
Abstract
Fluctuating asymmetries are subtle random deviations from perfect symmetry in bilateral traits caused by the inability of individuals to buffer their development against the disruptive effects of genetic and environmental stresses. The degree of asymmetry of secondary sexual characters is supposed to convey information about a male's phenotypic and/or genetic quality, and females may thus be expected to use bilateral symmetry as a cue in mate choice. We independently and simultaneously manipulated the length and the relative symmetry of the pelvic spines of computer-animated three-spined sticklebacks, Gasterosteus aculeatus. In all mate choice tests involving two male models differing in spine symmetry, females preferred the symmetric model, irrespective of spine length, though only significantly so when both male models had short spines. Females did not significantly discriminate between male models differing in spine length alone. The preference for symmetry was stronger the higher the females' physical condition. Our study demonstrates that female sticklebacks master the perceptual abilities required to discriminate between potential mates basing exclusively on small length differences between paired structures, and provides experimental evidence that spine symmetry is subject to sexual selection through female choice in this species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Fluctuating asymmetry (FA) is a common measure of developmental stability defined as small random deviations from bilateral symmetry in a morphological character for which differences between the right and left sides are normally distributed around a mean of zero in a population (Ludwig 1932; Van Valen 1962; Palmer and Strobeck 1986). The claim that FA is often negatively related to fitness parameters has led to the hypothesis that FA is used in signalling contexts as a surrogate measure of individual quality (Møller 1990), reflecting the ability to cope with genetic and environmental perturbations during development. Owing to a recent history of directional selection and condition-dependent expression, sexual characters are supposed to be particularly sensitive to the effects of stress, and their FA therefore especially suited as a predictor of individual performance (Møller and Pomiankowski 1993a; Watson and Thornhill 1994; but see Bjorksten et al. 2000a, 2000b). Females may thus be expected to use bilateral symmetry as a criterion in mate choice, thereby benefiting in terms of direct and/or indirect fitness advantages. In the last few years, numerous observational studies have lent support to this idea (Møller and Thornhill 1998). However, they do not answer the question of whether females actually use symmetry per se for assessment of male quality, or whether symmetry is simply correlated with other markers used in mate choice decisions. In order to infer causality, experimental manipulation of trait asymmetry independently of other confounding variables is essential. Reported findings of manipulative experiments are often open to alternative explanations, since the manipulation of asymmetry either simultaneously affected other potential aspects of quality (e.g. flight performance), resulted in differences far exceeding those naturally occurring, or was potentially confounded by interactions (Møller 1992; Swaddle and Cuthill 1994; Bennett et al. 1996; Morris and Casey 1998). These pitfalls can be overcome by using computer animations, an innovative non-interactive test paradigm that allows a rigorously standardised and repeatable presentation of stimuli differing exclusively in the investigated trait(s) (see Rosenthal 1999, 2000; Oliveira et al. 2000 and references therein for reviews of the advantages and pitfalls of video playback techniques in behavioural research).
Pelvic spines are one of several bilateral structures (e.g. pectoral fins, armour plates) whose symmetry is expected to be favoured by natural and/or sexual selection in sticklebacks. Male three-spined sticklebacks, Gasterosteus aculeatus, often erect their pelvic spines during courtship (Symons 1965), and the characteristic zig-zag dance performed by courting males may act as an amplifier (Hasson 1989, 1990) giving females the opportunity to assess both sides of a bilateral trait. Field data suggest that pelvic spines are a sexually selected trait in three-spined sticklebacks: spine symmetry (but not spine length) is significantly positively associated with reproductive success measured in terms of the number of eggs present in the nest of caring males, and males with longer spines exhibit lower degrees of FA in spine length (T.C.M. Bakker, D. Mazzi, S.B.M. Kraak, unpublished data), a predicted pattern for secondary sexual characters when assuming that high-quality individuals can afford to develop both large and symmetric structures (Møller 1990; Manning and Hartley 1991; but see also Hunt and Simmons 1997, 1998).
Manipulation of computer-animated models allowed us to offer females a choice between two courting virtual males differing either in relative spine symmetry, spine length, or in both these traits, and to test whether females discriminate between potential mates on the basis of the size and/or the relative symmetry of a bilateral morphological trait when all other male characteristics are held equal.
Methods
Females (ca. 250 in total) were collected during the 2000 spring migration on the island of Texel (The Netherlands) and transported to the facilities of the University of Bern in Hinterkappelen (Switzerland). They were housed in groups of six to eight individuals in 20 l plastic aquaria (water temperature 14±1°C, 16:8 h light:dark regime) and fed to satiation with frozen chironomid larvae or frozen brine shrimps (Artemia) 3 times a day to promote egg maturation.
Ripe females, as assessed by the distension of the abdomen and the opening of the cloaca, were selected daily for the preference tests. They were hand-netted and gently released in a 1-l container (12×7×17 cm, water level: 8.6 cm, corresponding to the height of the test movies displayed on the computer screen) made of clear polyacryl plastic with all the inner walls painted dull grey but for a spare front window (9.5×8 cm). Females were transferred to the test room (room temperature 18±1°C), where they were allowed 30 min of acclimatisation in a pre-test compartment (to account for the logistically imposed difference in temperature between the holding room and the test room), without computer display, but with the same lightning conditions as in the test chamber. Illumination was provided by a fluorescent tube (18 W/33) mounted 35 cm over the female container and dimmed with sheets of white paper. After acclimatisation, the test tank with the female was put in front of the computer screen displaying a still empty scene of a marbled brown mud layer on a green-yellowish background, simulating the natural breeding ground. The empty scene was shown for 5 min for the test females to become accustomed to the test situation. The female was then shown a still image featuring the two males in the back of the scene, at their nest positions. As the males were oriented sideways relative to the choosing female, with their spines retracted, the female was unable to make out any difference between the stimulus males at this time. A grey opaque PVC partition between the middle of the female container and the computer display prevented the female from seeing the left half of the scene when staying in the right half of the container and vice versa. Trials were viewed remotely via a video camera mounted above the test container connected to a monitor outside the test chamber. On the monitor, pieces of tape divided the top-view of the female container in two (right and left) thin front sections measuring 4.2×0.7 cm each, and a neutral back section. Movie playback began after the female had spent at least 2 s in each front section and had regained the neutral section. All females entered the three sections within 2 min of first males' presentation.
Females were given a choice between two identical copies (mirror-images produced by horizontally flipping the original) of a courting stickleback male model differing either in spine length, relative spine symmetry, or in both these traits. The construction of the 3D model and its animation are extensively detailed in Künzler and Bakker (1998). Briefly, a representative male three-spined stickleback was poisoned, fixated and cast with epoxy resin. The casting was cut into thin slices, scanned and digitized. Vectorized outlines were imported in a 3D program and lined up to the body's length axis. A coloured texture was wrapped around the body outline. Spines and eyes were constructed and attached to the body at the appropriate positions, along with the scanned fins. The model's motion pattern was based upon the path of a courting real male video-recorded from above. A standard courtship sequence, including a sequence featuring fanning bouts at the nest, was produced, and a test movie consisting of variable repeats of the different parts of the standard sequence (e.g. zig-zagging towards the front of the scene, swimming back to the nest, fanning) was composed. For the present study, the model described in Künzler and Bakker (1998) had its pelvic spines animated so that it raised and retracted them continuously while courting. The dorsal spines moved synchronously. The models had their spines raised at each turning point between zigs and zags. Males regularly erect their spines at the angular turns of their courtship path, as inferred from video-recordings of live courting males. The courtship sequence was altered so that the males faced the choosing female at the stops between zigs and zags, i.e. they were oriented towards the centre of their respective screen half rather than towards the centre of the screen (cf. Künzler and Bakker 1998). Also, the males spent slightly more time at the front-most position, compared with the models used in Künzler and Bakker's (1998, 2001) studies (10 frames longer, corresponding to a third of a second at a presentation rate of 30 frames per second, extending the whole clip duration by 5 s, from 2 min 20 s to 2 min 25 s).
Starting from the original male model with symmetric spines of average length (Künzler and Bakker 1998, 2001), four different variants were created (Fig. 1). The symmetric long male had both spines elongated by 10%. The symmetric short male had both spines shortened by 10%. The asymmetric long male had the left spine increased by 15.5% and the right spine increased by 4.5%. The asymmetric short male had the left spine reduced by 5.5% and the right spine reduced by 14.5%.

The four combinations of spine length and spine symmetry in computer-animated three-spined sticklebacks, Gasterosteus aculeatus, used in the choice tests
The relative length differences of 20% and relative asymmetry values of 10% obtained by such scaling fall in the range of natural variation observed in this population. An asymmetry of 10% of trait length is large though, and corresponds to the extreme values found in a sample of wild males from the Texel population collected in spring 1998 (n=45, mean±SD R−L=−0.039±0.344, with two individuals exhibiting asymmetries as large as 10% of spine length or larger; D. Mazzi, C.R. Largiadèr, T.C.M. Bakker, unpublished data). We found no evidence of sexual dimorphism in either spine length [ANCOVA on the regression of log10(average spine length) on log10(standard body length), slopes: F 1,81=0.87, P>0.3, intercepts: F 1,82=0.31, P>0.5)] or relative spine asymmetry (F 1,83=0.17, P>0.8) in a wild sample of 45 males and 40 females from the Texel population in 1998 (D. Mazzi, C.R. Largiadèr, T.C.M. Bakker, unpublished data).
Of the six possible pairwise combinations of male models, two involved a difference in relative spine symmetry (whereby either both males had long spines or both males had short spines), two a difference in spine length (whereby either both males had symmetric spines or both males had asymmetric spines), and two a difference in both traits (a male with symmetric long spines against a male with asymmetric short spines, and a male with symmetric short spines against a male with asymmetric long spines).
Test movies were randomly assigned, and male positions were alternated between tests within the six treatments. All females were used only once. A condition factor was calculated as 100 × body mass(g) / standard body length(cm)2.95, whereby the exponent in the formula is the slope of the regression of log10(mass) on log10(length) (Bolger and Connolly 1989). The measure of condition is unconfounded by egg mass, as females were weighed immediately after having spawned. The choice trials were recorded from above, and the recordings analysed a posteriori to avoid observer bias. From the time a female spent in the two front sections (see above), a preference index was calculated as the proportion of time spent oriented towards the preferred male. The arrangement of the set-up hardly allowed observations of specific female behaviours directed to the males, though occasionally females were seen assuming the "head-up" posture signalling spawning readiness (ter Pelkwijk and Tinbergen 1937), and two females (omitted from analyses) actually spawned in the test container. However, to ensure sexual motivation, rather than, for example, a mere schooling preference, only females that spawned with a live male within 24 h of the test were considered.
For analyses, preference scores were angular transformed [arcsin (√x)] to meet the requirements of parametric statistics. We performed a series of t-tests to test for the deviation from the null expectation of no preference. For the preference tests for single traits, as well as for one of the tests involving both traits, we could formulate a priori predictions about which male should be preferred (long over short, symmetric over asymmetric). In these cases a (non-transformed) preference index above 0.5 indicates a preference in the expected direction, below 0.5 a preference in the non-expected direction. As for the one "contradictory" combination of traits (long asymmetric versus short symmetric), a (non-transformed) preference index above 0.5 is arbitrarily set to mean a preference for the male with short symmetric spines.
Stronger preference for the a priori high-quality male when indicated by two "concordant" traits (long symmetric versus short asymmetric) compared to tests of single traits (either symmetry or length) were predicted, while the weakest preference was expected for the "contradictory" combination of traits (long asymmetric versus short symmetric). The strength of the preference expressed by females in the six choice tests was compared by using an ordered heterogeneity test (Rice and Gaines 1994), thus incorporating the information about an anticipated directional pattern into the conventional heterogeneity test.
Given test probabilities are two-tailed throughout. Analyses were performed using JMP IN, v. 4.0.3 (SAS Institute 2000) statistical package.
Results
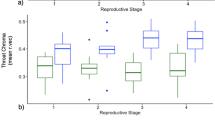
In all choice tests involving two male models differing in the relative symmetry of their pelvic spines, females spent more time oriented to the symmetric male model as opposed to the asymmetric one, regardless of spine length (Fig. 2). However, female preference for symmetry was statistically significant at the α=0.05 level only when both male models had short spines (t 29=2.74, n=30, P=0.011), while the response was non-significant when both models had long spines (t 29=1.85, n=30, P=0.075), and when symmetry and length both were available as discriminatory criteria (t 29=1.97, n=30, P=0.058 for the "concordant" combination of traits opposing a male with symmetric long spines to a male with asymmetric short spines, and t 29=1.99, n=30, P=0.056 for the "contradictory" combination of traits opposing a male with asymmetric long spines to a male with symmetric short spines, respectively). Females did not significantly discriminate between male models differing exclusively in spine length (t 29=0.63, n=30, P>0.5 and t 29=0.39, n=30, P>0.6 for symmetric and asymmetric spines, respectively). When the two treatments where only relative spine symmetry distinguished the males were pooled (first pair in Fig. 2), preference scores significantly deviated from the null expectation of no discrimination (t 59=3.26, n=60, P=0.002), whereas preference scores did not significantly deviate from the null expectation of no discrimination when the two treatments where only spine length distinguished the males were pooled (last pair in Fig. 2, t 59=0.72, n=60, P>0.4). Preference for symmetry was, though statistically not significant, numerically higher than preference for length in a comparison between the two treatments where only relative spine symmetry differed taken together versus the two treatments where only spine length differed taken together (first pair in Fig. 2 versus last pair in Fig. 2, F 1,118=3.04, P=0.08). Against expectation, the "concordant" combination of traits did not elicit a stronger response, nor did the "contradictory" combination of traits elicit a weaker response than tests of single traits [ordered heterogeneity test (Rice and Gaines 1994) based on ANOVA, F 3,176=1.16, r s P c=-0.21, P>0.4], providing indirect support for at most an only minor role of trait length in mate choice.

Mean (±SE) preference index for spine symmetry and spine length (n=30 females per treatment). For each preference test, at the top the a priori high-quality male is outlined (under the assumption that symmetric spines are preferred over asymmetric spines, and long spines are preferred over short spines). Accordingly, at the bottom the a priori low-quality male is outlined. The dotted line corresponds to the null expectation of no preference. A preference index above the line indicates a preference for the a priori high-quality male. Untransformed preference scores are given
When the two treatments where the stimulus male models differed exclusively in relative spine symmetry were pooled, preference for symmetry and female physical condition were significantly positively associated (F 1,55=7.59, P=0.008; Fig. 3). The relationship was statistically significant when both males had long spines (F 1,26=4.99, P=0.03), not so when both males had short spines (F 1,27=2.20, P>0.1). Overall, female physical condition and preference index were not significantly related (F 1,163=0.29, P>0.5). The six treatments were not biased with respect to female physical condition (one-way ANOVA, F 5,159=0.40, P>0.8).

Relationship between the preference index for symmetry and female physical condition. The dotted line corresponds to the null expectation of no preference. Three females that ate the spawned eggs were excluded from the analysis. Untransformed preference scores are given
Discussion
Females faced with the choice between two male models differing in the relative symmetry of their pelvic spines spent more time with the symmetric male in all of four choice tests involving a difference in relative spine symmetry, regardless of spine length. However, the difference was statistically significant at the α=0.05 level only in the comparison opposing a male model with symmetric short spines to a male model with asymmetric short spines.
The relative importance of size and symmetry in sexual selection is currently a controversial issue. The size of morphological traits was shown to be more sensitive to stress than trait FA in several recent studies (David et al. 1998; Bjorksten et al. 2000a, 2000b), leading to the hypothesis that size is a better predictor of individual quality than FA. In contrast, a meta-analysis of studies of sexual selection in relation to asymmetry (Thornhill and Møller 1998) revealed effect sizes of similar magnitudes for size and symmetry, suggesting that both contribute to success in sexual competition in a complementary fashion. We found no evidence for a role of spine length in mate choice. Females did not significantly discriminate between male models differing exclusively in spine length. Moreover, the preference for symmetry did not significantly vary depending on whether symmetry was the only discriminatory criterion available or whether symmetry and length were combined. The lack of preference for spine length is nevertheless puzzling, since just directional selection (arising from female choice or male-male competition) for larger size is supposed to relax control over development, thus increasing FA in sexually selected traits (Møller and Pomiankowski 1993a). However, in structures with a mechanical function to be performed effectively, the role of natural selection should not be neglected. Pelvic spines are part of an armour complex protecting against vertebrate predators (Hoogland et al. 1957; Bell and Foster 1994), such as piscivorous fish that likely constitute the major threat to sticklebacks of the studied population. Hence, natural (rather than sexual) selection may be the main force determining the size of pelvic spines. Selection for the ability to detect, store and respond to small length differences in bilateral structures indirectly suggests that females can secure advantages by mating with a symmetric partner, and supports a role for FA as an indicator of male quality. However, we cannot rule out that spine length and spine symmetry reflect different aspects of male quality, whose significance depends on the context or varies among females.
Female preference for male spine symmetry was stronger the higher female physical condition (Fig. 3), suggesting that either females with high condition can afford to be more choosy (even in a choice situation without apparent costs), or that they have more to gain by choosing a symmetric male. Alternatively, physiological constraints may impede the perceptual abilities of females with low condition.
We can only speculate about the relative importance of spine symmetry as a choice criterion with respect to other more conspicuous male traits that have been established as targets of sexual selection in sticklebacks, e.g. nuptial coloration (Milinski and Bakker 1990). Several hypotheses have been proposed to explain the evolution of multiple male ornaments (Møller and Pomiankowski 1993b), but little empirical work has addressed the question of how females weigh the information from multiple signals. One possibility is that suites of characters are organised in a hierarchical fashion, with "lower-order" cues being used when "higher-order" cues show low variation or are difficult to discriminate (Jennions and Petrie 1997). Given that in our test situation the two male stimuli were identical in all respects but relative spine symmetry and/or spine length, the absence of variation in more obvious cues may have forced females to shift to a rather inconspicuous criterion as a basis for choice.
The external validity of behavioural experiments based on video playback techniques has recently been challenged (Oliveira et al. 2000 and references therein). One issue raised is pseudoreplication, i.e. a discrepancy between the number of measurements made and the number of independent replicates available for statistical analyses (Hurlbert 1984), resulting in an inflated sample size for the hypothesis being tested (McGregor et al. 1992). The main argument thereby is that the general question of whether a particular aspect of a stimulus is preferred to its alternative cannot be answered through the repeated presentation of only one stimulus, but instead requires either the use of several stimuli (ideally as many as there are replicates) covering a wide range of phenotypic variation, or the de novo synthesis of an "average" stimulus from population data (Rosenthal 1999; McGregor 2000). However, by presenting females with identical copies of a virtual stimulus where only the trait under consideration (spine morphology) underwent artificial modifications, we confidently assume that the difference in the elicited response arises because of a difference in the manipulated trait, rather than because of some underlying aspect peculiar to the real stickleback male that was originally chosen as a template for the virtual model. Another serious concern relates to the potential mismatch between an animal's perception of the stimuli displayed on a screen and the natural counterparts those stimuli attempt to mimic. In this respect, sticklebacks are an adequate study species (Künzler and Bakker 1998), as their visual system closely matches that of humans (C.L. Baube, personal communication), for which video equipment is designed. In spite of potential technical constraints, our data show that computer animations can be a powerful tool in the study of female preferences, particularly if faithful colour rendition is not a major issue and instead features like shape or size are manipulated (Oliveira et al. 2000). We thus further promote the use of computer animations, especially as a valid alternative when proper experimental manipulation of natural ornaments is critical. In the context of the ongoing debate about the significance of FA in sexual selection, computer animations might contribute to shed light on the crucial issue of perceptual thresholds (Swaddle 1999a, 1999b). The accuracy to which asymmetry can be discriminated is decisive for the effectiveness of the signal (Swaddle 1999a, 1999b). In our study, their visual acuity enabled female sticklebacks to discern between perfect symmetry and extreme (albeit realistic) asymmetry. For symmetry to be effective as a signal, females must further be able to discriminate among individuals displaying asymmetry to varying extent (Swaddle 1999a, 1999b). Computer animations allow to stage preference tests with male stimuli exhibiting gradually decreasing levels of asymmetry, and may thus reveal particularly appropriate for understanding whether FA has enough discriminatory power to play a significant role in biological signalling.
References
Bell MA, Foster SA (1994) Introduction to the evolutionary biology of the threespine stickleback. In: Bell MA, Foster SA (eds) The evolutionary biology of the threespine stickleback. Oxford University Press, Oxford, pp 1–27
Bennett ATD, Cuthill IC, Partridge JC, Maier EJ (1996) Ultraviolet vision and mate choice in zebra finches. Nature 380:433–435
Bjorksten T, David P, Pomiankowski A, Fowler K (2000a) Fluctuating asymmetry of sexual and nonsexual traits in stalk-eyed flies: a poor indicator of developmental stress and genetic quality. J Evol Biol 13:89–97
Bjorksten TA, Fowler K, Pomiankowski A (2000b) What does sexual trait FA tell us about stress? Trends Ecol Evol 15:163–166
Bolger T, Connolly PL (1989) The selection of suitable indices for the measurement and analysis of fish condition. J Fish Biol 34:171–182
David P, Hingle A, Greig D, Rutherford A, Pomiankowski A, Fowler K (1998) Male sexual ornament size but not asymmetry reflects condition in stalk-eyed flies. Proc R Soc Lond B 265:2211–2216
Hasson O (1989) Amplifiers and the handicap principle in sexual selection: a different emphasis. Proc R Soc Lond B 235:383–406
Hasson O (1990) The role of amplifiers in sexual selection: an integration of the amplifying and the Fisherian mechanisms. Evol Ecol 4:277–289
Hoogland R, Morris D, Tinbergen N (1957) The spines of sticklebacks (Gasterosteus and Pygosteus) as means of defence against predators (Perca and Esox). Behaviour 10:205–236
Hunt J, Simmons LW (1997) Patterns of fluctuating asymmetry in beetle horns: an experimental examination of the honest signalling hypothesis. Behav Ecol Sociobiol 41:109–114
Hunt J, Simmons LW (1998) Patterns of fluctuating asymmetry in beetle horns: no evidence for reliable signaling. Behav Ecol 9:465–470
Hurlbert SH (1984) Pseudoreplication and the design of ecological field experiments. Ecol Monogr 54:187–211
Jennions MD, Petrie M (1997) Variation in mate choice and mating preferences: a review of causes and consequences. Biol Rev Camb Philos Soc 72:283–327
Künzler R, Bakker TCM (1998) Computer animations as a tool in the study of mating preferences. Behaviour 135:1137–1159
Künzler R, Bakker TCM (2001) Female preferences for single and combined traits in computer animated stickleback males. Behav Ecol 12:681–685
Ludwig W (1932) Das Rechts-Links Problem im Tierreich und beim Menschen. Springer, Berlin Heidelberg New York
Manning JT, Hartley MA (1991) Symmetry and ornamentation are correlated in the peacock's train. Anim Behav 42:1020–1021
McGregor PK (2000) Playback experiments: design and analysis. Acta Ethol 3:3–8
McGregor PK, Catchpole CK, Dabelsteen T, Falls JB, Fusani L, Gerhardt HC, Gilbert F, Horn AG, Klump GM, Kroodsma DE, Lambrechts MM, McComb KE, Nelson DA, Pepperberg IM, Ratcliffe L, Searcy WA, Weary DM (1992) Design of playback experiments: the Thornbridge Hall NATO ARW consensus. In: McGregor PK (ed) Playback and studies of animal communication. Plenum Press, New York, pp 1–9
Milinski M, Bakker TCM (1990) Female sticklebacks use male coloration in mate choice and hence avoid parasitized males. Nature 344:330–333
Møller AP (1990) Fluctuating asymmetry in male sexual ornaments may reliably reveal male quality. Anim Behav 40:1185–1187
Møller AP (1992) Female swallow preference for symmetrical male sexual ornaments. Nature 357:238–240
Møller AP, Pomiankowski A (1993a) Fluctuating asymmetry and sexual selection. Genetica 89:267–279
Møller AP, Pomiankowski A (1993b) Why have birds got multiple sexual ornaments? Behav Ecol Sociobiol 32:167–176
Møller AP, Thornhill R (1998) Bilateral symmetry and sexual selection: a meta-analysis. Am Nat 151:174–192
Morris MR, Casey K (1998) Female swordtail fish prefer symmetrical sexual signal. Anim Behav 55:33–39
Oliveira RF, Rosenthal GG, Schlupp I, McGregor PK, Cuthill IC, Endler JA, Fleishman LJ, Zeil J, Barata E, Burford F, Gonçalves D, Haley M, Jakobsson S, Jennions MD, Körner KE, Lindström L, Peake T, Pilastro A, Pope DS, Roberts SGB, Rowe C, Smith J, Waas JR (2000) Considerations on the use of video playbacks as visual stimuli: the Lisbon workshop consensus. Acta Ethol 3:61–65
Palmer AR, Strobeck C (1986) Fluctuating asymmetry: measurement, analysis, patterns. Annu Rev Ecol Syst 17:391–421
Pelkwijk JJ ter, Tinbergen N (1937) Eine reizbiologische Analyse einiger Verhaltensweisen von Gasterosteus aculeatus. Z Tierpsychol 1:193–200
Rice WR, Gaines SD (1994) Extending nondirectional heterogeneity tests to evaluate simply ordered alternative hypotheses. Proc Natl Acad Sci USA 91:225–226
Rosenthal GG (1999) Using video playback to study sexual communication. Environ Biol Fish 56:307–316
Rosenthal GG (2000) Design consideration and techniques for constructing video stimuli. Acta Ethol 3:49–54
SAS Institute (2000) JMP, Version 4. SAS Institute, Cary, N.C.
Swaddle JP (1999a) Limits to length asymmetry detection in starlings: implications for biological signalling. Proc R Soc Lond B 266:1299–1303
Swaddle JP (1999b) Visual signalling by asymmetry: a review of perceptual processes. Philos Trans R Soc Lond B 354:1383–1393
Swaddle JP, Cuthill IC (1994) Preference for symmetric males by female zebra finches. Nature 367:165–166
Symons PEK (1965) Analysis of spine-raising in the male three-spined sticklebacks. Behaviour 26:1–74
Thornhill R, Møller AP (1998) The relative importance of size and asymmetry in sexual selection. Behav Ecol 9:546–551
Van Valen L (1962) A study of fluctuating asymmetry. Evolution 16:125–142
Watson PJ, Thornhill R (1994) Fluctuating asymmetry and sexual selection. Trends Ecol Evol 9:21–25
Acknowledgements
We thank Carlo Largiadèr, Marc Zbinden and the referees for constructive comments on the manuscript, Heinz Richner for providing facilities, and the Swiss National Science Foundation for financial support. This study conforms to the legal requirements of Switzerland.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. Krause
Rights and permissions
About this article
Cite this article
Mazzi, D., Künzler, R. & Bakker, T.C.M. Female preference for symmetry in computer-animated three-spined sticklebacks, Gasterosteus aculeatus . Behav Ecol Sociobiol 54, 156–161 (2003). https://doi.org/10.1007/s00265-003-0609-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-003-0609-0