
Abstract
Promising anti-tumor responses have been observed in the clinic using monoclonal antibodies (mAbs) that block immune checkpoints. One concern with these therapeutic agents remains the potential induction of immune breakthrough events (IBEs) resulting from the disruption of T cell homeostasis or the breaking of tolerance to self antigens. As an approach to maintaining anti-tumor responses but decreasing the likelihood of these events, the local expression of a mAb in combination with a GM-CSF-secreting cancer immunotherapy was evaluated. Using anti-cytotoxic T lymphocyte antigen (CTLA)-4 as a model antibody to test this hypothesis, tumor cell lines were generated that expressed the full-length mAb in addition to GM-CSF. Evaluation of these cell lines in two therapeutic tumor models revealed that local, cell-mediated delivery of anti-CTLA-4 from a GM-CSF-secreting tumor cell immunotherapy activated potent anti-tumor responses and prolonged overall survival at significantly lower serum mAb levels in the host. Furthermore, lowering the systemic exposure of the host to the immune modulatory mAb correlated with reduced evidence of systemic autoimmunity. This approach has broad utility for the delivery of mAbs or proteins locally from cellular immunotherapies to minimize IBEs while retaining the potent therapeutic effects of such combination treatments.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Monoclonal antibodies are important therapeutic agents for the treatment of cancer, and constitute the second largest class of drugs after anti-infective vaccines [1]. The majority of mAbs approved for clinical use are administered by intravenous injection. Although systemic administration may offer advantages with respect to targeting multiple disease sites, high doses may be required for clinical responses. This in turn may create challenges in manufacturing, antibody processing and folding, and systemic toxicity. For example, therapeutic mAbs blocking immune checkpoints have yielded promising anti-tumor responses, but intravenous administration of these mAbs has been reported to potentially result in systemic or organ-specific autoimmune events [2]. Thus, improved methods of mAb delivery for such antibodies are desirable.
GM-CSF-secreting cancer immunotherapies have been shown to elicit specific, potent, and durable anti-tumor responses in preclinical and clinical studies [3, 4]. The irradiated tumor cells are known to persist at the immunization site [5], and thus could be used as a platform for the local secretion of proteins. In the studies presented here, this hypothesis was tested by engineering such cells to produce a blocking mAb to CTLA-4. CTLA-4 is an important negative regulator of activated T cells, and disruption of this pathway is one of the most prominent examples of using an immuno-modulatory mAb to enhance anti-tumor responses [2]. The combination of a GM-CSF-secreting tumor cell immunotherapy and CTLA-4 blockade has also previously been shown to yield specific, potent, and durable anti-tumor immune responses in preclinical and clinical studies [2, 6]. Although clinical studies evaluating CTLA-4 blockade have generated promising anti-tumor responses, treatment with anti-CTLA-4 has been shown to elicit IBEs in some patients [2, 6, 7]. As one approach to reducing the frequency and/or severity of these events while retaining the therapeutic benefit, the local secretion of anti-CTLA-4 from a GM-CSF-secreting tumor cell immunotherapy was evaluated in preclinical models.
Materials and methods
Antibody cloning and vectors
Using standard molecular biology techniques, the heavy and light chains of a mouse anti-mouse IgG2b anti-CTLA-4 antibody were cloned from a hybridoma cell line (clone 9D9). Consensus variable region sequences were obtained from the sequencing data derived from multiple independent PCR reactions. The DNA fragments that encode the heavy chain, a furin cleavage site, the 2A sequence, and the light chain were linked together and cloned into a retroviral transfer vector. Artificial oligonucleotides for FMDV 2A sequence were synthesized based on the peptide sequence APVKQTLNFDLLKLAGDVESNPGP. The retroviral transfer vector plasmid (rkat3F3) used for these studies has previously been described [8], and was subsequently modified to contain additional restriction enzyme sites flanking the extracellular domain of the human CD4 chimera (F3). Retroviral vectors were generated by transient transfection as previously described [8, 9].
Cell lines and in vitro assays
The murine CT26 adenocarcinoma and B16F10 melanoma cell lines were purchased from American Type Culture Collection. The B16GM and CT26GM cell lines have previously been described [10]. Anti-CTLA-4-producing CT26GM and B16GM cell lines were generated by transduction using concentrated F2A anti-CTLA-4 retroviral supernatants. The antibody expressing cells were used as a population derived from a single round of infection, and expression of the anti-CTLA-4 protein was confirmed using IgG2b (Bethyl) and anti-CTLA-4 specific ELISAs. The latter ELISA was performed using plates coated with recombinant mouse CTLA-4/Fc chimeric protein (R&D systems).
Mice
BALB/c and C57BL/6 mice were purchased from Taconic and DO11.10 mice were purchased from The Jackson Laboratory. Animal studies were initiated when mice were between 8 and 12 weeks of age. All mice were maintained according to National Institute of Health guidelines and experiments were performed according to protocols reviewed and approved by the Cell Genesys Animal Use and Care Committee.
In vivo proliferation assay
Recombinant antibody was purified from the cell culture supernatants of an anti-CTLA-4 hybridoma, B16GM F2A anti-CTLA4 cells, or a control hybridoma. The latter cross reacts with the synthetic peptide DYKDDDDK (clone 4E11, ATCC) and was used as an IgG2b isotype control. mAbs were purified using protein G columns (Pierce) and confirmed to have low levels of endotoxin levels by ELISA (Cambrex). To perform adoptive transfers, 1 × 106 erythrocyte depleted splenocytes from DO11.01 mice were injected into recipient BALB/c mice. Two days later, mice were treated with 500 μg of ovalbumin protein (Sigma) and 100 μg of purified recombinant antibody by subcutaneous injection. The antibody injections were repeated the following day. Three days later, the spleens and lymph nodes were harvested and OVA-expressing T cells were identified by FACS using the KJ126 monoclonal antibody (Caltag) specific for the T cell receptor of DO11.01 cells.
Tumor model studies
For CT26 studies, tumors were pre-established by subcutaneous injection of 5 × 105 live tumor cells at a dorsal site. Immunizations were performed using 1 × 106 irradiated immunotherapy cells, which were injected at a ventral site on day 3 post-challenge with live tumor cells. Recombinant anti-CTLA-4 was administered the same day. Local anti-CTLA-4 was delivered by premixing 10 μg of recombinant anti-CTLA-4 with the irradiated tumor cells prior to injection. For the systemic groups, 10 or 150 μg of recombinant anti-CTLA-4 was delivered by intraperitoneal injection. For B16F10 tumor studies, C57BL/6 mice were inoculated with 2 × 105 live B16F10 tumor cells. The following day mice were injected with either HBSS, systemic anti-CTLA-4 (D1 = 150 μg, D4 = 100 μg), or 3 × 106 irradiated B16GM cells. In the combination groups, mice were immunized with 3 × 106 irradiated B16GM cells and received either a high dose (D1 = 150 μg, D4 = 100 μg) or low dose (D1 = 15 μg, D4 = 10 μg) of systemic or local (D1 = 15 μg, D4 = 10 μg) recombinant anti-CTLA-4. Systemic administration of the antibody was performed by intraperitoneal injection, and local administration was performed by subcutaneous injection at the immunization site. For the latter group, the irradiated tumor cells and recombinant anti-CTLA-4 were premixed prior to injection. The final group received 3 × 106 irradiated B16GM cells modified to secrete anti-CTLA-4 using a F2A cassette. A second round of immunizations and antibody injections was administered starting on day 14. Tumor burden was monitored twice weekly, and mice were sacrificed when tumors reached 1,500 mm3 or severe ulceration developed. Data are presented as a Kaplan–Meier survival curve with n = 10 per group. Serum was collected by retro-orbital bleeds and circulating anti-CTLA-4 levels were detected by ELISA as previously described.
Autoimmune antibody levels
Mice were injected with HBSS, systemic anti-CTLA-4, 3 × 106 irradiated B16GM tumor cells, or 3 × 106 irradiated B16GM cells modified to secrete anti-CTLA-4. Systemic anti-CTLA-4 was administered by intraperitoneal injection on days 0 (150 μg) and 3 (100 μg). The immunization and antibody treatment regime was repeated bi-weekly. Serum was collected every 2 weeks by retro-orbital bleeds, and antibodies to ANA, ssDNA, and dsDNA were detected by ELISA (Alpha Diagnostics).
Statistical analysis
Multi-parameter statistics for the Kaplan–Meier survival curves were performed by a log-rank test. Where indicated, the relative differences between groups was evaluated by a Student’s t test and reported as a P value with a 95% confidence interval. Analyses were performed using Prism (GraphPad software) and a P value of less than 0.05 was considered to be statistically significant.
Results and discussion
Cloning and functional analysis of anti-CTLA-4 cassette
The nucleotide sequence of a mouse anti-mouse mAb to CTLA-4 was derived from a hybridoma, and a retroviral transfer plasmid was constructed with the heavy (HC) and light (LC) chain coding sequences connected using the foot-and-mouth-disease virus (FMDV)-derived 2A self-processing sequence (Fig. 1a). The 2A peptide sequence is located between the P1 and P2 proteins in some members of the picornavirus family, and efficiently “cleaves” at its own C terminus through an enzyme-independent mechanism (ribosomal skip) during protein translation [11–13]. To generate a fully native antibody, the expression cassette was further engineered to include a furin cleavage site sequence located between the HC and the 2A sequence that cleaves residual amino acids that are derived from the 2A sequence post-cleavage. The final expression cassette provides efficient and equimolar expression of the antibody HC and LC from a single promoter, and yields high levels of full-length, native, functional mAbs in vitro and in vivo [14].

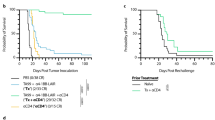
Local secretion of anti-CTLA-4 from a whole cell immunotherapy enhances immunotherapy efficacy. a Diagram of a retroviral transfer vector expressing a full-length murine anti-CTLA-4 monoclonal antibody from a single promoter by linking the antibody heavy and light chains using furin and 2A cleavage sites. LTR, long-terminal repeat, Ψ, packaging signal. b Functional analysis of F2A anti-CTLA-4 in vivo. Splenocytes were collected from ovalbumin-specific T cell receptor (OVA-TCR) transgenic mice (designated DO11.01), and adoptively transferred by intravenous injection into recipient BALB/c mice. Two days later, mice received a subcutaneous injection of 500 μg OVA protein and 100 μg of the following purified mAbs: an IgG2b isotype control (naive); anti-CTLA-4 purified from a hybridoma; anti-CTLA-4 purified from B16GM F2A anti-CTLA4 cells; or HBSS. The antibody injections were repeated the following day. Three days after the last injection, splenocytes were collected and evaluated for the presence of OVA-TCR-specific T cells by FACs analysis using antibodies specific for CD4 and the OVA-TCR (designated KJ126). The absolute number of KJ126+/CD4+ T cells is shown
A retroviral vector was generated using the anti-CTLA-4 F2A expression cassette and used to infect CT26 and B16F10 cells that were previously modified to secrete GM-CSF (designated CT26GM and B16GM, respectively). The resulting CT26GM F2A anti-CTLA-4 and B16GM F2A anti-CTLA-4 populations express 11 and 3 pg/cell/day, respectively, of anti-CTLA-4 in vitro (data not shown). The parental CT26GM and B16GM cell lines secrete approximately 50 and 200 ng/1 × 106 cells/day, respectively, of murine GM-CSF in vitro and no changes in the level of GM-CSF secretion were detected in the mAb-producing cell lines. To confirm the function of the mAb secreted from the cell lines, recombinant anti-CTLA-4 was purified from B16GM F2A anti-CTLA-4 cell lines and compared to recombinant antibody purified from the corresponding hybridoma using an in vivo T cell proliferation assay (Fig. 1b). Splenocytes were collected from DO11.01 mice, which contain T-cells transgenic for an ovalbumin (OVA)-specific T cell receptor (OVA-TCR) at high frequency, and adoptively transferred by intravenous injection into recipient BALB/c mice. Mice were treated with a subcutaneous injection of OVA protein and either 100 μg of an IgG2b isotype control, recombinant anti-CTLA-4 purified from a hybridoma, or recombinant antibody expressed from B16GM F2A anti-CTLA4 cells, or HBSS. Three days later, splenocytes were collected from mice and evaluated for OVA-TCR transgenic T cells by FACs analysis. The anti-CTLA-4 mAb expressed from either the B16GM immunotherapy cells or the parental hybridoma augmented T cell expansion to approximately the same levels, confirming that the recombinant anti-CTLA-4 mAb produced from the B16GM cells is functionally active.
Local secretion of anti-CTLA-4 enhanced anti-tumor responses with lower systemic mAb exposure
The administration of anti-CTLA-4 as a monotherapy enhances immune responses and promotes tumor regression in multiple types of immunogenic tumors [2, 6]. In this study the synergy between a GM-CSF-secreting immunotherapy and anti-CTLA-4 was evaluated using the immunogenic, murine CT26 colon adenocarcinoma model. The dose and timing of the GM-CSF-secreting immunotherapy was selected to result in moderate anti-tumor efficacy as a monotherapy [10], providing a window to observe enhanced therapeutic benefit upon anti-CTLA-4 administration and/or mAb co-secretion. BALB/c mice were inoculated with a lethal dose of live CT26 tumor cells, and 3 days later immunized with CT26GM cells. By day 35, 20% of the mice immunized with GM-CSF-secreting immunotherapy cells, and 90% of the mice treated with the combination of CT26GM and a high dose (150 μg) of systemic anti-CTLA-4, rejected the tumor (Fig. 2a). To evaluate the local secretion of anti-CTLA-4 from the immunotherapy cells, mice were immunized at the same dose and timing with CT26GM cells genetically modified to secrete anti-CTLA-4 (designated CT26GM F2A anti-CTLA-4). All of the mice in this group rejected the tumor challenge, demonstrating an enhanced anti-tumor response with antibody-secreting tumor cells when compared to mice immunized with CT26GM alone. Serum was collected from mice at the indicated days and evaluated for anti-CTLA-4 levels by ELISA (Fig. 2b). The half life of anti-CTLA4 in serum was ∼4 days in the systemic group, whereas in the cell-secreting group the levels peaked at day 3 and gradually declined over a period of 2 weeks. Thus, although the serum antibody levels in mice treated with systemic anti-CTLA-4 were 5–20-fold higher than those in mice treated with the cell-secreting immunotherapy, there was no significant difference in the survival benefit observed between these groups (P = 0.3173).

Local secretion of anti-CTLA-4 from a GM-CSF-secreting tumor cell immunotherapy enhances anti-tumor responses. a BALB/c mice were inoculated subcutaneously with 5 × 105 live CT26 tumor cells. Three days later mice were immunized with 1 × 106 irradiated CT26GM cells in combination with either systemic (10 or 150 μg) or local (10 μg) recombinant anti-CTLA-4. For the latter group, the irradiated tumor cells and recombinant anti-CTLA-4 were premixed prior to injection. The final group was immunized with CT26GM cells modified to secrete anti-CTLA-4. Local secretion of anti-CTLA-4 from CT26GM cells (CT26GM F2A anti-CTLA-4) significantly enhanced anti-tumor responses when compared to the CT26GM (*P = 0.0003), CT26 systemic anti-CTLA-4 Low dose (*P = 0.0039), and CT26GM local anti-CTLA-4 Low dose (*P = 0.0115) groups. b Kinetic expression profile of cell secreting and systemic anti-CTLA-4. Retro-orbital bleeds were performed at the indicated time points from all mice treated in panel a. Serum anti-CTLA-4 levels were evaluated by ELISA. c Expression levels of anti-CTLA-4. Retro-orbital bleeds were performed 3 days post-immunization/antibody injection from satellite mice treated as described in Fig. 2a. Serum anti-CTLA-4 levels were evaluated by ELISA. Compared to mice treated with the 150 μg systemic antibody dose (CT26GM systemic anti-CTLA-4), significantly lower levels of serum anti-CTLA-4 were detected in all other groups (*P < 0.0001)
To further evaluate whether co-injection of recombinant anti-CTLA-4 at the immunization site with the GM-CSF secreting tumor cell immunotherapy could yield comparable efficacy to antibody-secreting immunotherapy cells, CT26 tumor-bearing BALB/c mice were immunized with CT26GM cells in combination with either systemic (10 or 150 μg doses) or local (10 μg) anti-CTLA-4. Although low doses of systemic or local anti-CTLA-4 in combination with CT26GM yielded increased survival compared to immunization with CT26GM alone, the anti-tumor responses were less pronounced than those obtained in mice immunized with the antibody-secreting immunotherapy cells (Fig. 2a). To relate the survival data to the levels of circulating antibody, retro-orbital bleeds were performed 3 days post-immunization/antibody injection and serum antibody levels were evaluated by ELISA (Fig. 2c). High serum levels of anti-CTLA-4 were observed in mice treated with the 150 μg systemic antibody dose. In contrast, significantly lower levels of serum anti-CTLA-4 were detected in all other groups. Taken together, these data suggest that antibody secreting immunotherapy cells generate enhanced anti-tumor responses compared to the co-administration of an equivalent dose of recombinant antibody with the GM-CSF secreting tumor cell immunotherapy at the immunization site.
Local secretion of anti-CTLA-4 enhanced anti-tumor responses in the B16F10 tumor model
To confirm the results obtained in the CT26 model, the effects of local secretion of anti-CTLA-4 were also evaluated in the poorly immunogenic B16F10 model. The susceptibly of implanted tumors to anti-CTLA-4 therapy is known to be related to the inherent immunogenicity of the tumor [6, 15], and treatment of B16F10 tumors with anti-CTLA-4 monotherapy failed to show any survival benefit over HBSS-injected mice (data not shown). As observed in the CT26 model, treatment with either B16GM in combination with a high dose of systemic anti-CTLA-4 or immunization with B16GM F2A anti-CTLA-4 cells prolonged the survival of tumor-bearing mice compared to immunization with B16GM alone (Fig. 3a). Mice treated with B16GM cells, or B16GM cells combined with a low dose of either systemic or local anti-CTLA-4, demonstrated a slight survival advantage compared to HBSS-injected control animals. Administration of B16GM F2A anti-CTLA-4 cells resulted in similar serum anti-CTLA4 kinetics as observed in the CT26 model, which peaked at day 3 and gradually decreased over a period of 18 days (Fig. 3b). These data confirm that durable, local secretion of an anti-CTLA-4 improves the efficacy of GM-CSF-secreting immunotherapies at lower systemic antibody exposure. It has also recently been shown that continuous secretion of anti-CTLA-4 from live TC-1 cervical cancer cells enhanced their regression, whereas systemic and intratumoral injection of anti-CTLA-4 had no significant impact on tumor growth [16]. Taken together, these data suggest that potent anti-tumor responses can be elicited by sustained local secretion of anti-CTLA-4.

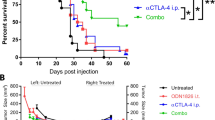
Local secretion of anti-CTLA-4 from a GM-CSF-secreting tumor cell immunotherapy enhances anti-tumor responses. a C57BL/6 mice were inoculated with live B16F10 tumor cells. The following day mice were injected with either HBSS, systemic anti-CTLA-4 (D1 = 150 μg, D4 = 100 μg), or irradiated B16GM cells. In the combination groups, mice were immunized with irradiated B16GM cells and received either a high dose (D1 = 150 μg, D4 = 100 μg) or a low dose (D1 = 15 μg, D4 = 10 μg) of systemic or local recombinant anti-CTLA-4. The final group received irradiated B16GM cells modified to secrete anti-CTLA-4. A second round of immunizations and antibody injections was administered starting on day 14. The median survival time (MST) of mice treated in the B16GM group was 80 days, whereas, the MST was undefined in the B16GM systemic anti-CTLA-4 and B16GM F2A anti-CTLA-4 groups. b Immunization with antibody-secreting cells results in sustained in vivo expression. Mice (n = 5 per group) received a single injection of 15 or 150 μg systemic anti-CTLA-4, 15 μg of local anti-CTLA-4, or 3 × 106 irradiated B16GM tumor cells engineered to express anti-CTLA-4. Retro-orbital bleeds were performed at the indicated time points, and serum anti-CTLA-4 levels were evaluated by ELISA
Anti-CTLA-4 secreting tumor cell immunotherapy resulted in potent anti-tumor responses without inducing autoimmunity associated autoantibodies
Clinical studies evaluating CTLA-4 blockade have generated promising anti-tumor responses, however, treatment with anti-CTLA-4 can in some patients result in adverse immune related events [2, 7]. One potential advantage of administering or secreting a protein or antibody locally is the reduction of toxicity or adverse immune events resulting from systemic exposure. To evaluate this hypothesis, the serum levels of several antibodies indicative of autoimmunity were determined. CTLA-4 blockade has previously been shown to increase the levels of antinuclear antibodies (ANA) in clinical studies [17–19], as well as anti-ssDNA [20] and anti-dsDNA [21] antibodies in preclinical studies. Mice were injected with HBSS, systemic anti-CTLA-4, irradiated B16GM tumor cells, or irradiated B16GM F2A anti-CTLA-4 immunotherapy cells. This treatment regime was repeated bi-weekly, and serum was collected and evaluated for the presence of autoimmune associated antibodies to ANA, ssDNA, and dsDNA by ELISA (Fig. 4). Increased levels of all evaluated autoantibodies were observed in mice treated with systemic anti-CTLA4, whereas only baseline levels were observed in serum of mice treated with B16GM F2A anti-CTLA4. These data show that compared to systemic administration of anti-CTLA-4, local administration of anti-CTLA-4 from a GM-CSF-secreting immunotherapy results in significant anti-tumor responses while inducing lower levels of autoantibodies indicative of systemic autoimmunity.

Decreased levels of autoimmunity associated antibodies are observed in mice treated with a whole tumor cell immunotherapy that locally secretes GM-CSF and anti-CTLA-4. Mice were injected with HBSS, systemic anti-CTLA-4 (D0 = 150 μg, D3 = 100 μg), 3 × 106 irradiated B16GM tumor cells, or 3 × 106 irradiated B16GM cells modified to secrete anti-CTLA-4. This treatment regime was repeated bi-weekly. At the indicated time points serum was collected and evaluated for the presence of anti-nuclear (a) anti-ssDNA (b) and anti-dsDNA (c) antibodies by ELISA. The asterisks indicate time points where statistically significant differences of autoimmunity associated antibodies were detected between the B16GM systemic anti-CTLA-4 and B16GM F2A anti-CTLA-4 groups (*P < 0.05)
CTLA-4 blockade has been shown to induce autoimmune phenomena in numerous human studies [6], and in some instances these events have been demonstrated to correlate with clinical responses [17]. However, it is unclear whether the clinical responses and IBEs can be uncoupled. The data presented here suggest that the anti-tumor and autoimmune responses may indeed be dissociated by delivering the antibody locally from a GM-CSF-secreting cancer immunotherapy. GM-CSF has a well-established role in the recruitment and activation of dendritic cells that efficiently present tumor antigens derived from the immunotherapy to T cells [3]. CTLA-4 blockade has been shown to lower the threshold for activation of weakly reacting T cell clones [6], thus sustained levels of anti-CTLA-4 at the immunization site may enhance T cell activation and reduce or eliminate IBEs associated with systemic exposure to high doses of the antibody.
References
Carter PJ (2006) Potent antibody therapeutics by design. Nat Rev Immunol 6:343–357
Peggs KS, Quezada SA, Korman AJ, Allison JP (2006) Principles and use of anti-CTLA4 antibody in human cancer immunotherapy. Curr Opin Immunol 18:206–213
Dranoff G (2003) GM-CSF-secreting melanoma vaccines. Oncogene 22:3188–3192
Nemunaitis J (2005) Vaccines in cancer: GVAX, a GM-CSF gene vaccine. Expert Rev Vaccines 4:259–274
Simmons AD, Li B, Gonzalez-Edick M, Lin C, Moskalenko M, Du T, Creson J, Vanroey MJ, Jooss K (2007) GM-CSF-secreting cancer immunotherapies: preclinical analysis of the mechanism of action. Cancer Immunol Immunother 56:1653–1665
Korman AJ, Peggs KS, Allison JP (2006) Checkpoint blockade in cancer immunotherapy. Adv Immunol 90:297–339
Sanderson K, Scotland R, Lee P, Liu D, Groshen S, Snively J, Sian S, Nichol G, Davis T, Keler T, Yellin M, Weber J (2005) Autoimmunity in a phase I trial of a fully human anti-cytotoxic T-lymphocyte antigen-4 monoclonal antibody with multiple melanoma peptides and Montanide ISA 51 for patients with resected stages III and IV melanoma. J Clin Oncol 23:741–750
Finer MH, Dull TJ, Qin L, Farson D, Roberts MR (1994) Kat: a high-efficiency retroviral transduction system for primary human T lymphocytes. Blood 83:43–50
Dull T, Zufferey R, Kelly M, Mandel RJ, Nguyen M, Trono D, Naldini L (1998) A third-generation lentivirus vector with a conditional packaging system. J Virol 72:8463–8471
Li B, Lalani AS, Harding TC, Luan B, Koprivnikar K, Huan Tu G, Prell R, VanRoey MJ, Simmons AD, Jooss K (2006) Vascular endothelial growth factor blockade reduces intratumoral regulatory T cells and enhances the efficacy of a GM-CSF-secreting cancer immunotherapy. Clin Cancer Res 12:6808–6816
Donnelly ML, Luke G, Mehrotra A, Li X, Hughes LE, Gani D, Ryan MD (2001) Analysis of the aphthovirus 2A/2B polyprotein “cleavage” mechanism indicates not a proteolytic reaction, but a novel translational effect: a putative ribosomal “skip”. J Gen Virol 82:1013–1025
Donnelly ML, Hughes LE, Luke G, Mendoza H, ten Dam E, Gani D, Ryan MD (2001) The “cleavage” activities of foot-and-mouth disease virus 2A site-directed mutants and naturally occurring “2A-like” sequences. J Gen Virol 82:1027–1041
Ryan MD, King AM, Thomas GP (1991) Cleavage of foot-and-mouth disease virus polyprotein is mediated by residues located within a 19 amino acid sequence. J Gen Virol 72(Pt 11):2727–2732
Fang J, Qian JJ, Yi S, Harding TC, Tu GH, VanRoey M, Jooss K (2005) Stable antibody expression at therapeutic levels using the 2A peptide. Nat Biotechnol 23:584–590
van Elsas A, Hurwitz AA, Allison JP (1999) Combination immunotherapy of B16 melanoma using anti-cytotoxic T lymphocyte-associated antigen 4 (CTLA-4) and granulocyte/macrophage colony-stimulating factor (GM-CSF)-producing vaccines induces rejection of subcutaneous and metastatic tumors accompanied by autoimmune depigmentation. J Exp Med 190:355–366
Tuve S, Chen BM, Liu Y, Cheng TL, Toure P, Sow PS, Feng Q, Kiviat N, Strauss R, Ni S, Li ZY, Roffler SR, Lieber A (2007) Combination of tumor site-located CTL-associated antigen-4 blockade and systemic regulatory T-cell depletion induces tumor-destructive immune responses. Cancer Res 67:5929–5939
Attia P, Phan GQ, Maker AV, Robinson MR, Quezado MM, Yang JC, Sherry RM, Topalian SL, Kammula US, Royal RE, Restifo NP, Haworth LR, Levy C, Mavroukakis SA, Nichol G, Yellin MJ, Rosenberg SA (2005) Autoimmunity correlates with tumor regression in patients with metastatic melanoma treated with anti-cytotoxic T-lymphocyte antigen-4. J Clin Oncol 23:6043–6053
Phan GQ, Yang JC, Sherry RM, Hwu P, Topalian SL, Schwartzentruber DJ, Restifo NP, Haworth LR, Seipp CA, Freezer LJ, Morton KE, Mavroukakis SA, Duray PH, Steinberg SM, Allison JP, Davis TA, Rosenberg SA (2003) Cancer regression and autoimmunity induced by cytotoxic T lymphocyte-associated antigen 4 blockade in patients with metastatic melanoma. Proc Natl Acad Sci USA 100:8372–8377
Hodi FS, Mihm MC, Soiffer RJ, Haluska FG, Butler M, Seiden MV, Davis T, Henry-Spires R, MacRae S, Willman A, Padera R, Jaklitsch MT, Shankar S, Chen TC, Korman A, Allison JP, Dranoff G (2003) Biologic activity of cytotoxic T lymphocyte-associated antigen 4 antibody blockade in previously vaccinated metastatic melanoma and ovarian carcinoma patients. Proc Natl Acad Sci USA 100:4712–4717
van Wijk F, Hoeks S, Nierkens S, Koppelman SJ, van Kooten P, Boon L, Knippels LM, Pieters R (2005) CTLA-4 signaling regulates the intensity of hypersensitivity responses to food antigens, but is not decisive in the induction of sensitization. J Immunol 174:174–179
Kocak E, Lute K, Chang X, May KF Jr, Exten KR, Zhang H, Abdessalam SF, Lehman AM, Jarjoura D, Zheng P, Liu Y (2006) Combination therapy with anti-CTL antigen-4 and anti-4-1BB antibodies enhances cancer immunity and reduces autoimmunity. Cancer Res 66:7276–7284
Acknowledgments
We thank P. Working for critical reading of the manuscript, and R. Prell for helpful discussions. B. Batiste, J. Ho, T. Langer and S. Tanciongo are gratefully acknowledged for their technical assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Simmons, A.D., Moskalenko, M., Creson, J. et al. Local secretion of anti-CTLA-4 enhances the therapeutic efficacy of a cancer immunotherapy with reduced evidence of systemic autoimmunity. Cancer Immunol Immunother 57, 1263–1270 (2008). https://doi.org/10.1007/s00262-008-0451-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00262-008-0451-3