
Abstract
The study of intraspecific variation can provide insights into the evolution and maintenance of behavior. To evaluate the relative importance of ecological, demographic and social conditions thought to favor lekking, I studied variation in mating behavior among and within populations of the blackbuck, Antilope cervicapra, an Indian antelope. Rather than viewing lekking as a discrete mating strategy, I took a continuous approach and treated lekking as a question of the clustering of mating territories, with leks representing one extreme in a range of territory distributions. I surveyed nine blackbuck populations, which differed in population density and in habitat conditions. For each population, I described the mating system in terms of the clustering of mating territories, and measured various factors suggested to favor lekking. I found that large-scale, among-population variation in territory clustering was most strongly related to female group size. Territory clustering was not related to population density. Female group size, in turn, was best explained by habitat structure. Interestingly, these among-population patterns were repeated at a finer spatial scale within one intensively studied population. These findings suggest that territorial males respond to local patterns in female distribution (represented by group size) when making decisions regarding territory location. Finally, although female distribution may explain territory clustering at the population level and more locally within a population, other selective factors (e.g., female preference, male competition, male harassment) are likely to shape the clustering and size of territories at even finer scales.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Mating systems in which males do not provide any parental care have usually been explained in terms of males monopolizing females or resources attractive to females (Clutton-Brock 1989). From this perspective, the evolution and maintenance of the lek mating system is intriguing because males defend neither females nor any obvious resources. Lekking males typically defend small mating territories in aggregations. These territories are devoid of resources such as food or water, and females are thought to visit leks for the sole purpose of mating (Bradbury 1981). The lek mating system is rare (occurring in less than 0.5% of birds and 0.2% of mammals; Davies 1991), but it is found in a wide diversity of taxa (in at least 14 bird families and 5 mammal families; Höglund and Alatalo 1995). Furthermore, many species that lek also display variation in their mating system. This is especially common among lekking ungulates. For example, in topi (Damaliscus lunatus) and fallow deer (Dama dama), males in some populations lek while males in others show resource defense and female following strategies (Gosling 1991; Thirgood et al. 1999). Male mating behavior may even vary within populations (Clutton-Brock et al. 1988; Gosling and Petrie 1990; Apollonio et al. 1992). Such intraspecific variation offers the opportunity to explore the correlates of male mating behavior and identify the conditions associated with lekking, thereby providing insights into the evolution of this unusual mating system. I studied the blackbuck (Antilope cervicapra), an Indian antelope with highly variable mating behavior, to evaluate the relative importance of ecological, demographic, and social conditions hypothesized to favor lekking.
The main ecological factors suggested to influence lekking are resources, habitat structure, and predation (Wiley 1974; Gosling 1986; Clutton-Brock 1989). Resources and habitat structure are thought to affect lekking by influencing female density and distribution (Gosling 1986; Deutsch 1994; Nefdt and Thirgood 1997). Habitat structure may also constrain mating options available to males, for example, if habitat suitable for mating territories is limiting (Wegge and Rolstad 1986). A third ecological factor, predation, may favor lekking, if territorial males or visiting females experience reduced risk from predation at larger clusters of territories (Koivisto 1965; Wiley 1974).
Demographic factors, principally female density, are also thought to influence lekking. Many studies have found that lekking is associated with relatively high population densities (Langbein and Thirgood 1989; Balmford et al. 1993a, 1993b; Clutton-Brock et al. 1993; Bro-Jørgensen 2003a). Furthermore, there are several accounts of switches in mating system following changes in density (Clutton-Brock et al. 1993). Population sex ratio may also influence lekking (Apollonio 1989).
Two social factors correlate with lekking in several species: the social behavior of females (grouping and ranging), and harassment of estrous females by males. Lekking often occurs in species in which females move unpredictably in large groups and have large home ranges (Bradbury et al. 1986; Balmford et al. 1993a; Clutton-Brock et al. 1993). Male harassment of estrous females is suggested to favor leks in ungulates (Clutton-Brock et al. 1992; Nefdt 1995; Nefdt and Thirgood 1997; but see Bro-Jørgensen 2002, 2003b).
Overall, many ecological, demographic, and social factors have been related to lekking, but studies do not provide a consistent picture. While it is likely that the conditions favoring lekking vary among taxa, evidence for the role of various ecological and demographic factors is mixed even within a species (Apollonio 1989; Balmford et al. 1993a). One possible reason for the lack of clear patterns is that most studies focus on a single population and on testing specific processes thought to maintain lekking in that population. The general conditions associated with lekking are typically assessed in reviews that compile results from different single-population studies, studies that differ in the behavioral patterns and factors measured and in the methods used. Very few studies have systematically examined variation among species or populations of a species in mating system. In one such study (Langbein and Thirgood 1989), the factor that explained most of the variation in mating system among fallow deer populations was male density, a factor not highlighted in reviews of lekking (e.g., Clutton-Brock et al. 1993). Langbein and Thirgood’s (1989) study suggests that cross-population comparisons in species with variable mating behavior provide an important means of identifying the conditions under which lekking may evolve.
I studied variation among and within populations of blackbuck to identify the ecological, demographic, and social conditions associated with lekking. Previous studies have treated lekking as a discrete mating system and have classified populations as lekking or non-lekking (e.g., Apollonio 1989; Balmford et al. 1993a; Nefdt and Thirgood 1997). However, studies of blackbuck (Mungall 1978; Ranjitsinh 1989; Isvaran and Jhala 2000) suggest that this classification conceals much useful variation. Furthermore, most theoretical studies model lekking as a matter of the clustering of territories, and thus use a continuous rather than a categorical approach (Bradbury 1981; Gibson et al. 1990; Stillman et al. 1993, 1996; Isvaran and St. Mary 2003). Hence, rather than classify the mating system of a population into discrete categories, I take a novel empirical approach and describe the mating system in terms of the distribution of mating territories, specifically the degree to which territories are clustered. I surveyed nine blackbuck populations and measured the distribution of territories, and the ecological, demographic, and social factors hypothesized to favor lekking. In addition to making cross-population comparisons, I also studied the correlates of territory clustering at a finer spatial scale, within a lekking population. Here, I examine the factors associated with variation in territory clustering, both among and within populations. I evaluate the correlates of extreme territory clustering (lekking) and discuss the implications of my findings in light of current hypotheses about lek evolution.
Methods
Study organism
The blackbuck, A. cervicapra, is an endangered antelope native to the Indian subcontinent. It is a medium-sized (23–45 kg; Mungall 1978; Ranjitsinh 1989), sexually dimorphic species in which males are larger and more conspicuously colored than females, and males alone bear long, spiral horns. It is found in a wide range of habitats, from grasslands to open woodlands. It is a selective grazer living in groups that range from two to several hundred individuals (Ranjitsinh 1989). The social groups typically observed are: (1) all-male herds; (2) female herds (containing adult females and juveniles of both sexes); (3) mixed-sex herds (containing adults and juveniles of both sexes). There are two annual mating peaks, from March to April and from August to October (Ranjitsinh 1989). Males display to females on mating territories and in mixed-sex herds, but no matings have been observed in herds (Mungall 1978; Prasad 1989; this study).
Study sites
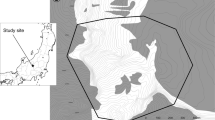
I surveyed eight populations in India and one in Texas, USA, from August to November 1998 and 1999. The Indian populations were scattered throughout the distribution of blackbuck in India and represented a wide range of habitat type, blackbuck density, and predator density (Fig. 1, Table 1, electronic Appendix S1). All eight Indian populations lie in semi-arid regions and habitats range from open grasslands to thorny shrublands and forest. I also studied a free-ranging population on Kyle ranch in Texas (Table 1). Since their introduction from zoos to Texas ranches 70–80 years ago (Mungall 1978), blackbuck numbers have rapidly increased and currently rival their overall numbers in India (Mungall 1998). The Texas population thus provides an opportunity to examine whether the correlates identified for the Indian populations hold in a population introduced into a new habitat. Kyle ranch is located in a semi-arid region and comprises shrublands and forest with small grassy openings.

Location of the eight Indian study populations. Velavadar in western India was the intensive study site.
I studied spatial variation in territory clustering more intensively within one of the Indian populations, Velavadar National Park, Gujarat, from January to May 2000. This area is a mosaic of grasslands, shrublands, and mudflats, and holds one of the largest blackbuck populations in India.
Among-population variation in mating system
Distribution of mating territories
Because most of the study areas were relatively small (1–36 km2), each site was repeatedly surveyed, and the distribution of territorial males was recorded on maps of the site. From these distributions, I identified clusters of territories. A cluster included all males that shared territory boundaries. This was inferred from observing areas used by each territorial male and interactions between neighboring males. Clusters were typically unambiguously identified. For example, at Velavadar, the mean nearest neighbor distance of territorial males within a cluster was 85 m (22 territories in 5 clusters) while the mean distance from a cluster to its nearest neighboring cluster was 900 m (5 clusters).
Correlates of mating system variation
Ecological variables
I divided each study site into four to seven units (depending on the area of the site), and randomly laid a 1-km line transect in each unit. At 50-m intervals along these transects, habitat structure (habitat openness and habitat homogeneity) and resource abundance were measured as described below.
Habitat openness
Because blackbuck prefer open habitats (primarily grasslands; Ranjitsinh 1989; Jhala 1997), the measured habitat-structure variables focused on open habitats. I placed habitats in two categories—open and closed. Open habitats included those without woody shrubs and trees more than 1 m high (mainly grassland and bare ground). Closed habitats were defined as those with woody shrubs and trees more than 1 m high (mainly shrubland and forest). At 50-m intervals along each transect, I visually estimated the percentage of the area occupied by open and closed habitats in circular plots of 10 m radius. Habitat openness was calculated as the mean percentage of open habitat in a plot (estimates were first averaged across plots and then across transects).
Habitat homogeneity
As a measure of the distribution of open habitat patches, for each transect, I counted the number of successive sampling points with open habitat as the major habitat type. Thus, each transect can be broken down into segments of varying lengths (1–20), where each segment consists of successive points with open habitat. These segments are an index of the size of open-habitat patches through which the transect ran. A transect that did not go through any open habitat was assigned a value of zero. A transect with all 20 points falling in open habitat represented maximum homogeneity of open habitat (value of 20). The mean length of segments of open habitat was averaged across transects to obtain a measure of homogeneity of open habitat at each study site.
Resource abundance
At each sampling point along a transect, I measured grass height and visually estimated the percentage of area covered by grass in circular plots of 1 m radius. Grass height×percent cover was used as an index of resource abundance.
Predator density
To test whether territory clustering was correlated with predation levels, I obtained estimates of the number of predators (wolves, Canis lupus) of adult blackbuck known to use each study site from Forest Department records and from information obtained from researchers working at the sites. Using these estimates, I calculated predator density (number of wolves per km2) for each site.
Demographic variables
I used total counts or line transects to estimate: (1) population density; (2) male density (number of males per km2); (3) female density (number of females per km2); and (4) sex ratio (number of females divided by the number of males). Only mature females (~1 year and older) and males (~2 years and older) were included in the calculation of sex ratio (Mungall 1978). To identify mature individuals, I used body proportions for females and body proportions and horn characteristics (primarily the number of twists) for males (Mungall 1978). Fawns were not included in the density and sex ratio measures.
At six sites, total counts (a recommended census technique for conspicuous animals that aggregate and that are found in relatively open areas; Sutherland 1996) were used to estimate demographic parameters. During each total count, I systematically surveyed the study area in the morning and evening hours when animals were most active, and recorded the sex and stage (immature, mature) of every individual encountered. I conducted three to four total counts at each of the six sites. At three sites (Point Calimere, Savainagar and Kyle ranch), total counts were difficult to conduct because of relatively dense vegetation. At these three sites, I walked six to seven 1-km strip transects stratified as described for the measurement of habitat variables. Along each transect, I recorded every animal encountered within 50 m on either side of the transect. The density of individuals was calculated as the total number of individuals sighted divided by the area sampled (the study area for total counts and the area of the strip transect for the transects).
Social variables
Group size
I used the total counts and transects described above to estimate group sizes. I defined a group as all individuals within at least 50 m of another individual (Clutton-Brock et al. 1982; Lingle 2001). During total counts and transects, for each individual or group encountered, I recorded the number, sex, and stage of individuals.
Female group size
Because male mating behavior is hypothesized to be influenced by the degree to which females in particular (rather than all individuals) are aggregated in an area, I took the data on group sizes (recorded as described above), selected only groups that contained females (female groups and mixed-sex groups), and estimated the mean number of females per group.
Female ranging
Female ranging is thought to influence male mating behavior by modifying the rate at which males encounter females. For example, when females range widely over large areas, males are expected to experience low female encounter rates at any given location (Deutsch 1994). Due to the limited time that I spent at each study site, I could not measure female home-range sizes. Instead, I used an index of female ranging, the distance moved during a sampling period. In each population, I followed 10–15 (mean=13, SD=3) female or mixed-sex groups for 30–60 min (mean=40; SD=7) on different days and at different times of the day. During each group follow, I noted the location of the group at the beginning and the end of the follow and measured the straight-line distance between the two locations. This measure (meters moved per hour) was used as an index of female ranging.
Male harassment
During the follows of mixed-sex groups described above, I recorded the number and duration of chases of females by males. I used the number of chases per female per hour as a measure of male harassment of females.
Within-population variation in mating system
I examined the correlates of variation in territory clustering within a primarily lekking population (Velavadar National Park), from January to May 2000. I divided the study area into approximately 1 km2 (1.1×1.1 km) sampling units. The approximate grid size was chosen based on the spatial scale over which factors were expected to vary. The exact grid size was chosen based on the scale of the map obtained from the Forest Department. Every 2 weeks during the rutting peak (March and April), I surveyed the study area three to five times on consecutive days and recorded the location and distribution of territories on maps. From these maps, I estimated the number of territorial males, and cluster sizes (number of males in a territory cluster) in each spatial unit. I also conducted three to five total counts during morning and evening hours and estimated the number of males and females in each 1.2-km2 spatial unit. In each unit, I laid two 500-m transects at random and measured habitat characteristics and resource abundance along these transects as described in the among-population comparison.
Analyses
Among-population variation in territory clustering
The extent of territory clustering in the different populations was estimated in two ways.
Typical cluster size
First, to estimate the cluster size (number of males in a cluster) that the average male was found in, I used the measure proposed for social groups (typical group size; Jarman 1982):
where x is the number of males in a cluster, n is the number of clusters and N is the total number of territorial males. A solitary male was counted as a cluster of one. The typical cluster size is the number of territorial males in the territory cluster that the average territorial male occupies.
Standardized Clustering Index (SCI)
As the total number of territorial males differed among populations, I sought an index of territory clustering that was independent of the number of males in a population. To achieve this, I divided the typical cluster size by the number of territorial males in the population. Thus, the Standardized Clustering Index ranges between near 0 and 1 and represents a relative rather than an absolute measure of territory clustering. For example, using this index, a territory cluster of 10 in a population with 10 territorial males represents the same level of clustering as a cluster of 60 in a population with 60 males. The SCI is based on the null expectation that absolute cluster size increases linearly with the number of territorial males in a population. Note that this expectation may be unreasonable when territorial male populations and/or study areas are very large, problems that were unlikely in this study.
I used parametric correlation analyses to explore the relationship between the extent of territory clustering (typical cluster size and SCI) and the various ecological, demographic, and social factors. To meet the assumptions of bivariate normality and linearity, all variables were loge-transformed except habitat openness and habitat homogeneity. Habitat openness was bounded by 0 and 100 and homogeneity by 0 and 20. They were best treated as proportions and accordingly arcsine square-root transformed (Sokal and Rohlf 1995).
I also used stepwise multiple regression analyses with SCI as the dependent variable to test whether factors identified as correlates (r≥0.5) in bivariate correlation analyses were statistically independent predictors of territory clustering. The independent variables used were habitat homogeneity and female group size. To minimize problems associated with multicollinearity, habitat openness was not included in the regression because it was highly correlated with habitat homogeneity (r=0.97, N=9, P<0.0001). Female ranging, the only other correlate of SCI (at r≥0.5), was not included in the regression analysis because data on ranging could not be collected in all populations. The residuals from regression analyses were checked for deviations from normality and linearity. A similar stepwise regression was performed with typical cluster size as the dependent variable. Because the results from this analysis were very similar to those from the regression analysis of SCI, only the latter are reported here.
Within-population variation in territory clustering
I used typical cluster size to represent the clustering of territories in each 1.2-km2 sampling unit. At this spatial scale, SCI was not appropriate, because there was usually one and not more than three clusters per spatial unit. The relationship between typical cluster size and the various factors measured was explored using parametric correlation analyses. To identify the factor(s) independently explaining variation in typical cluster size, I used stepwise multiple regression analyses with independent variables that showed a relatively large correlation (r≥0.5) with typical cluster size: resource abundance, male numbers and female group size. The only other correlate (at r≥0.5) was female numbers, which I did not include because it was correlated with female group size (r=0.88, N=14, P<0.0001). All analyses were performed on loge-transformed data.
The above analyses assume that data from each sampling unit are independent, and do not take into account potential spatial correlations among sampling units. I incorporated possible nonindependence between sampling units using generalized least squares (GLS) methods (Pinheiro and Bates 2000). Results from GLS models were similar to those from the stepwise regression models ignoring spatial correlations. For this reason, I report results from only the stepwise regression analyses.
Results
Among-population variation in territory clustering
The distribution of territories varied widely among populations (Fig. 2). The typical cluster size, the number of territorial males in the cluster that the average territorial male occupied, ranged from 1.3 males in Vanasthali to 40.4 males in Tal Chappar (see electronic Appendix, S1). The typical cluster size reflects absolute levels of territory clustering and is affected by the total number of territorial males in a population. Hence, I used a relative measure of clustering, SCI, which represents the level of clustering adopted by the average territorial male in a population taking into account differences among populations in the total number of males. Using this index, I found that males in populations with different numbers of territorial males and typical cluster sizes still exhibited a similar relative level of clustering (SCI; see electronic Appendix, S1).

Relative frequency of territorial males in clusters of different sizes in nine blackbuck populations. N represents the number of territorial males in a population.
Of all the factors measured, female group size was most strongly correlated with both SCI and typical cluster size (Table 2). Clustering increased with female group size and extreme clustering (classical leks) was associated with large female groups (Fig. 3). Neither SCI nor typical cluster size was related to harassment. Typical cluster size increased and SCI tended to increase with the ranging of female groups (Table 2).

Relationship between the degree of territory clustering (SCI) and female group size across blackbuck populations. Each data point represents a population.
Of the ecological factors measured, both habitat openness and homogeneity were positively correlated with typical cluster size, but their relationships with SCI were not significant. Territory clustering was not associated with resource abundance (Table 2). I also used annual rainfall as an index of resource abundance (Maher 2000), but did not find a significant correlation between annual rainfall and either SCI (r=−0.39, N=9, P=0.294) or typical cluster size (r=−0.31, N=9, P=0.423). Similarly, predator density was not associated with SCI or typical cluster size (Table 2). Because for several populations I relied on information from secondary sources for predator density estimates, and because such densities are difficult to estimate in the absence of intensive sampling, the error around these estimates is uncertain. Therefore, I also tested for the effect of predator density in a more general manner. I placed populations in Low (wolves absent) and High (wolves present) categories of predation pressure and tested for a difference in SCI between the two categories. SCI did not differ significantly between the two categories (t-test on loge-transformed SCI: t=0.396, df=6, P=0.706). I found no clear relationship between either typical cluster size or SCI and any demographic factor including female density (Table 2).
In the stepwise regression analysis of SCI, only female group size entered the regression and it explained a large portion of the variation in SCI (R2=0.80, N=9, F1,7=27.24, P=0.001). The degree of territory clustering at the Kyle ranch (Texas) population matched expectations based on the relationship between clustering and group size in the Indian populations. Group sizes were relatively small at Kyle ranch and, correspondingly, the level of territory clustering was relatively low (see electronic Appendix, S1, and Fig. 3).
The main predictor of female group size was habitat homogeneity. In a stepwise regression with loge-transformed female group size as the dependent variable and habitat homogeneity and resource abundance as independent variables, only habitat homogeneity entered the regression (R2=0.62, N=9, F1,7=11.51, P=0.012). Because habitat homogeneity was strongly correlated with habitat openness, only the former was included in the multiple regression to minimize problems associated with multicollinearity.
Within-population variation in territory clustering
At Velavadar, where I studied the correlates of territory clustering within a population, cluster sizes varied from solitary territories and small clusters of 2–4 males to a lek of 50 males. The distribution of territories among the 1.2-km2 sampling units was related to female distribution. The number of territorial males in a unit was positively correlated with the number of females in a unit (r=0.75, N=14, P=0.002, Fig. 4).

Relationship between the number of territorial males and the number of females among sampling units within the Velavadar population. Each data point represents a sampling unit.
Territories were observed in 8 of the 14 sampling units at Velavadar. Variation in territory clustering (measured as typical cluster size) among these eight units was correlated with variation in mean female group size, number of females observed in each unit, and male numbers (Table 3). In a stepwise regression analysis of typical cluster size, female group size was the only variable that was included in the regression (R2=0.85, N=8, F1,6=34.83, P=0.001) (Fig. 5).

Relationship between the typical cluster size (degree of clustering of territorial males) and female group size among sampling units within the Velavadar population. Each data point represents a sampling unit.
Discussion
Patterns of variation in territory clustering
A striking result of this study was the wide variation among populations in mating system, specifically in the degree to which mating territories were clustered. Two populations (Velavadar and Tal Chappar) displayed extreme clustering, with most males aggregating on large classical leks, while males in one population (Vanasthali) defended large scattered territories in foraging areas used by females (resource territories). The remaining six populations displayed different degrees of intermediate clustering of territories; this ranged from populations in which males were found in several small territory clusters to those in which males defended territories in a range of small and moderately sized clusters (2–18 males). Furthermore, territory sizes ranged widely from 350 m2 to 100,000 m2 as did abundance of forage on territories (Isvaran 2003). Due to this variation, the mating system in the majority of blackbuck populations could not be easily placed in discrete categories (e.g., resource defense territoriality and lekking). These findings suggest that classical lekking is one extreme in a wide range of territorial behavior. Understanding lekking is thus embedded in the larger question of understanding the processes that lead to variation in territorial behavior.
Such variation in territory clustering has also been reported in groups of closely related species (e.g. grouse species, Hjorth 1970; Wiley 1974; Lewis 1985), and within some ungulate species (e.g., fallow deer, Thirgood et al. 1999; topi, Gosling 1991). Furthermore, several studies (e.g., black lechwe Kobus leche smithemani, Thirgood et al. 1992; little bustard Tetrax tetrax, Jiguet et al. 2000; grassquit Volatinia jacarina, Almeida and Macedo 2001) also describe intermediate levels of territory clustering that are difficult to categorize. Such variation suggests that the difference between many territorial distributions (e.g., dispersed leks vs classical leks) is continuous rather than discrete (Höglund and Alatalo 1995). While this variation makes the placement of observed mating systems into currently recognized categories problematic, it provides the opportunity to study the conditions that influence mating-system variation and that favor unusual mating systems such as lekking.
Correlates of territory clustering from among-population comparisons
A second striking result of this study was that female group size was the principal correlate of among-population variation in male territory clustering. In populations in which females were in small groups, males defended territories either alone or in small clusters. At the other extreme, classical leks were associated with large groups of females. Although few previous studies have examined continuous variation in territory clustering, comparisons of lekking and non-lekking mating systems identify large group size as one of the three main conditions associated with lekking (Wiley 1974; Bradbury et al. 1986; Davies 1991; Clutton-Brock et al. 1993). Thus, results from blackbuck provide strong quantitative support to previous observations on the relationship between female group size and mating system. The main explanation for the association between large female groups and lekking is that other male mating strategies, such as the defense of female groups or of resources that attract females, are not economical when females occur in large unstable groups and local numbers of females are high (Davies 1991; Clutton-Brock et al. 1993). This argument, however, does not address why, under such conditions, the payoffs from lekking are higher than those from other mating strategies. A possible alternative interpretation of the correlation between female group size and clustering is that females aggregate in large groups in response to male territory clustering. However, this is unlikely in blackbuck because large female groups are present throughout the year, even during times of low territorial activity.
A second important condition thought to influence mating system is population density. Lekking may be favored in high-density populations because when local numbers of females are high, clusters of males may attract enough females to offset the large costs of clustering (Clutton-Brock et al. 1988). An association between density and lekking has been reported in most lekking ungulates (Langbein and Thirgood 1989; Balmford 1992; Balmford et al. 1993a, 1993b). Surprisingly, I found no relationship between population density and territory clustering in blackbuck.
Why might female group size rather than population density be associated with territory clustering in blackbuck? Population density is thought to influence lekking through large local numbers of females. However, in blackbuck, female group size is likely a better estimate of local numbers of females than is overall population density. Groups show some overlap in home ranges, but within a season this overlap is usually limited (e.g., 5–30% in blackbuck, Prasad 1983). Furthermore, females in most ungulates rarely leave their daily ranges to mate and a previous study of lekking antelope found that most estrous females sample mates within their usual home ranges (Balmford et al. 1992). Hence, female group sizes, in effect, represent the number of estrous females available at a local spatial scale. Overall population density, however, may not always represent local distribution of females, especially in heterogeneous environments (Apollonio 1989). Indeed, I found no relationship between population density and local density (represented by group size).
Because population density and group size are not correlated in blackbuck (unlike in many other ungulate populations, Clutton-Brock et al. 1993), this allows one to analyze their relative influence on male behavior. My findings suggest that male territorial behavior is more influenced by local patterns in female distribution (represented by female group size), than by overall population patterns. Apollonio (1989) arrived at a similar conclusion for fallow deer in Italy where, unlike in England, fallow deer lek even when population densities are relatively moderate.
A third important characteristic thought to be associated with lekking is large home-range size of females (Bradbury 1981; Davies 1991; Balmford et al. 1993a; Clutton-Brock et al. 1993). When females have large ranges, other more common mating strategies, such as the defense of resources consistently used by females, may no longer be economical (Davies 1991; Gosling 1991; Clutton-Brock et al. 1993). Across blackbuck populations, I found that the degree of territory clustering was only weakly related to female ranging. It is possible that the measure I used is an insufficient index of female ranging. Alternatively, female group size may have a stronger influence on male mating behavior than female ranging; to my knowledge, the relative influence of group size, ranging, and density has not been previously evaluated.
Female group size, the main correlate of mating system variation, was in turn most closely related to habitat openness and homogeneity of open habitat. These habitat features were weakly related to territory clustering with large classical leks tending to be found in populations where large grasslands predominated. These findings support the notion that habitat features do not act directly on male mating behavior, but influence male behavior through their effects on females (Wiley 1974; Gosling 1986; Höglund and Alatalo 1995; Thirgood et al. 1999). Because of the reduced statistical power associated with a sample size of nine populations, more work is needed on the factors that did not show clear relationships with territory clustering in this study.
Patterns and correlates of territory clustering from within-population comparisons
Within the Velavadar blackbuck population, variation in male clustering was largely explained by local female distribution. This result is consistent with findings from the among-population comparison. Furthermore, this result suggests that males respond flexibly to variation in female distribution at small spatial scales. The number of territories in a sampling unit was positively correlated with the number of females. Furthermore, the degree to which these territories were clustered was correlated with both local female numbers and with female group size.
I also found that female group size was a good index of local female distribution, supporting the idea presented in the among-population comparison. At the local spatial scale at which the within-population study was conducted, local female numbers and female group size were strongly correlated.
How might female group size influence territory clustering? Most hypotheses in the literature do not directly explain how large group sizes favor lekking but instead focus on how large female groups reduce the payoffs to mating strategies alternative to lekking. One possible explanation is that local numbers of females act as a ceiling on clustering because they reflect the maximum potential mating benefits to males from clustering (Clutton-Brock et al. 1988). Studies are needed of the ways in which payoffs to male and female mating behavior vary under different group size conditions.
While local female distribution may predict territorial male distribution and clustering at the spatial scale of approximately a square kilometer, this factor is insufficient to explain territory clustering at still finer scales. Within each sampling unit, territory clusters occupied a very small part of the area used by females. For example, in the area of highest local female numbers (280 females km−2), males did not defend dispersed territories covering the whole area; instead they defended small territories, often less than 20 m in diameter, in a single classical lek that covered only a small portion of the area. Studies of sage grouse (Bradbury et al. 1989) and topi (Bro-Jørgensen 2003a) similarly report that local female density explains male dispersion only at coarse spatial scales; they suggest that other selective factors, such as female preference, explain clustering at finer scales.
Implications for different hypotheses of lek evolution
My results support some forms of the hotspot hypothesis, which proposes that males establish territories in areas of maximal overlap of female ranges (Bradbury et al. 1986). The first explicit model of this process (Bradbury et al. 1986) predicted that male clustering should increase with female home-range size and decrease with female density. These predictions are not well supported in blackbuck because my results weakly support the first prediction about home ranges, but not the second concerning female density. Variations on the original hotspot hypothesis propose that females cluster on environmental hotspots and males in turn cluster their territories in accordance with the distribution of females (e.g., Gosling and Petrie 1990). Accordingly, clustering is predicted to increase with local female density. This prediction is supported at relatively coarse spatial scales in blackbuck, by the correlation between female group size and clustering. However, at finer scales (within the sampling units), other factors must be invoked to explain why territorial males occupied only a small part of the area used by females.
Results from blackbuck are also consistent with the female preference hypothesis, which proposes that males cluster their territories because females prefer to mate with clustered males (Bradbury 1981). For such a female preference to favor territory clustering, high local numbers of females are essential. This is because the number of estrous females in an area limits the mating benefits to males holding territories in that area. When local female numbers are low, the benefits to clustered males from female mating preferences are unlikely to offset the costs of defending a territory in a large cluster. Thus, clustering is predicted to increase with local female numbers, a prediction supported by results from blackbuck.
Harassment of estrous females by males has been highlighted as a factor favoring lekking, particularly in species with relatively low mobility such as ungulates (Clutton-Brock et al. 1993; Nefdt and Thirgood 1997). When harassment levels are high, males in clustered territories may be able to monopolize females better than are males in solitary territories and mixed-sex herds. Furthermore, harassment avoidance may be an important benefit favoring female preference for mating on leks rather than in solitary territories or herds (Nefdt and Thirgood 1997). One prediction of these hypotheses is that extreme territory clustering should be associated with high harassment rates in mixed-sex herds in the population (Nefdt and Thirgood 1997). One might also expect territory clustering to be associated with more male-biased sex ratios, because harassment levels likely increase with an increase in male numbers relative to female numbers. These predictions were not supported in blackbuck because neither harassment rates in mixed-sex herds nor sex ratio [both (mature females:mature males) and (mature females: all males)] was correlated with the level of clustering in a population. One possible reason for the lack of pattern in harassment rates is that females move out of mixed-sex herds and remain on territories throughout their estrous period and thus avoid harassment by males in mixed-sex herds; this is unlikely in blackbuck (K. Isvaran, unpublished data). In contrast to findings from lekking fallow deer (Clutton-Brock et al. 1992) and Kafue lechwe (K. leche kafuensis, Nefdt 1995), studies of topi (Bro-Jørgensen 2002, 2003b) and Uganda kob (K. kob thomasi, Balmford 1990) report that harassment cannot explain mating patterns in these species. The results from blackbuck together with results from topi and kob suggest that the role of harassment in ungulate lek evolution needs to be investigated further.
Conclusions
Female group size explained most of the variation in male territory clustering both among populations and, at a smaller spatial scale, within a population. My findings suggest that territorial males respond to local patterns in female distribution when making decisions regarding territory location. One explanation for this male response is that local female numbers may influence clustering by placing a limit on potential mating benefits that males gain from clustering their territories in an area. Studies are needed of the ways in which payoffs to male and female mating behavior vary under different group size conditions. Finally, while female distribution can largely explain male territorial behavior at relatively coarse spatial scales, other selective factors are likely to shape the size and clustering of territories at even finer scales.
More generally, the observed variation in territory distribution in blackbuck, along with evidence from other species (Gosling 1991; Thirgood et al. 1999), suggest that a continuous approach may be preferable to more common categorical approaches to studying lekking. First, separating variable behavior into a few mating system categories can be a matter of judgment. It is likely that intermediate mating behavior that shares features with more than one mating system will, to an extent, be arbitrarily placed in one or another mating system category. Second, a continuous approach allows one to estimate the shape and magnitude of the relationship between selective factors and territory clustering, which may provide insights into evolutionary processes.
References
Almeida JB, Macedo RH (2001) Lek-like mating system of the monogamous blue-black grassquit. Auk 118:404–411
Anonymous (1999) Climatological tables of observatories in India, 1951–1980. Government of India
Anonymous (2002) Comparative climatic data for the United States [1971–2000]. Southern Regional Climate Center
Apollonio M (1989) Lekking in fallow deer—just a matter of density? Ethol Ecol Evol 1:291–294
Apollonio M, Festa-Bianchet M, Mari F, Mattioli S, Sarno B (1992) To lek or not to lek—mating strategies of male fallow deer. Behav Ecol 3:25–31
Balmford A (1990) Lekking in Uganda kob. PhD Thesis. University of Cambridge
Balmford A (1992) Social dispersion and lekking in Uganda kob. Behaviour 120:177–191
Balmford A, Albon S, Blakeman S (1992) Correlates of male mating success and female choice in a lek-breeding antelope. Behav Ecol 3:112–123
Balmford A, Deutsch JC, Nefdt RJC, Clutton-Brock TH (1993a) Testing hotspot models of lek evolution—data from 3 species of ungulates. Behav Ecol Sociobiol 33:57–65
Balmford A, Bartos L, Brotherton P, Herrmann H, Lancingerova J, Mika J, Zeeb U (1993b) When to stop lekking: density-related variation in the rutting behaviour of sika deer. J Zool 231:652–656
Bradbury J (1981) The evolution of leks. In: Alexander RD, Tinkle TW (eds) Natural selection and social behavior. Carron, New York, pp 138–169
Bradbury J, Gibson R, Tsai IM (1986) Hotspots and the dispersion of leks. Anim Behav 34:1694–1709
Bradbury JW, Gibson RM, McCarthy CE, Vehrencamp SL (1989) Dispersion of displaying male sage grouse.2. The role of female dispersion. Behav Ecol Sociobiol 24:15–24
Bro-Jørgensen J (2002) Overt female mate competition and preference for central males in a lekking antelope. Proc Natl Acad Sci USA 99:9290–9293
Bro-Jørgensen J (2003a) The significance of hotspots to lekking topi antelopes (Damaliscus lunatus). Behav Ecol Sociobiol 53:324–331
Bro-Jørgensen J (2003b) No peace for estrous topi cows on leks. Behav Ecol 14:521–525
Clutton-Brock TH (1989) Mammalian mating systems. Proc R Soc Lond B 236:339–372
Clutton-Brock TH, Guinness FE, Albon SD (1982) Red deer: behavior and ecology of two sexes. Edinburgh University Press, Edinburgh
Clutton-Brock TH, Green D, Hiraiwa-Hasegawa M, Albon SD (1988) Passing the buck—resource defense, lek breeding and mate choice in fallow deer. Behav Ecol Sociobiol 23:281–296
Clutton-Brock TH, Price OF, Maccoll ADC (1992) Mate retention, harassment, and the evolution of ungulate leks. Behav Ecol 3:234–242
Clutton-Brock TH, Deutsch JC, Nefdt RJC (1993) The evolution of ungulate leks. Anim Behav 46:1121–1138
Davies NB (1991) Mating systems. In: Krebs JR, Davies NB (eds) Behavioural ecology, 3rd edn. Blackwell, Oxford
Deutsch JC (1994) Lekking by default—female habitat preferences and male strategies in Uganda kob. J Anim Ecol 63:101–115
Gibson RM, Taylor CE, Jefferson DR (1990) Lek formation by female choice: a simulation study. Behav Ecol 1:36–42
Gosling LM (1986) The evolution of the mating strategies in male antelopes. In: Rubenstein DI, Wrangham RW (eds) Ecological aspects of social evolution: birds and mammals. Princeton University Press, Princeton, pp 244–281
Gosling LM (1991) The alternative mating strategies of male topi, Damaliscus lunatus. Appl Anim Behav Sci 29:107–119
Gosling LM, Petrie M (1990) Lekking in topi—a consequence of satellite behavior by small males at hotspots. Anim Behav 40:272–287
Hjorth I (1970) Reproductive behaviour in Tetraonidae. Swed Wildl 7:190–596
Höglund J, Alatalo RV (1995) Leks. Princeton University Press, Princeton
Isvaran K (2003) The evolution of lekking: insights from a species with a flexible mating system. PhD Thesis, University of Florida, Gainesville
Isvaran K, Jhala Y (2000) Variation in lekking costs in blackbuck (Antilope cervicapra): relationship to lek-territory location and female mating patterns. Behaviour 137:547–563
Isvaran K, St. Mary CM (2003) When should males lek? Insights from a dynamic state variable model. Behav Ecol 14:876–886
Jarman PJ (1982) Prospects for interspecific comparison in sociobiology. In: King’s College Sociobiology group (eds) Current problems in sociobiology. Cambridge University Press, Cambridge, pp 323–342
Jhala YV (1997) Seasonal effects on the nutritional ecology of blackbuck Antelope cervicapra. J Appl Ecol 34:1348–1358
Jiguet F, Arroyo B, Bretagnolle V (2000) Lek mating systems: a case study in the little bustard Tetrax tetrax. Behav Proc 51:63–82
Koivisto I (1965) Behavior of the black grouse, Lyrurus tetrix, during spring display. Finn Game Res 26:1–60
Langbein J, Thirgood SJ (1989) Variation in mating systems of fallow deer (Dama dama) in relation to ecology. Ethology 83:195–214
Lewis RA (1985) Do blue grouse form leks? Auk 102:180–184
Lingle S (2001) Anti-predator strategies and grouping patterns in white-tailed deer and mule deer. Ethology 107:295–314
Maher CR (2000) Quantitative variation in ecological and hormonal variables correlates with spatial organization of pronghorn (Antilocapra americana) males. Behav Ecol Sociobiol 47:327–338
Mungall EC (1978) The Indian blackbuck antelope: a Texas view. Kleberg Studies in Natural Resources, College Station
Mungall EC (1998) Bucks in the black: India vs. Texas. Exotic Wildl 8:1–3
Nefdt RJC (1995) Disruption of matings, harassment and lek-breeding in Kafue lechwe antelope. Anim Behav 49:219–429
Nefdt RJC, Thirgood SJ (1997) Lekking, resource defense, and harassment in two subspecies of lechwe antelope. Behav Ecol 8:1–9
Pinheiro JC, Bates DM (2000) Mixed-effects models in S and S-PLUS. Statistics and Computing Series, Springer, Berlin Heidelberg New York
Prasad NLNS (1983) Home range size of blackbuck, Antilope cervicapra, at Mudmal. Z Saeugetierkd 48:109–117
Prasad NLNS (1989) Territoriality in the Indian blackbuck, Antilope cervicapra (Linnaeus). J Bombay Nat Hist Soc 86:187–193
Ranjitsinh MK (1989) The Indian blackbuck. Natraj, Dehradun
Sokal RR, Rohlf FJ (1995) Biometry: The principles and practice of statistics in biological research. Freeman, Oxford
Stillman RA, Clutton-Brock TH, Sutherland WJ (1993) Black-holes, mate retention, and the evolution of ungulate leks. Behav Ecol 4:1–6
Stillman RA, Deutsch JC, Clutton-Brock TH, Sutherland WJ (1996) Black hole models of ungulate lek size and distribution. Anim Behav 52:891–902
Sutherland WJ (1996) Mammals. In: Sutherland WJ (ed) Ecological census techniques: a handbook. Cambridge University Press, Cambridge, pp 260–278
Thirgood SJ, Robertson A, Jarvis AM, Belbin SV, Robertson D, Nefdt RJ, Kamweneshe B (1992) Mating system and ecology of black lechwe (Kobus, Bovidae) in Zambia. J Zool 228:155–172
Thirgood S, Langbein J, Putman RJ (1999) Intraspecific variation in ungulate mating strategies: the case of the flexible fallow deer. Adv Stud Behav 28:333–361
Wegge P, Rolstad J (1986) Size and spacing of capercaillie leks in relation to social behavior and habitat. Behav Ecol Sociobiol 19:401–408
Wiley RH (1974) Evolution of social organization and life-history patterns among grouse. Q Rev Biol 49:201–227
Acknowledgements
I am very grateful to J. Brockmann, C. St. Mary, Y.V. Jhala, C. Chapman, M. Sunquist, J. Eisenberg, B. Bolker, and S. Quader for useful discussions and valuable comments on previous versions of the manuscript; the University of Florida, the Animal Behavior Society, the American Society for Mammalogists, and Sigma Xi for research funds; the Forest Departments of Andhra Pradesh, Gujarat, Maharashtra, Rajasthan, and Tamil Nadu in India for considerable logistical support and kind permission to work in various protected areas; Y.V. Jhala, the USFWS funded project titled Conservation of the Indian Wolf, and the Wildlife Institute of India for immense logistical help; K.V.R. Priyadarshini and B. Jethva for useful discussions and help in the field; E. Mungall, W. Kyle and J. Kyle for extensive support and help with the Texas population; and T. Czeschlik and three anonymous reviewers for constructive comments on the manuscript. This work complies with the current laws of the countries in which the work was carried out.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by T. Czeschlik
Electronic Supplementary Material
Appendix
Appendix
Table 4 is the Electronic Appendix, S1, and gives estimates (mean±SE) of territory clustering and ecological, demographic and social variables in nine blackbuck populations.
Rights and permissions
About this article
Cite this article
Isvaran, K. Female grouping best predicts lekking in blackbuck (Antilope cervicapra). Behav Ecol Sociobiol 57, 283–294 (2005). https://doi.org/10.1007/s00265-004-0844-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-004-0844-z