).
The colony site consists of six artificial islands of equal size (10.7x4.6 m), arranged in a line with 0.9 m between the islands. The islands are surrounded by concrete walls on which 44 resting boxes (0.3×0.3×0.3 m) are mounted. The walls are surmounted with wires to prevent terns landing anywhere except on the boxes. Each box has an antenna linked to a system which remotely and automatically records the presence of all marked terns, breeders as well as non-breeders (for details see Becker 1996).
The transponders require no battery, have an unlimited life and are activated by the antennas at a distance of <11 cm at any angle. When interrogated by the antenna system, each transponder transmits a unique 10-digit code. By this means, each individual’s presence at a resting box is checked for by the system every 5–10 s throughout the breeding season. The precise times and dates of each individual’s attendance at the colony is thus logged. During the 2002 breeding season, about 300 pairs bred at the colony and nearly 500 terns—breeders and non-breeders—were registered by the transponder system.
We did not observe any negative effects of marking with transponders (see Gonzáles-Solís et al. 1999). The studies were done under licence from the Niedersächsisches Landesamt in Hanover, and the Bezirksregierung Weser-Ems in Oldenburg.
Recruits and experienced breeders
Common terns return at the age of 2 years to their natal colony, and recruit after 0–3 years prospecting (Dittmann and Becker 2003). The term “recruit” is defined as a bird breeding for the first time at the site (native recruit). After marking the first cohort of chicks in 1992, the first native recruit was detected in 1994. Recruits which had 1 or more prospecting years and were used for the analysis in this paper were present at the colony for several days in the prospecting years before recruitment (all prospecting periods pooled: 51±22 days colony attendance per prospecting season, range 7–108 days, n=355, >95% have been >14 days at the colony; for more details see Ludwigs and Becker 2002; Dittmann and Becker 2003). Thus, it seems unlikely that any of the birds bred elsewhere before joining the Banter See colony. This finding clearly demonstrates the effectiveness of the registration system, and indicates that the data are reliable.
“Experience” is defined as the number of years during which a bird bred. An “experienced breeder” is defined as one that has already bred in a previous year (experience ≥1; a recruit has an experience =0). Since 1993, the breeders in the colony were identified by placing a portable antenna at the nest for 1–2 days during the incubation period. Among all the first clutches checked since the inception of the study, 1,126 pairs were found consisting of at least one transponder-marked bird, of which 247 pairs contained at least one transponder-marked recruit. Hence, we categorised the recruits in terms of experience specific pair mates. If an experienced breeder disappeared and the previous partner changed to a recruit we classified the recruits’ mate as “widowed”. If the previous mate of an experienced partner was registered as breeder or non-breeder in the colony during the season the recruit started to breed we classified the experienced mate of the recruit as “divorced”.
Because of immigration into the colony (from 1992–2002 the colony grew from 90 to 300 breeding pairs), the majority (55.1%) of recruit pair bonds (n=247) consisted of one native recruit and one unmarked bird of unknown origin. Within the recruit/unknown-pairs we found no difference in age of recruits between the sexes (2 to 5-year-old recruits; χ2=2.82, df=3; P=0.42). In the adult population, the proportion of native males and females shifted: more females fledged, but native males joined the colony in a slight majority (P.H. Becker, unpublished data). Thus, more males were mated with an unknown partner (79 vs 57). The proportions of different pair mate types (experienced breeder, recruit or unknown tern) did not differ between male and female recruits (χ2=1.77, df=2, P=0.41). Hence the composition of the unmarked (and therefore unknown) mates of recruits seems to be similar to that of the known pairs, and are therefore a representative sample for the analysis in this paper.
We sexed adult birds by observing copulations of marked terns, and since 1998, all fledglings by using routine PCR methods (Becker and Wink 2002).
Reproductive parameters
To record clutch initiation, clutch and chick fate until fledging, we visited the colony every 2–3 days throughout the breeding season until the last chicks fledged. The mean fledging age of common terns was 26 days. Because of interannual variation in reproductive parameters and output (e.g. 1994–2002 means of clutch size 2.2–2.9; reproductive success 0.2–2.4 fledglings/pair; for further details see Becker 1998; Becker and Wink 2003) we calculated reproductive parameters for each recruit relative to those of the colony mean. In the common tern reproductive success is valid for both members of the pair, because extrapair copulations and extrapair paternity is very rare in this species (González-Solís et al. 2001).
Arrival and laying date, arrival mass
For each breeder in each season, the date of first registration at the colony is defined as arrival date. The date on which the first egg was laid is defined as the laying date. This term is used for female as well as male recruits. Because arrival and laying dates differed between years (for details see Ludwigs and Becker 2002), we calculated individual relative dates related to the annual colony mean. The asynchrony in arrival at the colony between two members of a pair is the number of days between their respective arrivals in a given breeding season. The time before laying was measured as the difference in days between an individual’s arrival and the laying dates in the same breeding season. A few birds were not recorded until they were found incubating their clutch. Therefore, we calculated these parameters only if a tern’s arrival date was registered at least 10 days before its laying date (cf. Gonzáles-Solís et al. 1999). This is the main period of courtship feeding in the common tern (Wendeln and Becker 1996), hence the period of establishment and possible maintenance of pair bonds.
Up to 16 resting platforms were equipped with electronic balances (SARTORIUS PT and BL series, accuracy ±1 g) for automatic recording of the body mass of the terns using them (for details see Wendeln and Becker 1996, Dittmann and Becker 2003). For birds with three or more independent mass records per day, mean daily body mass values were calculated (for details see Wendeln and Becker 1996), and the mean of daily body mass values of the 3-day period immediately after a tern’s arrival was used as individual arrival mass in this paper.
Statistics
Student t-tests and Mann-Whitney U-tests are used for independent samples, Wilcoxon test for related samples. Frequencies are tested with χ2-tests. The amount of previous breeding experience is treated as factor in analyses of variance to compare the possible influence on reproductive performance, including sequential Bonferroni adjustment (Holm 1979; Rice 1989). To analyse temporal aspects, the same procedure was used. Pairwise comparisons were performed by Scheffé post-hoc in order to test for significant differences between groups of pairs with members with different degrees of breeding experience. All statistics were calculated using SPSS 10.0. Results are presented as arithmetic means ± standard deviation, and all tests are two-tailed with the significance level set at alpha=0.05.
Results
Pair bond types in recruiting common terns
The majority of experienced breeders which switched mates were divorced and not widowed (Table 1). Of 111 pairs where both partners were known, 59.5% consisted of two recruits and the remaining 40.5% contained a recruit and an experienced bird. Experienced breeders were mostly birds which had bred only once before they paired with a recruit, although some individuals had more than 4 years of breeding experience (Table 2).
Pair bond in relation to age
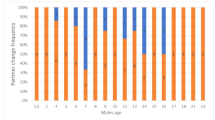
Most male recruits paired with same-aged females (Fig. 1; rS=0.416, P<0.01, n=89), but nearly 50% of female recruits were paired with older males (Fig. 2, rS=0.191, P<0.08, n=87), resulting in a significant difference in relative mate age (older, same aged or younger) between the sexes (data from Fig. 2; χ2=16.73, df=2, P<0.001). However, in pairs where both members were recruits, 58% were the same age.

Relationship between age of male recruits and their female mates in the common tern, Sterna hirundo, including the status of pair mates (black recruit mate; grey experienced mate; the diagonal line connects all same aged birds)

Relationship between age of female recruits and their male mates in the common tern, including the status of pair mates (black recruit mate; grey experienced mate; the diagonal line connects all same aged birds)
The age of male recruits paired with experienced females ranged from 3 to 6 years (Fig. 1), whereas nearly all female recruits pairing with an experienced male were 3 years old (Fig. 2). These patterns resulted in significant age differences between the members of four possible pair bond types (F3,160=9.74, P<0.001). Male recruits pairing with experienced females were the younger on average by 1.0±1.7 years (n=24); female recruits paired with experienced males were on average 2.2±2.6 years (n=21) the younger; the members of recruit/recruit-pairs and pairs of two experienced terns were of similar age with, on average, only slightly older males (0.3±0.9 years, n=64, and 0.3±2.7 years, respectively; the latter value, n=52, is calculated from the age of known pairs of two experienced breeders in 2002). Post-hoc testing clearly showed an age difference of female recruit/experienced male pairs and all other pair types (P<0.005).
The mean age of female recruits did not differ between birds paired with experienced mates (data from Fig. 2: 3.1±0.4 years) or another recruit (3.0±0.6 years, n=87, Mann-Whitney U-test: Z=−0.91, P=0.363). However, male recruits paired with experienced females tended to be older (data from Fig. 1: 4.0±0.8 years) than those paired with female recruits (3.3±0.6 years, n=89, Mann-Whitney U-test: Z=−4.23, P<0.001). Male recruits paired with the most experienced females were also the oldest individuals (Table 3).
Prerequisites of pairing with experienced mates
Recruits who were paired with experienced breeders arrived significantly earlier than recruits paired with another first-time breeder (relative arrival date: 8±11 days vs 16±10 days in pairs of recruits; t166=4.56, P<0.001), but this difference is mainly due to the early arrival of male recruits (Tables 4, 5).
Arrival mass correlated negatively with arrival date at the colony site in male recruits (rP=–0.202, P<0.03, n=125), but not in females (rP=0.008, P=0.93, n=111). Corresponding to this relationship, male recruits paired with ≥2-year-old experienced females had significantly greater arrival mass than other male recruits (Table 4). There was no such relationship in female recruits (Table 5).
Male recruits arrived at a similar time to their experienced mates (2±8 days later). Female recruits arrived more than 1 week later (9±12 days) than their experienced male mates—a significant difference between male and female recruits (n=42, Mann-Whitney U-test: Z=−2.56, P<0.011). Male recruits arrived 5±11 days later than the colony mean arrival date, and not significantly different compared to their experienced females (3±9 days later than colony mean; n=44, Wilcoxon test: Z=−1.74, P=0.081). Female recruits paired with experienced males, arrived 12±11 days later than mean of the colony, and significantly delayed in comparison to their males (3±10 days later than the colony mean arrival date; n=40, Wilcoxon test: Z=−2.88, P<0.005).
Consequences of mates’ breeding experience
The mean clutch sizes of recruits paired with experienced mates was similar to the mean clutch size for the whole colony. Recruit/recruit-pairs laid significantly fewer eggs. The largest clutches were laid by females with ≥2 years’ experience paired with recruits (Table 6).
We found a significantly higher reproductive success among recruits paired with a mate with more than 1 year’s previous breeding experience (Table 6). There was no relationship between the greater reproductive success of recruits paired with experienced mates and their clutch size. The number of fledglings per egg laid (Table 6) as well as the success rate from three-egg clutches confirm these general findings (relative number of fledglings from three-egg clutches: recruit/recruit-pairs −0.3±0.4; recruit/breeder with 1 year’s experience pairs −0.3±0.8; recruit/breeder with ≥2 years’ experience pairs 0.4±0.6; F2,40=5.468, P<0.009).
The only difference in reproductive performance between the sexes of the recruits’ experienced mates was the relative number of chicks which disappeared, which were significant lower if the experienced tern was the female (–0.1±0.1) compared to recruits paired with experienced males (0.0±0.2, n=38, Mann-Whitney U-test: Z=−2.01, P<0.05).
Of recruits paired with mates with no or only 1 year’s breeding experience, 71.3% hatched chicks (n=94). 91.1% of recruits with more experienced mates hatched chicks, and this difference is significant (n=17: χ2=3.982, df=1, P<0.05). This difference between pairs with differing amounts of experience became stronger with respect to their ability to rear young to fledging: 44.8% of 67 versus 81.3% of 16, χ2=6.882, df=1, P<0.01.
Discussion
With whom to pair for the first time?
The proportion of recruit/recruit pairs in which both members were the same age was nearly 60%, so pairing in recruiting common terns appears to be age-assortative. This is often documented in birds (Reid 1988), as also in Sternidae. In many published studies, however, the sample sizes of pairs of known ages have been small, and given without reference to the status and in some cases the sex of the birds (Harrington 1974; Coulson and Horobin 1976; Mills and Shaw 1980).
Nisbet et al. (1984) showed that 39% of 13 pairs of common tern were of birds of equal ages. Of these birds, all but one were 5 years old. These birds could then have been recruits within the age range when common terns are known to begin breeding (Ludwigs and Becker 2002; Figs. 1, 2). Neubauer (1997) found a lower proportion (27%; n=99) of same-age pairs than in our study in a much larger sample than that of Nisbet et al. (1984), although Neubauer (1997) had caught terns >5 years old in a higher proportion. This author also included data on pair bonds persisting over several years, and about 30% of his “pairs” were multiply represented. Coulson (1980) implied that much of the correspondence in kittiwake age between members of breeding pairs is due to the continuity of pair bonds. By reducing the samples in such a way that each pair bond was included only once in the analysis, Coulson (1980) and Bradley et al. (1995) did not find a correlation between the ages of members of pairs in kittiwakes or short-tailed shearwater, Puffinus tenuirostris, respectively. Thus, the discrepancy between the available published data and our analysis of recruit/recruit pairs may arise from higher proportions of already established breeders in the data used by Nisbet et al. (1984) and Neubauer (1997).
In 28 pairs of arctic terns, Sterna paradisaea, Coulson and Horobin (1976) showed that the older members of a pair were mostly males, but in pairs with a large difference in ages, the older members were females paired with 3- or 4-year-old males, probably recruits. In 13 pairs in which neither member was older than 6 years, the males were on average 0.6±1.1 years older than their mates (calculated with the data from Coulson and Horobin 1976). This is comparable to our study with the common tern, and indicates a similar age pattern within pairs containing one recruit in the closely related arctic tern.
Recruiting common terns arrive later than experienced breeders (Ludwigs and Becker 2002). Therefore the pool of potential mates for arriving recruits consists mainly of birds who also arrived later, and were of similar age. Thus, recruits may only have the choice of mates which are almost all recruits themselves (for gulls, see Coulson and Thomas 1983; Klomp and Furness 1991). Probably for the same reason, we found that most recruits were paired with another first-time breeders. However, among recruiting common tern pairs, slightly more than 40% had a partner which had already bred in a previous season, and only in pairs containing a male recruit were the ages of members of a pair positively correlated. Even if females had some previous experience, some male recruits were the older member of the pair (Fig. 1). Finally, the diverging age of male recruits (mainly of same age as their mates) and female recruits (mainly younger) obviously resulted from the pairs consisting of a recruit and an experienced breeder.
How to get an experienced mate
Ludwigs and Becker (2002) demonstrated that arrival time at the colony was a decisive factor in determining whether or not a prospecting common tern will recruit. The results of this paper show that early arrival also helps to decide with whom a recruit will mate. It is generally supposed that the reason for age-related mating in seabirds (especially between new pair members or first-time breeders) is their concurrent arrival at the colony (for references, see above). However, the precise prerequisites for pairing with an experienced mate in the common tern were different for male and female recruits. Male recruits arrived at almost the same time as their experienced mates, and significantly earlier than female recruits paired with experienced males. Hence, the threshold arrival date for facilitated recruitment postulated by Ludwigs and Becker (2002) seems to occur earlier in males, particularly if they are to pair with experienced partners.
Dittmann and Becker (2003) found a positive relationship between body mass and early arrival in prospecting common terns, and that birds became heavier between one prospecting season and the next one (see also Weimerskirch 1992). Our results indicate that high body mass in male recruiting common terns is related to earlier arrival, or offers the opportunity of access to experienced females, respectively. However, all male recruits which were paired with experienced females, irrespective of the amount of previous breeding experience in the female, showed similar arrival dates. Significant mass differences could only be found in the small group of males paired with females of 2 or more years of breeding experience. This finding leads us to conclude that higher body mass seems to influence the chance of pairing with an experienced female as an effect secondary to arrival time or in a ‘second step’, respectively. Potential recruits (all of whom have arrived early enough to start breeding with an experienced female) could be separated into birds of lesser and greater body mass and competitive abilities, of which the latter can win more experienced mates.
The shorter time between arrival and laying in pairs containing early-arriving male recruits with the greatest body mass on arrival supports this finding. In particular, because pairs containing a male recruit and a female with 2 or more years of breeding experience produced more eggs despite the shorter time available for courtship feeding. Probably, these males with a greater body mass are able to feed their females adequately, and can replace disappeared or delayed males, or even displace experienced males in competition: the majority of experienced females in pairs containing male recruits were divorced, not widowed.
What are the consequences of pairing with an experienced mate?
In recruiting common terns, the clear advantage of breeding with an experienced mate was shown by a significantly higher breeding success for the recruit compared with pairs consisting of two first-time breeders. The general finding that more experienced birds show greater reproductive success has been shown for various seabird species (Davis 1976; Ollason and Dunnet 1978; Wooller et al. 1990; Hamer and Furness 1991; Ratcliffe et al. 1998) and even for songbirds (Harvey et al. 1979; Sasvári and Hegyi 2000). In the common tern, age affects reproductive performance profoundly until the age of 5 years (Nisbet et al. 1984; Gonzáles-Solís et al. 2004), at which age nearly all common terns had begun breeding (Ludwigs and Becker 2002).
Our results allow a more detailed view of the influence of mate experience within individual known pairs and indicate a clear effect of previous breeding experience on reproductive performance. The number of eggs per clutch was significantly greater in pairs in which one member had at least 1 year previous breeding experience than in pairs consisting of two first-time breeders. In terms of reproductive success, however, significant differences only became evident if the experienced partner had more than one previous season’s experience. The number of fledglings produced increased with the degree of experience in male and female mates, indicating the positive influence of experience for reproductive success in both sexes.
The positive effect of greater experience in birds generally is related to improvements in competence in such skills as foraging and chick feeding, resulting in greater reproductive success overall (Rowley 1983; Village 1986; Pyle et al. 1991; Forslund and Larsson 1992; Weimerskirch 1992; Bradley et al. 1995; Newton and Rothery 1998; Pyle et al. 2001; Nielsen and Drachmann 2003). This can also be assumed in common terns during the beginning of their reproductive career. So, our results support the constraint hypothesis (Curio 1983; Forslund and Pärt 1995), at least during the early stages of the lifespan of the common tern.
The only significant difference made by the gender of recruits’ experienced mates was the lower number of disappeared chicks in pairs with experienced females. This may be the result of the more pronounced effort by females in brooding the chicks in common terns (Wiggins and Morris 1987; Wendeln 1997). Intra-specific kleptoparasitism can cause unguarded chicks to be removed from the colony together with stolen fish (Ludwigs 1998, Sudmann 1998). In the colony we studied, chick-removal incidents were observed every year and were probably the main cause of chicks’ disappearance, possibly assisted by insufficient attendance by inexperienced females. Male common terns attend the brood less, because during the first week of chicks’ life they can provide up to 90% of feeds (Wiggins 1989; Burness et al. 1994; Galbraith et al. 1999). Thus, after being fed by their father, chicks may be left unguarded if the female is absent, leaving them vulnerable to attacks from other birds. Our findings suggest that sex-related roles have, at least in part, to be learned by recruiting common terns, to deal with the demands of breeding.
How male and female recruits start reproduction
The majority of common tern recruits paired with other first-time breeders, but some individuals reached the colony earlier in the season than others, and benefited by then being able to pair with experienced mates which thereafter were not available for the later-arriving recruits. In the wandering albatross, Diomedea exulans, Jouventin et al. (1999) found that recruits were paired with mates of similar ages to themselves, but preferred those with the most experience. The best option to enlarge the pool of potential experienced mates for recruiting common terns seems to be to arrive at the colony as early as possible in the season. It is remarkable that those male recruits paired with the most experienced females tended to be the oldest and heaviest. Ludwigs and Becker (2002) showed that recruits prospecting the colony over a number of seasons, and hence with increasing age and sexual maturity, arrive in the colony in the season prior to recruitment at around the mean arrival date of birds which did breed, and thus actually early enough for to breed themselves, but nevertheless did not do so. So, at least some males choose to delay their first breeding attempt in order to obtain an experienced mate, with the concomitant effect of greater breeding success.
Ens et al. (1996) interpret vacancies as opportunities to switch to a better mate, and changes in arrival time at the colony as an indication of birds’ quality. Similar processes could be assumed for recruiting male common terns, but our results indicate that this would not seem to be valid for all males. Conversely, nearly all female first-time breeders paired with an experienced male were 3 years old. Moreover, female recruits arrived later in the season and, if they paired with experienced males, they showed neither earlier arrival dates nor greater masses on arrival than male recruits. They also arrived much later compared to their experienced mates. In comparison, male recruits paired with experienced females arrived on about the same day. It seems probable that female recruits may choose the experienced males from the pool of predominantly inexperienced candidates.
In the long term, behavioural improvements during the first breeding attempt seem to have no strong effect on reproductive success. If both members of a long-standing pair co-ordinate their arrival times and breeding performance over the years, they may improve their reproductive rate by maintaining their output of fledglings even in years when conditions are poor. Females did not have the same requirements in order to start reproduction, and male recruits may adjust their mating opportunities according to their status by (1) arriving at the colony earlier in the year, or (2) delay recruitment until another year in order to pair with experienced females. Despite the fact that only minorities of individuals seem to decide to delay their recruitment, we show some evidence that male common terns may use this as alternative tactic to recruit into the breeding population. This study shows some evidence that experience affects reproductive success also in males, in particular because extra-pair copulations and extra-pair paternity are exceptional in common terns (González-Solís et al. 2001, Ludwigs and Becker 2002). The lifespan of the common tern can be over 20 years (Nisbet et al. 2002). Consequently, over their lifetime, the greater reproductive success of older male recruits paired with the most experienced females may be outweighed by younger recruits, as they, though less experienced and with lower reproductive output in their early breeding attempts, will have more breeding years in their lives, to reach at least a similar lifetime reproductive success.
References
Becker PH (1996) Flußseeschwalben in Wilhelmshaven. Oldenburger Jahrb 96:263–296
Becker PH (1998) Langzeittrends des Bruterfolgs der Flussseeschwalbe Sterna hirundo und seiner Einflußgrößen im Wattenmeer. Vogelwelt 119:223–234
Becker PH, Ludwigs J-D (2004) Sterna hirundo common tern. BWP Update 6 (in press)
Becker PH, Wendeln H (1997) A new application for transponders in population ecology of the common tern. Condor 99:534–538
Becker PH, Wink M (2002) Geschlechtsabhängige Größenunterschiede von Flüglingen der Flussseeschwalbe (Sterna hirundo). J Ornithol 143:51–56
Becker PH, Wink M (2003) Influences of sex, sex composition of brood and hatching order on mass growth in common terns Sterna hirundo. Behav Ecol Sociobiol 54:136–146
Becker PH, Wendeln H, Gonzáles-Solís J (2001) Population dynamics, recruitment, individual quality and reproductive strategies in Common Terns Sterna hirundo marked with transponders. In: Jenni L (ed) Bird ringing 100 years. Ardea 89 (special issue): 241–252
Black JM, Choudhury S, Owen M (1996) Do barnacle geese benefit from lifelong monogamy? In: Black JM (ed) Partnerships in birds – the study of monogamy. Oxford University press, Oxford, pp 91–117
Bradley JS, Wooller RD, Skira IJ, Serventy DL (1990) The influence of mate retention and divorce upon reproductive success in short-tailed shearwaters Puffinus tenuirostris. J Anim Ecol 59:487–496
Bradley JS, Wooller RD, Skira IJ (1995) The relationship of pair-bond formation and duration to reproductive success in short-tailed shearwaters Puffinus tenuirostris. J Anim Ecol 64:31–38
Bried J, Pontier D, Jouventin P (2003) Mate fidelity in monogamous birds: a re-examination of the Procellariiformes. Anim Behav 65:235–246
Burness GP, Morris RD, Bruce JP (1994) Seasonal and annual variation in brood attendance, prey type delivered to chicks, and foraging patterns of male common terns (Sterna hirundo). Can J Zool 72:1243–1251
Coulson JC (1980) A study of factors influencing the duration of the pair-bond in the Kittiwake Gull Rissa tridactyla. In: Nöhring R (ed) Proc Int Ornithol Congr 17:823–833
Coulson JC, Horobin J (1976) The influence of age on the breeding biology and survival of the arctic tern Sterna paradisaea. J Zool 178:247–260
Coulson JC, Thomas CS (1983) Mate choice in the Kittiwake gull. In: Bateson P (ed) Mate choice. Cambridge University Press, Cambridge, pp 361–376
Curio E (1983) Why do young birds reproduce less well? Ibis 125:400–404
Davis JWF (1976) Breeding success and experience in the arctic skua, Stercorarius parasiticus (L.). J Anim Ecol 45:531–535
Dittmann T, Becker PH (2003) Sex, age, experience and condition as factors of arrival date in prospecting common terns Sterna hirundo. Anim Behav 65:981–986
Ens BJ, Choudhury S, Black JM (1996) Mate fidelity and divorce in monogamous birds. In: Black JM (ed) Partnerships in birds – the study of monogamy. Oxford University Press, Oxford, pp 118–137
Forslund P, Larsson K (1992) Age-related reproductive success in the barnacle goose. J Anim Ecol 61:195–204
Forslund P, Pärt T (1995) Age and reproduction in birds. Trends Ecol Evol 10:374–378
Fowler GS (1995) Stages of age-related reproductive success in birds: Simultaneous effects of age, pair-bond duration and reproductive success. Am Zool 35:318–328
Galbraith H, Hatch JJ, Nisbet ICT, Kunz TH (1999) Age-specific reproductive efficiency among breeding common terns Sterna hirundo: measurement of energy expenditure using doubly-labelled water. J Avian Biol 30:85–96
Gonzáles-Solís J, Becker PH, Wendeln H (1999) Divorce and asynchronous arrival in common terns, Sterna hirundo. Anim Behav 58:1123–1129
Gonzáles-Solís J, Sokolov E, Becker PH (2001) Courtship feedings, copulations and paternity in common terns Sterna hirundo. Anim Behav 61:1125–1132
Gonzáles-Solís J, Becker PH, Jover L, Ruiz X (2004) Individual changes underlie age-specific pattern of laying date and egg-size in female common terns Sterna hirundo. J Ornithol 145:129–136
Hamer KC, Furness RW (1991) Age-specific breeding performance and reproductive effort in great skuas Catharacta skua. J Anim Ecol 60:693–704
Harrington BA (1974) Colony visitation behavior and breeding ages of sooty terns (Sterna fuscata). Bird-Banding 45:115–144
Harvey PH, Greenwood PJ, Perrins CM, Martin AR (1979) Breeding success of great tits Parus major in relation to age of male and female parent. Ibis 121:216–219
Holm S (1979) A simple sequentially rejective multiple test procedure. Scand J Stat 6:65–70
Jouventin P, Lequette B, Dobson FS (1999) Age-related mate choice in the wandering albatross. Anim Behav 57:1099–1106
Klomp NI, Furness RW (1991) Recruitment in long-lived birds: studies of non-breeding great skuas. In: Bell BD (ed) Proc Int Ornithol Congr 20:1678–1688
Ludwigs J-D (1998) Kleptoparasitismus bei der Flußseeschwalbe Sterna hirundo als Anzeiger für Nahrungsmangel. Vogelwelt 119:193–203
Ludwigs J-D, Becker PH (2002) The hurdle of recruitment: Influences of arrival date, colony experience and sex in the common tern Sterna hirundo. Ardea 90:389–399
Martin K (1995) Patterns and mechanisms of age-dependent reproduction and survival in birds. Am Zool 35:123–141
Mills JA, Shaw PW (1980) The influence of age on laying date, clutch size, and egg size of the white-fronted tern, Sterna striata. NZ J Zool 7:147–153
Neubauer W (1997) Beziehungen zwischen Paarbindung, Alter und Bruterfolg bei der Flußseeschwalbe Sterna hirundo. Beitr Vogelw Hiddensee 14:37–45
Newton I, Rothery P (1998) Age-related trends in the breeding success of individual female sparrowhawks Accipiter nisus. Ardea 86:21–31
Nielsen JT, Drachmann J (2003) Age-dependent reproductive performance in northern goshawks Accipiter gentilis. Ibis 145:1–8
Nisbet ICT, Winchell JM, Heise AE (1984) Influence of age on the breeding biology of common terns. Col Waterbirds 7:117–126
Nisbet ICT, Apanius V, Friar MS (2002) Breeding performance of very old common terns. J Field Ornithol 73:117–124
Ollason JC, Dunnet GM (1978) Age, experience and other factors affecting the breeding success of the fulmar, Fulmarus glacialis, in Orkney. J Anim Ecol 47:961–976
Ollason JC, Dunnet GM (1988) Variation in breeding success in fulmars. In: Clutton-Brock, TH (ed) Reproductive success. University of Chicago Press, Chicago, pp 268–278
Pärt T (1995) Does breeding experience explain increased reproductive success with age? An experiment. Proc R Soc Lond B 360:113–117
Pyle P, Spear LB, Sydeman WJ, Ainley DG (1991) The effects of experience and age on the breeding performance of western gulls. Auk 108:25–33
Pyle P, Sydeman WJ, Hester M (2001) Effects of age, breeding experience, mate fidelity and site fidelity on the breeding performance in a declining population of Cassin’s auklets. J Anim Ecol 70:1088–1097
Ratcliffe N, Furness RW, Klomp NI (1998) Influences of breeding experience on the reproductive performance of great skuas Catharacta skua. J Avian Biol 29:293–298
Rees EC, Lievesley P, Pettifor RA, Perrins C (1996) Mate fidelity in swans: an interspecific comparison. In: Black JM (ed) Partnerships in birds – the study of monogamy. Oxford University Press, Oxford, pp 118–137
Reid WV (1988) Age correlations within pairs of breeding birds. Auk 105:278–285
Rice WR (1989) Analyzing tables of statistical tests. Evolution 43:223–225
Rowley I (1983) Re-mating in birds. In: Bateson P (ed) Mate choice. Cambridge University Press, Cambridge, pp 331–360
Ryder JP, 1981. The influence of age on the breeding biology of colonial nesting seabirds. In: Burger J, Olla BL, Winn HE (eds) Behaviour of marine animals, vol 4. Plenum Press, New York, pp 153–168
Saether B-E, 1990. Age-specific variation in the reproductive performance of birds. In: Power DM (ed) Current ornithology, vol 7. Plenum Press, New York, pp 251–283
Sasvári L, Hegyi Z (2000) Sex-related local recruitment in colonial and solitary breeding European tree sparrows Passer montanus L. Ibis 142:119–122
Sudmann SR (1998) Wie dicht können Flußseeschwalben Sterna hirundo brüten? Extremsituation auf Brutflößen. Vogelwelt 119:181–192
Village A (1986) Breeding performance of kestrels at Eskdalemuir, south Scotland. J Zool 208:367–378
Weimerskirch H (1992) Reproductive effort in long-lived birds: age-specific patterns of condition, reproduction and survival in the wandering albatross. Oikos 64:464–473
Wendeln H (1997) Allocation of parental duties and foraging behavior influence body condition of adult common terns, Sterna hirundo. Bird Behav 12:47–54.
Wendeln H, Becker PH (1996) Body mass change in breeding common terns (Sterna hirundo). Bird Study 43:85–95
Wendeln H, Becker PH, Gonzáles-Solís J (2000) Parental care of replacement clutches in common terns (Sterna hirundo). Behav Ecol Sociobiol 47:382–392
Wiggins DA (1989) Consequences of variation in brood size on the allocation of parental care in common terns (Sterna hirundo). Can J Zool 67:2311–2413
Wiggins DA, Morris RD (1987) Parental care of the common tern Sterna hirundo. Ibis 129:533–540
Wooller RD, Bradley JS, Skira IJ, Serventy DL (1990) Reproductive success of short-tailed shearwaters Puffinus tenuirostris in relation to their age and breeding experience. J Anim Ecol 59:161–170
Wooller RD, Bradley JS, Croxall JP (1992) Long-term population studies of seabirds. Trends Ecol Evol 7:111–114
Acknowledgments
Our studies would not have been possible without the support of various people and organisations: M. Wagener, H. Wendeln, R. Nagel and many students and helpers in the field studies; T. Dittmann and three anonymous reviewers provided useful advice on how to improve the manuscript, as did P. Prosser, who also checked the English. This study was supported by the Deutsche Forschungsgemeinschaft (BE 916/5), TROVAN Cologne; DA-Electronics, Tecklenburg; BRAUN Melsungen; INEFA Itzehoe, Standortverwaltung and Bundesvermögensamt Wilhelmshaven, and ANITECH, Oldenburg.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by F. Trillmich
Rights and permissions
About this article
Cite this article
Ludwigs, JD., Becker, P.H. What do pairing patterns in common tern, Sterna hirundo, recruits reveal about the significance of sex and breeding experience?. Behav Ecol Sociobiol 57, 412–421 (2005). https://doi.org/10.1007/s00265-004-0880-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-004-0880-8