
Abstract
A new type II collagenopathy, caused by the p.Gly1170Ser mutation of COL2A1, which presents as premature hip osteoarthritis (OA), avascular necrosis of the femoral head (ANFH) or Legg-Calvé-Perthes (LCP) disease, was recently found in several families with an inherited disease of the hip joint. In this study, femoral head cartilage was harvested for histological and ultrastructural examination to determine the pre-existing generalised abnormalities of the mutant cartilage. The histological results showed that the hierarchical structure of the mutant cartilage and the embedded chondrocytes were markedly abnormal. The expression and distribution of type II collagen was non-uniform in sections of the mutant cartilage. Ultrastructural examination showed obvious abnormal chondrocytes and disarrangement of collagen fibres in the mutant cartilage. Furthermore, the predicted stability of type II collagen dramatically decreased with the substitution of serine for glycine. Our study demonstrated that the p.Gly1170Ser mutation of COL2A1 caused significant structural alterations in articular cartilage, which are responsible for the new type II collagenopathy.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Type II collagen, a major structural component of hyaline cartilage and vitreous humour, consists of three identical polypeptide alpha 1(II) chains [1, 2]. Mutations reported in the gene encoding collagen II (COL2A1) [3], including single base substitution, partial deletions, and splice-site mutations, resulted in a series of type II collagenopathies manifested as inheritable skeletal disorders, such as spondyloepiphyseal dysplasia congenita, Kniest dysplasia, Stickler dysplasia and spondyloepiphyseal dysplasia with premature onset arthrosis. The most common genotype of type II collagenopathies are missense mutations [4], where bulky amino acids are substituted for glycine residues and present as dominant negative effects through the assembly of abnormal alpha 1(II) chains into trimeric collagen molecules, usually causing spondyloepiphyseal dysplasia congenita, which often affects the whole skeletal system.
Recently, a novel mutation of COL2A1 was found to be correlated with diseases of the hip joint [5–8], including avascular necrosis of the femoral head (ANFH), Legg-Calvé-Perthes (LCP) disease, and precocious hip osteoarthritis (OA). The first mutation to be correlated with ANFH was a G-to-A transition in the 50th exon of COL2A1 [6, 8], causing a glycine-to-serine change at codon 1170 (p.G1170S), in two Taiwanese families with ANFH. The affected patients were all adults. Subsequently, the first report of a mutation in hereditary LCP disease was found in a Japanese family with the same missense mutation (p.G1170S) [5]. Recently, we reported age at onset-dependent presentations of premature hip OA, ANFH, or LCP disease in a family from the Chinese mainland carrying the same p.G1170S mutation as the Japanese and Taiwanese families [7]. This is the fourth family whose pathologies are reported to be caused by the same mutation. The phenotypic variations between individuals cannot be explained by environmental factors, because family members had been living in separate provinces with different environmental exposures. Therefore, we speculated that the p.G1170S mutation of COL2A1 might lead to a new type II collagenopathy presented as ANFH/precocious hip OA/LCP disease [7].
Although missense mutations of COL2A1 have been extensively reported, the phenotypes were mostly a combination of skeletal dysplasia and vitreous degeneration [4, 9]. The clinical manifestations of affected patients often included short status, spondyloepiphyseal dysplasia, high myopia, or even death, either just before or shortly after birth [3, 9]. Histological and ultrastructural study of some patients with lethal mutations demonstrated severe skeletal abnormalities, including disorganised growth plate and cartilage, decreased biosynthesis of type II collagen, as well as dysfunction chondrocytes with dilated rough endoplasmic reticulum inclusion [10–13]. The p.G1170S mutation, in contrast with the most missense mutations, only leads to milder phenotypes confined to the hip joint, without involvement of spinal or ocular abnormalities [5–7]. Furthermore, the phenotypes of p.G1170S were ANFH/hip OA/LCP disease, which were thought to be diseases with multiple aetiologies [14, 15] and requiring various treatment strategies [16–18]. Hence, studying the histological and ultrastructural characteristics of mutant cartilage with the p.G1170S mutation will help to understand the pathogenesis of this new type II collagenopathy, as well as ANFH/hip OA/LCP disease.
In this study, femoral head cartilage was harvested from patients for histological and ultrastructural examination to determine the preexisting generalised abnormalities of the p.G1170S mutant cartilage. Furthermore, prediction of the stability of the mutant collagen triple-helix was performed using the online collagen stability calculator.
Materials and methods
Patients and mutant femoral head cartilage
We found a five-generation family with 42 members with a new type II collagenopathy caused by the p.G1170S mutation of COL2A1 [7], including 16 affected members (five with isolated precocious hip OA, six with ANFH, and five with LCP disease). The availability of mutant cartilage in this family was limited. It was impossible to biopsy cartilage specimens from patients. Fortunately, a 56-year-old woman (family member III-2, age at onset 15 years, diagnosed as ANFH ) [7] and a 65-year-old man (family member II-6, age at onset 21 years, diagnosed as OA of the hip joint) [7] underwent total hip replacement in our hospital, and the two femoral heads were collected for histological evaluation and transmission electron microscopy examination. As age controls, three femoral cartilage specimens were obtained from age- and gender-matched patients with ANFH during surgery. All tissue samples used in this study was approved by the ethics committee of Sun Yat-Sen University and conducted in compliance with the Declaration of Helsinki. Written informed consent was obtained from all subjects.
Fixation, section and toluidine blue staining
Both mutant and control cartilage specimens were taken from the same position in the margin of the femoral head, because the patients who had undergone total hip replacement were suffering from severe osteoarthritis, according to the radiographic appearance [19], as a result of which, the cartilage in the load-bearing surface was almost completely destroyed due to wear and tear. The cartilage specimens were fixed in 10% neutral-buffered formalin overnight, dehydrated in an alcohol gradient, and embedded in low melting point paraffin. Continuous 5-µm thick tissue sections were cut and fixed onto silicified slides. The sections were deparaffinised and rehydrated with deionised Millipore water. Sections were stained with 1% toluidine blue with borate for 20 min, rinsed with tap water, dehydrated, immersed in xylene for 10 min, and then mounted with neutral resins.
Immunohistochemistry analysis
Immunohistochemistry was performed using the streptavidin–peroxidase-conjugated method. Briefly, cartilaginous tissue sections were deparaffinised, rehydrated, and incubated with fresh 3% hydrogen peroxide in methanol for 10 min at room temperature. After rinsing with phosphate-buffered saline (PBS), antigen retrieval was carried out by microwave treatment in 0.01 M sodium citrate buffer (pH 6.0) at 100°C for 15 min. Non-specific binding was blocked with normal goat serum for 15 min at room temperature, followed by incubation with monoclonal mouse anti-human collagen II antibody (Santa Cruz Biotechnology, Santa Cruz, USA) diluted in PBS containing 0.2% Triton X-100 and appropriate 2% normal serum (Serva, Heidelberg, Germany) overnight at 4°C. After rinsing with PBS, slides were incubated for 10 min at room temperature with biotin-conjugated secondary antibodies, followed by incubation with a streptavidin-conjugated peroxidase working solution for 10 min. Subsequently, sections were stained for 10 min with 3,3′-diaminobenzidine tetrahydrochloride (DAB), counterstained with Mayer’s haematoxylin, dehydrated, and mounted. Negative controls were prepared by substituting PBS for the primary antibody.
Transmission electron microscopy (TEM)
The cartilage specimens used for TEM examination were taken from the same position in the outer layer of femoral head cartilage. According to standard protocols [20, 21], the cartilage specimens of controls and patients were chemically fixed in an aqueous solution of glutaraldehyde (2.5%) in 0.1 M sodium cacodylate buffer (pH 7.4) for two days at room temperature. Decalcification was achieved by treatment of the samples with 10% EDTA, 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) for five days. The samples were then post-fixed with a 1% osmium tetroxide solution (sodium cacodylate buffer, pH 7.4) over a period of four hours at room temperature and then over 20 hours at 4°C. The specimens were then washed four times in isotonic sodium cacodylate buffer (0.1 M; pH 7.4) and dehydrated through a graded series of ethanol. Thereafter, the specimens were embedded in epoxy resin. Polymerisation was performed at 60°C over one week. Thin sections were cut at a thickness of about 60 nm and stained with uranyl acetate and lead citrate. After staining, the sections were examined by transmission electron microscopy (Phillips CM-10, Eindhoven, Netherlands).
Prediction of the stability of collagen fibril from amino acid sequence
The stability of the collagen triple-helix was predicted from the amino acid sequence using an algorithm based on experimental host–guest peptide thermal stability data [22] (http://jupiter.umdnj.edu/collagen_calculator/index.php). Protein sequence information of type II collagen alpha 1 isoform 1 precursor (NP_001835) was acquired from the NCBI protein database. Part of the sequence of type II collagen alpha 1 isoform 1 precursor (amino acid 903 to 1214) was selected and all prolines in the Y position were converted to hydroxyproline according to the instructions [22]. The relative stability was then generated by the collagen stability calculator.
Results
Pathological appearance of the mutant cartilage
The density and morphological appearance of the chondrocytes embedded in cartilage varies with the depth of the tissue, according to which normal articular cartilage can be divided into three zones: superficial zone (10 ∼ 20%), middle zone (40 ∼ 60%), and deep zone (30%) [23]. There were obvious differences in the toluidine blue-stained sections of femoral head cartilage between the family patients and the control patients. The proportion of deep zone cartilage, where chondrocytes are aligned into columns perpendicular to the tidemark, extended dramatically, to at least 70% in the mutant femoral head cartilage, as a result of which the middle zone shrunk to 20%. The cartilage from control patients showed normal structural hierarchy. In addition, the chondrocytes embedded in the mutant cartilage were morphologically abnormal and showed a disturbed arrangement (Fig. 1a, b). The distribution and appearance of chondrocytes were normal in cartilage from control patients (Fig. 1c, d). The subchondral bone under the deep zone of the cartilage, however, appeared normal in both mutant and control specimens, though the bone in the core of the femoral head had undergone necrosis.

Histological analysis of the cartilage. a, b Toluidine blue staining of cartilage from patients in a large Chinese family with inherited diseases of the hip joint. c, d Toluidine blue staining of cartilage from patients with sporadic avascular necrosis of the femoral head (ANFH). (a and c, magnification ×40. b and d, magnification ×100. Scale bar = 100 μm)
The expression of type II collagen in mutant cartilages
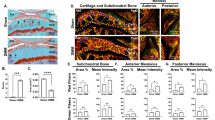
The expression of type II collagen was relatively homogeneous and the collagen fibrils were distributed in both intercellular and pericellular districts of control cartilages (Fig. 2c, d). However, the collagen fibrils in the mutant cartilage were particularly concentrated in the pericellular-territorial matrix compartment. Furthermore, the collagen fibrils were apparently disarranged and gathered in clusters in most of the mutant cartilage sections. The presence of clustered collagen fibres, which were vertical to the tide line, confirmed the result that the proportion of the deep zone had increased in the mutant cartilage (Fig. 2a, b). In addition, there was no apparent accumulation of type II collagen in the intracellular region of both mutant and control chondrocytes. There was no difference in staining between the mutant and control cartilage for type I collagen (data not shown).

Immunohistochemical analysis of collagen type II in the cartilage matrix. a, b Maldistribution collagen fibres in the cartilage from patients in the family with inherited diseases of the hip joint. b Uniform distribution collagen fibres in the cartilage from patients with sporadic avascular necrosis of the femoral head (ANFH). (a and c, magnification ×100. b and d, magnification ×400. Scale bar = 100 μm)
Ultrastructural characterisation of the mutant cartilage
Ultrastructural study of mutant cartilages by transmission electron microscopy (TEM) revealed distinctive differences compared to control cartilages. In the intercellular matrix of the mutant cartilage, the diameter and distribution of collagen fibres were non-uniform, and they were disarranged (Fig. 3a, c), indicating that the biomechanical properties of the mutant cartilage might be significantly weakened. The control cartilage, in contrast, demonstrated a regular arrangement of collagen fibres (Fig. 3b, d). Furthermore, the mutant chondrocytes displayed a bigger nucleus and a few dilated rough endoplasmic reticula, while the nucleus and the rough endoplasmic reticulum (RER) in the control chondrocytes were normal (Fig. 3a, b).

Electron micrographs of chondrocyte and cartilage matrices from the femoral head cartilage of the familial patients (a, c) and sporadic cases of avascular necrosis of the femoral head (ANFH) patients (b, d). a The mutant chondrocyte increased in size, karyoplasmic ratio, and shows dilated rough endoplasmic reticulum (black arrow). b Wild-type chondrocyte. c Disarrangement of collagen fibers in the mutant cartilage. d Arrangement of collagen fibres in a normal cartilage matrix. (a and b, magnification ×8900, scale bar = 1 μm. c and d, magnification ×21000, scale bar = 100 nm)
Prediction of the stability of the type II collagen fibril
Evaluation of the local thermal stability revealed that the p.G1170S mutation significantly undermined the thermal stability of the type II collagen fibril, which decreased from 35°C (wild-type) to 28°C (p.G1170S mutation) (Fig. 4).

Relative stability profiles for wild-type and p.Gly1170Ser mutation type II collagen (amino acids 903 to 1214 are shown) obtained from the collagen stability calculator. The arrows (p.1170th amino acid) denote the low thermal stability of type II collagen fibril
Discussion
Type II collagenopathies, caused by mutation of the COL2A1 gene, include achondrogenesis II, hypochondrogenesis, spondyloepiphyseal dysplasia congenita, Kniest dysplasia, as well as Stickler dysplasia [3, 4, 9]. Most of these are severe congenital diseases that involve the whole skeletal system.
In this study, mutant femoral head cartilage from a milder type II collagenopathy, manifested as hip joint disorders, were subjected to histological and ultrastructural investigation to reveal pre-existing abnormalities of mutant cartilage. Our data demonstrated obvious alterations in the expression and distribution of type II collagen, in the arrangement of collagen fibers, as well as in the hierarchical structure of the mutant cartilage. In addition, the predicted thermal stability of the mutant collagen amino acid sequence decreased dramatically. As a result, the biomechanical properties of mutant cartilage might be reduced, though experiments to confirm this were not performed in this study. Hence, we postulate that the abnormal cartilage caused by the p.G1170S mutation of the COL2A1 gene resulted in this new type II collagenopathy.
Although this new collagenopathy presented as premature hip OA, ANFH, or LCP disease, our data implied that the pathology was not in fact OA, ANFH, or LCP disease, but some form of similar lesion of the femoral head cartilage. The histological and ultrastructural characteristics were normal in the control cartilages from sporadic ANFH patients, while the mutant cartilage displayed obvious abnormal hierarchical structures and arrangements of collagen fibrils. OA, ANFH, and LCP disease were thought to be diseases with multiple aetiologies, including vascular occlusion or ischemia, altered fat metabolism, and fat emboli [14]. We speculate that the p.G1170S mutation of COL2A1 gene undermines the properties of the mutant cartilage, increasing susceptibility to hip joint disorders. Hence, if the carriers found in these families with hereditary hip joint disorders [5–7] were asked to pay more attention to protecting their cartilage, they might avoid hip joint disorders, or might be affected later in life.
Most collagenopathies caused by mutations of the COL2A1 gene are congenital, symmetrical, and progressive [3, 4]. Some severe collagenopathies are lethal or significantly affect the development of skeletal system, which present as confused cartilage structures on the growth plate with numerous vascular invasions [10, 12, 13]. Furthermore, mutant chondrocytes from severe phenotype collagenopathies were found to have problems processing type II collagen synthesis, as well as with post-translational modification, secretion, and assembly into collagen fibers [12]. However, in contrast to the severe phenotypes, the p.G1170S mutation mainly produced milder clinical manifestations. Histological and ultrastructural examination revealed a relative mild alteration in the hierarchical structure. Although disarrangement of collagen fibers was found in the mutant cartilage by TEM, the amount of collagen fibre was not decreased compared to the control cartilage. In contrast with the numerous dilated RER observed in chondrocytes from the severe phenotypes [13], there were only a few dilated RERs in chondrocytes with p.G1170S mutation, indicating that the p.G1170S mutation has a milder effect on the synthesis of type II collagen. Although the thermal stability of the collagen fibril was undermined (Fig. 4), the p.G1170S mutation of COL2A1, to the best of our knowledge, only has a moderate effect on cartilage.
As type II collagen is mainly found in hyaline cartilage and vitreous humour, its deficiency might be associated with spinal, articular and ocular abnormalities [9]. However, the p.G1170S mutation of the COL2A1 gene mainly produced abnormalities confined to the hip joint. Except for the abnormalities in the mutant cartilage, several other factors might be attributed to the onset of hip joint diseases, including the joint’s weight-bearing function, specific local biomechanical properties, poor vascular supply, and pre-existing dysplasia of femoral head. However, the mechanism by which the p.G1170S mutation of COL2A1 causes the hip joint diseases remains unclear.
In conclusion, this study demonstrated the histological and ultrastructural characteristics of cartilage from a new type II collagenopathy caused by the p.G1170S mutation of the COL2A1 gene. Although the precise molecular mechanism underlying the morphological alterations requires further research, carriers of the mutation could avoid or delay the emergence of hip joint disorders by taking some action to protect their hip joints.
References
Prockop DJ, Kivirikko KI (1995) Collagens: molecular biology, diseases, and potentials for therapy. Annu Rev Biochem 64:403–434
Ramirez F, Boast S, D’Alessio M, Lee B, Prince J, Su MW, Vissing H, Yoshioka H (1990) Fibrillar collagen genes. Structure and expression in normal and diseased states. Ann N Y Acad Sci 580:74–80
Kuivaniemi H, Tromp G, Prockop DJ (1997) Mutations in fibrillar collagens (types I, II, III, and XI), fibril-associated collagen (type IX), and network-forming collagen (type X) cause a spectrum of diseases of bone, cartilage, and blood vessels. Hum Mutat 9:300–315
Nishimura G, Haga N, Kitoh H, Tanaka Y, Sonoda T, Kitamura M, Shirahama S, Itoh T, Nakashima E, Ohashi H, Ikegawa S (2005) The phenotypic spectrum of COL2A1 mutations. Hum Mutat 26:36–43
Miyamoto Y, Matsuda T, Kitoh H, Haga N, Ohashi H, Nishimura G, Ikegawa S (2007) A recurrent mutation in type II collagen gene causes Legg-Calve-Perthes disease in a Japanese family. Hum Genet 121:625–629
Liu YF, Chen WM, Lin YF, Yang RC, Lin MW, Li LH, Chang YH, Jou YS, Lin PY, Su JS, Huang SF, Hsiao KJ, Fann CS, Hwang HW, Chen YT, Tsai SF (2005) Type II collagen gene variants and inherited osteonecrosis of the femoral head. N Engl J Med 352:2294–2301
Su P, Li R, Liu S, Zhou Y, Wang X, Patil N, Mow CS, Mason JC, Huang D, Wang Y (2008) Age at onset-dependent presentations of premature hip osteoarthritis, avascular necrosis of the femoral head, or Legg-Calve-Perthes disease in a single family, consequent upon a p.Gly1170Ser mutation of COL2A1. Arthritis Rheum 58:1701–1706
Chen WM, Liu YF, Lin MW, Chen IC, Lin PY, Lin GL, Jou YS, Lin YT, Fann CS, Wu JY, Hsiao KJ, Tsai SF (2004) Autosomal dominant avascular necrosis of femoral head in two Taiwanese pedigrees and linkage to chromosome 12q13. Am J Hum Genet 75:310–317
Spranger J, Winterpacht A, Zabel B (1994) The type II collagenopathies: a spectrum of chondrodysplasias. Eur J Pediatr 153:56–65
Godfrey M, Keene DR, Blank E, Hori H, Sakai LY, Sherwin LA, Hollister DW (1988) Type II achondrogenesis-hypochondrogenesis: morphologic and immunohistopathologic studies. Am J Hum Genet 43:894–903
Godfrey M, Hollister DW (1988) Type II achondrogenesis-hypochondrogenesis: identification of abnormal type II collagen. Am J Hum Genet 43:904–913
Bonaventure J, Cohen-Solal L, Ritvaniemi P, van Maldergem L, Kadhom N, Delezoide AL, Maroteaux P, Prockop DJ, Ala-Kokko L (1995) Substitution of aspartic acid for glycine at position 310 in type II collagen produces achondrogenesis II, and substitution of serine at position 805 produces hypochondrogenesis: analysis of genotype-phenotype relationships. Biochem J 307(Pt 3):823–830
Korkko J, Cohn DH, Ala-Kokko L, Krakow D, Prockop DJ (2000) Widely distributed mutations in the COL2A1 gene produce achondrogenesis type II/hypochondrogenesis. Am J Med Genet 92:95–100
Assouline-Dayan Y, Chang C, Greenspan A, Shoenfeld Y, Gershwin ME (2002) Pathogenesis and natural history of osteonecrosis. Semin Arthritis Rheum 32:94–124
Mont MA, Jones LC, Hungerford DS (2006) Nontraumatic osteonecrosis of the femoral head: ten years later. J Bone Jt Surg Am 88:1117–1132
Floerkemeier T, Thorey F, Daentzer D, Lerch M, Klages P, Windhagen H, von Lewinski G (2010) Clinical and radiological outcome of the treatment of osteonecrosis of the femoral head using the osteonecrosis intervention implant. Int Orthop. doi:10.1007/s00264-009-0940-9
Vukasinovic Z, Spasovski D, Vucetic C, Cobeljic G, Zivkovic Z, Matanovic D (2009) Triple pelvic osteotomy in the treatment of Legg-Calve-Perthes disease. Int Orthop 33:1377–1383
Kim YH, Choi Y, Kim JS (2009) Cementless total hip arthroplasty with ceramic-on-ceramic bearing in patients younger than 45 years with femoral-head osteonecrosis. Int Orthop. doi:10.1007/s00264-009-0878-y
Kellgren JH, Jeffrey MR, Ball J (1963) The epidemiology of chronic rheumatism. Vol II: Atlas of standard radiographs of arthritis. Blackwell Scientific, Oxford
Hunziker EB, Wagner J, Studer D (1996) Vitrified articular cartilage reveals novel ultra-structural features respecting extracellular matrix architecture. Histochem Cell Biol 106:375–382
Hunziker EB, Michel M, Studer D (1997) Ultrastructure of adult human articular cartilage matrix after cryotechnical processing. Microsc Res Tech 37:271–284
Persikov AV, Ramshaw JA, Brodsky B (2005) Prediction of collagen stability from amino acid sequence. J Biol Chem 280:19343–19349
Mow VC, Gu WY, Chen FH (2005) Structure and function of articular cartilage and meniscus. Lippincott Williams and Wilkins, Philadelphia, pp 181–258
Acknowledgments
We thank the patients and their families for their cooperation. We also thank Professor Yiming Wang (Zhongshan School of Medicine and Center for Genome Research, Sun Yat-Sen University ) for providing technical guidance. This work was supported by grants from the National Natural Science Foundation of China (NO.30971587), the Natural Science Foundation of Guangdong province (NO.8151008901000141) and the Yat-Sen scholarship for young scientists.
Conflict of interest
There were no conflicts of interest in this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
Peiqiang Su and Liangming Zhang contributed equally to this work.
Rights and permissions
About this article
Cite this article
Su, P., Zhang, L., Peng, Y. et al. A histological and ultrastructural study of femoral head cartilage in a new type II collagenopathy. International Orthopaedics (SICOT) 34, 1333–1339 (2010). https://doi.org/10.1007/s00264-010-0985-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00264-010-0985-9