
Abstract
Increasing evidence suggests that maintenance of homeostasis between gut microbiota and host plays an important role in human health. Many diseases, such as those affecting the liver, have been linked to imbalances in gut microbial communities. However, it is not clear whether an imbalance in gut microbiota promotes the onset of liver injury or if the imbalance results from the pathological state. In the current study, antibiotics were used to disturb the gut microbiota of both rats fed a high-cholesterol diet and rats fed a normal diet (controls). The prevalence of Bacteroidetes and Firmicutes were reduced, and Proteobacteria was greatly increased in the guts of rats after antibiotic treatment. The antibiotic-induced perturbation of gut microbiota aggravated cholesterol accumulation and liver injury in rats fed a high-cholesterol diet. This may have been due to an increase in intestinal permeability and plasma lipopolysaccharide (LPS), which lead to an increase in LPS absorption and activation of TLR4 signaling, resulting in the synthesis of pro-inflammatory cytokines and chemokines in liver tissues. This study suggests that imbalances in gut microbiota may be a predisposing factor for the onset of metabolic diseases and liver injuries related to cholesterol and high-cholesterol diets. Modulation of gut microbiota could be a novel target for preventing cholesterol-related metabolic disorders.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The human intestines contain trillions of microorganisms, of which bacteria are the most abundant. Increasing evidence suggests that maintenance of homeostasis between the gut microbiota and host plays an important role in human health and that disruption of this homeostasis promotes illness and contributes to the occurrence of various diseases (Claus et al. 2011; Clemente et al. 2012; Hooper and Gordon 2001; Owyang and Wu 2014). It is estimated that there are about 10 times more bacterial cells in the gut than the total number of human somatic and germ cells combined (Savage 1977). Members of the microbial community have been shown to play critical roles in the digestion of food, production of essential vitamins, and in shaping innate immunity (Hill 1997; Mazmanian et al. 2005; Nicholson et al. 2005).
Many diseases, such as obesity (Ley et al. 2006), diabetes (Qin et al. 2012), and inflammatory bowel disease (Frank et al. 2007), have been linked to imbalances in the gut microbial community. Recent evidence has demonstrated that dysbiotic changes in gut microbiota are common in patients with liver disease (Miyake and Yamamoto 2013; Qin et al. 2014; Schnabl and Brenner 2014), though it is not clear whether gut microbial imbalances contribute to the onset of disease or are caused by disease. A study in humans suggested that oral administration of vancomycin significantly impacted host physiology by decreasing gut microbiota diversity, bile acid metabolism, and insulin sensitivity (Vrieze et al. 2014). Furthermore, antibiotic administration during early life was found to alter the gut microbiota, as well as hepatic metabolism of lipids and cholesterol (Cho et al. 2012). These results indicate that an imbalance in gut microbiota may promote the onset of pathology and that gut microbiota may be a regulatory factor in hepatic cholesterol metabolism and liver injury.
It could be that differences in the composition of gut microbiota, resulting from perturbations in the gut microbial community, affect hepatic metabolism of cholesterol leading to liver injury. In the current study, rats were fed a high-cholesterol diet and concurrently treated with oral antibiotics to disrupt gut microbial composition, in order to investigate the relationship between changes in gut microbiota and hepatic cholesterol metabolism and liver injury and the possible mechanisms for these effects.
Materials and methods
Animals
Forty male, conventionally clean, 3 week old, Sprague–Dawley rats were purchased from Vital River Laboratory Animal, Ltd. (Beijing, China). Rats were individually housed in cages and maintained at controlled conditions of temperature (23 ± 2 °C) and humidity (55 ± 5 %) and on a 12-h light/dark cycle (lights on at 7:00 a.m.). Food and water were supplied ad libitum. All animals were cared for in accordance with the Guide for the Care and Use of Laboratory Animals. The Institutional Animal Care and Use Committee of the Institute of Psychology, Chinese Academy of Sciences approved all experiments.
High-cholesterol diet and antibiotic treatment
All rats underwent a 1-week adaptive period on a regular diet containing 32 % (weight/weight) protein, 5 % fat, 2 % fiber, 1.8 % calcium, 1.2 % phosphorus, and nitrogen-free extract as the remainder (Commercial chow, Keao Xieli Feed Co. Ltd., Beijing, China). The rats were then divided into four groups as follows: a regular diet group (CT, n = 10), a high-cholesterol diet group (HCD, n = 10), a regular diet group + antibiotic group (CT + Ab, n = 10), and a high-cholesterol diet + antibiotic group (HCD + Ab, n = 10). The CT + Ab and HCD + Ab groups received 1 g/L each of ampicillin, neomycin, and metronidazole in drinking water during the experimental period (Carvalho et al. 2012). The high-cholesterol diet contained 1.5 % (weight/weight) cholesterol, 10 % lard, 10 % sucrose, 0.3 % sodium cholate, 0.2 % propylthiouracil, and 78 % regular diet.
Intestinal microbiota analysis
Fecal samples were collected at euthanasia and frozen at −80 °C until use. Bacterial DNA was extracted using a TIANamp stool DNA kit (Tiangen Biotech Co. Ltd., Beijing, China) following the manufacturer’s instructions. The microbiota composition of the samples was established by amplicon sequencing of the V3-V5 regions of 16S rRNA gene on the Miseq system. The bioinformatic analysis was mainly performed using the QIIME pipeline (Caporaso et al. 2010). Only reads with lengths >400 bp and <480 bp, average quality scores >30, and without ambiguous bases were retained for further analysis. The resultant high-quality sequences were clustered into operational taxonomic units (OTUs) with a 97 % threshold identity using UCLUST. The taxonomy of each OTU representative sequence was assigned using the Ribosomal Database Project (RDP) classifier with an 80 % confidence limit. To evaluate the alpha diversity, the rarefaction estimators and Chao1 and Shannon diversity indexes were calculated. Genewiz Corporation (Suzhou, China) completed the sequencing and bioinformatic analysis.
Assay for blood serum indexes
Blood samples from each rat were collected via cardiac puncture just prior to euthanasia, following food deprivation for 12 h. The serum was separated from the blood by centrifugation at 3500 rpm for 10 min. The total cholesterol (TC), triglycerides (TG), high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C), alanine aminotransferase (ALT), and aspartate aminotransferase (AST) were measured with an automatic chemical analyzer 7020 (Hitachi, Tokyo, Japan). The atherogenic indexes (TC/HDL-C and LDL-C/HDL-C ratios) were also calculated. Serum endotoxin concentration was determined using a kit based on a Tachypleus Amebocyte Lysate (Chromogenic endpoint TAL kit, Tiandz Inc., Beijing, China), according to the manufacturer’s instructions.
Assay for liver TC and TG
After euthanasia, the livers of rats were removed and rinsed with physiological saline solution and blotted dry with filter paper. Liver TC and TG were determined using a tissue total cholesterol and triglyceride assay kit (Applygen Technologies Inc., Beijing), according to the manufacturer’s instructions.
Liver histopathology
Part of each liver was fixed in 10 % (volume/volume) formaldehyde, embedded in paraffin, and stained with hematoxylin-eosin (HE). Another part of each liver was rapidly frozen to about −20 to −30 °C and then cut for oil red O staining. A masked, expert pathologist evaluated three different high-power areas of the histologic sections.
Real-time quantitative PCR
Total RNA from each individual liver was isolated via TRIzol reagent (Tiangen Biotech Co. Ltd., Beijing, China), according to the manufacturer’s protocol. A UV spectrophotometer, at 260 and 280 nm, was used to estimate RNA concentration, which was then transcribed into cDNA (Tiangen Biotech Co. Ltd., Beijing, China). Expression of messenger RNA (mRNA) was measured by RT-PCR using the RealStar Power SYBR mixture (GenStar BioSolutions Co. Ltd., Beijing, China), with values presented as 2−ΔΔCT. The housekeeping gene for glyceraldehyde-3-phosphate dehydrogenase (GAPDH; EC1.2.1.12) was used for normalization. The genes and primer sequences used for RT-PCR are described in Table 1.
Statistical analysis
The experimental data are presented as mean ± standard deviation (SD). The statistical significance of differences was analyzed by one-way ANOVA followed by a post hoc Bonferroni’s multiple comparison test using SPSS 17.0 statistical software (SPSS Inc., Chicago, IL, USA). Values of p < 0.05 were considered statistically significant.
Accession numbers of sequence data
The Miseq sequencing data obtained in this study were deposited in the NCBI Sequence Read Archive database with accession numbers SRR2005927 and SRR2005929 to SRR2005939.
Results
Antibiotic treatment changed composition of gut microbiota in rats fed a high-cholesterol diet
Intestinal microbiota sequencing analysis from fecal samples of each group displayed differences in the bacterial profiles between CT and HCD rats. The proportion of Bacteroidetes was higher in HCD rats compared to CT rats. In contrast, the prevalence of Firmicutes was lower in HCD rats than in CT rats. In one group of HCD and one group of CT rats, broad-spectrum antibiotics were used to modulate the gut microbiota community in order to explore the role of gut microbiota composition in lipid metabolism changes induced by a high-cholesterol diet. Treatment with antibiotics for 4 weeks led to major changes in the gut bacterial profiles of both CT + Ab and HCD + Ab rats. The proportion of Bacteroidetes and Firmicutes were greatly reduced after antibiotic treatment compared to the rats not treated with antibiotics. There was a huge increase in the proportion of Proteobacteria after antibiotic treatment in both the CT and HCD groups (Fig. 1a).

Antibiotic treatment changed the gut microbiota profile. a Comparison of gut microbiota composition at the phylum level. b Comparison of gut microbiota composition at the genus level. c Phylotype richness among groups. d Chao1. e Shannon diversity analysis. CT control group, CT + Ab antibiotic treatment only, HCD high-cholesterol group, HCD + Ab high-cholesterol and antibiotic treatment group
The differences were further distinguished via analysis of the microbial flora at the genus level (Fig. 1b). More Bacteroides, Parabacteroides, and Prevotella were found in HCD rats compared to CT rats. In addition, more Achromobacter, Parasutterella, Escherichia/Shigella, Proteus, and Stenotrophomonas were found after antibiotic treatment in HCD rats.
To measure how phylotype richness in the four groups varied with sample size, the phylotype rarefaction curves were calculated at 97 % similarity levels. Different phylotype richness was observed among groups (Fig. 1c). The Chao1 and Shannon diversity analysis suggested that species diversity was different between groups (Fig. 1d, e). These data suggest that the gut microbiota profile in HCD rats was considerably altered as a consequence of antibiotic treatment.
Antibiotic treatment aggravated the serum cholesterol elevation induced by the high-cholesterol diet
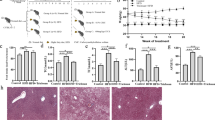
Serum from CT and CT + Ab rats was transparent; however, serum from both HCD and HCD + Ab rats was milky white. Serum from HCD + Ab rats appeared to be even more milky white when compared with HCD rats (Fig. 2a). The absorbance of sera was measured at 600 nm in order to objectively distinguish differences among the groups. There was no difference in the A600 values between CT and CT + Ab groups. The A600 values in the HCD group were significantly higher than in CT rats. The A600 values were higher in HCD + Ab rats than in HCD rats (Fig. 2b).

Antibiotic treatment increased serum cholesterol accumulation in HCD + Ab rats. a Representative photos of serum from each group. b The absorbance of sera at 600 nm. c Total cholesterol (TC). d Triglyceride (TG). e High-density lipoprotein cholesterol (HDL-C). f Low-density lipoprotein cholesterol (LDL-C). g TC/HDL-C ratios. h LDL-C/HDL-C ratios. The data are shown as the mean ± standard deviation. (a, b, c) Mean values in each panel with different letters differ significantly (p < 0.05). CT control group, CT + Ab antibiotic treatment only, HCD high-cholesterol group, HCD + Ab high-cholesterol and antibiotic treatment group
The high-cholesterol diet induced a significant increase in serum TC and LDL-C levels but did not significantly influence the levels of serum TG and HDL-C. Serum lipid accumulation was greater in the HCD + Ab group than in the other three groups. Serum TC, TG, HDL-C, and LDL-C levels were significantly higher in HCD + Ab rats compared with HCD rats; however, there were no effects on these parameters in CT + Ab group compared to CT group (Fig. 2c–f). TC/HDL-C and LDL-C/HDL-C ratios are strong risk factors for predicting cardiovascular disease (CVD). The high-cholesterol diet induced a significant elevation in the TC/HDL-C and LDL-C/HDL-C ratios, thus increasing the risk of CVD. HCD + Ab rats had a significantly elevated CVD risk compared with HCD rats (Fig. 2g, h). These results suggest that modulation of gut microbiota by antibiotics likely increased serum lipid accumulation in HCD + Ab rats.
Antibiotic treatment worsened liver injuries induced by the high-cholesterol diet
The high-cholesterol diet induced accumulation of total cholesterol and triglycerides in the liver. Treatment with antibiotics aggravated cholesterol accumulation, but not triglyceride accumulation, in the livers of HCD rats. Antibiotic treatment did not influence cholesterol or triglyceride accumulation in the livers of CT rats (Fig. 3a). Elevated serum ALT levels were found in HCD rats and even higher in HCD + Ab rats than in HCD rats indicating that the high-cholesterol diet induced liver injury and that antibiotic treatment intensified this effect. However, serum AST levels were not significantly different among the groups (Fig. 3b). These results suggest that antibiotic treatment aggravated cholesterol accumulation in the liver and worsened liver injuries in the HCD + Ab rats. Histopathological alterations in the liver tissues confirmed these results. Hematoxylin-eosin (HE) and oil red O staining showed the differences in liver tissue structure and lipid accumulation among the four groups (Fig. 3c, d). The livers of CT rats had well-organized structures. Hepatic sinusoids were clearly visible, and hepatic cords were neatly arranged and distributed radially around the central veins. There were no significant structural changes except for some mild fatty degeneration and hepatic congestion in the CT + Ab rats. There was a moderate degree of liver structure damage in HCD rats. In addition, hepatocytes of HCD rats showed significant hepatic steatosis, cell necrosis, vacuolization, and increased lipids deposition in the cytoplasm. The above effects were aggravated by the antibiotic treatment in the HCD + Ab rats.

Antibiotic treatment aggravated liver cholesterol accumulation and liver injury in HCD rats. a Liver total cholesterol and triglycerides. b Serum ALT and AST levels. c Representative photomicrographs of hematoxylin and eosin staining. d Representative photomicrographs of oil red O staining. The data are shown as the mean ± standard deviation. (a, b, c) Different letters indicate significant differences between mean values in each panel (p < 0.05). CT control group, CT + Ab antibiotic treatment only, HCD high-cholesterol group, HCD + Ab high-cholesterol and antibiotic treatment
Antibiotic treatment increased the occurrence of liver inflammation in rats fed a high-cholesterol diet
Histopathological analysis suggested that liver injury and inflammation occurred in rats fed a high-cholesterol diet. The mRNA levels of inflammation markers TNF-α, IL-1β, IL-6, iNOS, and COX-2 were quantified in liver tissues (Fig. 4a–e). The mRNA concentrations of TNF-α, iNOS, and COX-2 were significantly higher in HCD rats compared with CT fed rats, but there were no significant differences in mRNA levels of IL-1β and IL-6. The increases in TNF-α, iNOS, and COX-2 levels were significantly greater in HCD + Ab rats, and mRNA levels of IL-1β and IL-6 were also significantly higher in HCD + Ab rats compared with HCD and CT rats. Similar results were observed in chemokine MCP-1 mRNA levels (Fig.4f). MCP-1 mRNA expression was significantly increased in HCD + Ab rats compared with HCD and CT rats. There was no difference in MCP-1 mRNA expression when comparing HCD to CT rats. F4/80 is a marker, which is specific for mature macrophages. No significant change in F4/80 mRNA expression was found in HCD + Ab compared with HCD or CT rats (Fig. 4g). Liver histopathology also suggested hepatic cell necrosis in HCD rats. The mRNA expression of pro-apoptotic Bax and anti-apoptotic Bcl-2 mRNA was also analyzed in the liver tissue (Fig. 4h, i). There was no difference in Bax mRNA expression; however, Bcl-2 mRNA expression was significantly increased in HCD compared to CT rats. The levels of both Bax and Bcl-2 were significantly increased in HCD + Ab rats compared with HCD and CT rats. These results suggest a possible link between the gut microbiota and liver inflammation in rats fed a high-cholesterol diet.

Antibiotic treatment promoted the mRNA expression of liver injury markers in rats fed a high-cholesterol diet. Inflammation markers: a TNF-α, b IL-1β, c IL-6, d inos, e COX-2; macrophage infiltration markers: f MCP-1, g F4/80; apoptosis markers: h ax, i Bcl-2 mRNA concentrations in liver tissue. The data are shown as the mean ± standard deviation. (a, b, c, d) Different letters indicate significant differences between mean values in each panel (p < 0.05). CT control group, CT + Ab antibiotic treatment only, HCD high-cholesterol group, HCD + Ab high-cholesterol and antibiotic treatment
Antibiotic treatment increased metabolic endotoxemia in rats fed a high-cholesterol diet
Increased levels of serum endotoxin (bacterial lipopolysaccharide, LPS) were found in HCD compared with CT rats. This increase in LPS was aggravated by antibiotic treatment (Fig. 5a). The expression of the intestinal epithelial tight junction protein, occludin, was significantly reduced in HCD compared to CT rats. Occludin expression was reduced significantly more in HCD + Ab rats compared to HCD and CT rats (Fig. 5b). These data suggest that gut microbiota may be involved in modulating intestinal epithelial tight junction protein expression, thus influencing intestinal permeability and, in turn, playing a role in metabolic endotoxemia.

Antibiotic treatment increased metabolic endotoxemia and promoted the activation of the TLR4 signaling pathway in rats fed a high-cholesterol diet. a Serum endotoxin levels. b mRNA expression of the intestinal epithelial tight junction protein, occludin. c HMGB1. d TLR4. e NF-κB mRNA expressions in liver tissue. The data are shown as the mean ± standard deviation. (a, b, c, d) Different letters indicate significant differences between mean values in each panel (p < 0.05). CT control group, CT + Ab antibiotic treatment only, HCD high-cholesterol group, HCD + Ab high-cholesterol and antibiotic treatment
Antibiotic treatment promoted the activation of the toll-like receptor 4 (TLR4) pathway in rats fed a high-cholesterol diet
The mRNA expression of genes related to TLR4 signaling (high-mobility group box-1(HMGB1), TLR4, and NF-κB) were quantified in liver tissue (Fig. 5c–e). The mRNA expression levels of TLR4 and NF-κB were significantly higher in HCD rats compared with CT rats. Expressions of TLR4 and NF-κB were increased significantly more in HCD + Ab compared with HCD and CT rats. The high-cholesterol diet did not induce significant changes in HMGB1 expression; however, expression of HMGB1 was significantly higher in HCD + Ab compared to CT rats. There was no significant difference in the level of HMGB1 mRNA between HCD + Ab and HCD rats.
Discussion
The gut microbiota is an immense ecosystem, which inhabits the gastrointestinal tract of the host. The microorganisms that make up the gut microbiota are considered to be important to the host because of their numerous functions and contributions to the physiology of the gastrointestinal tract. However, the role of the gut microbiota and the effects of gut microbiota perturbations in various disease states have not yet been fully elucidated.
In this study, it was shown that a high-cholesterol diet changed the gut microbial profile in rats. The percentage of Bacteroidetes was higher (72.76 vs. 53.54 %) and the percentage of Firmicutes was lower (18.80 vs. 36.96 %) in HCD rats compared to CT rats. These results are similar to those reported in previous studies comparing gut microbiota profiles in non-alcoholic steatohepatitis (NASH) patients and healthy controls (Zhu et al. 2013). There was a vast difference in gut microbiota profiles in rats treated with antibiotics. The prevalence of Proteobacteria was higher (82.69 %), and the number of Bacteroidetes and Firmicutes were lower in HCD + Ab rats. These results demonstrate significant alterations in the composition of gut microbiota as a result of treatment with antibiotics.
Alterations of the gut microbiota induced by diet was considered to exert a strong influence on the development of metabolic syndrome, and alterations of the gut microbiota induced by high-fat diet is strongly linked to oxidative stress in mice (Qiao et al. 2013). In this study, the results indicate that perturbation of the gut microbiota by antibiotics caused the rats to be more sensitive to the high-cholesterol diet. Serum and liver cholesterol levels, as well as the incidence of liver injury, were higher in HCD rats compared with CT rats. When antibiotics were used to perturb the gut microbiota of HCD rats, the serum and liver cholesterol levels, as well as incidence of liver injuries, were significantly higher than in HCD rats not treated with antibiotics. However, antibiotic treatment alone did not affect serum and liver cholesterol levels and/or lead to liver injuries. These results suggest that the gut microbiota may have played a role in the changes in lipid metabolism, which were induced by the high-cholesterol diet, in rats.
TC/HDL-C and LDL-C/HDL-C ratios are recognized as two important risk factors for predicting cardiovascular disease (CVD) and have greater predictive value than using isolated parameters independently (Millan et al. 2009). In the current study, the high-cholesterol diet induced significant increases in both the TC/HDL-C and LDL-C/HDL-C ratios, suggesting that the high-cholesterol diet significantly increased the risk of CVD. HCD + Ab rats had a significantly heightened CVD risk when compared to HCD rats, though antibiotic treatment alone also did not affect the TC/HDL-C and LDL-C/HDL-C ratios. These results suggest that the gut microbiota might be involved in high-cholesterol diet-related CVDs. In general, this study suggests that antibiotic-induced imbalances in gut microbiota might be a predisposing factor for the onset of cholesterol-related metabolic diseases in those who consume a high-cholesterol diet. However, these results are contrary to those reported in previous studies (Cani et al. 2008; Carvalho et al. 2012; Membrez et al. 2008). Cani et al. reported that changes in gut microbiota induced by antibiotics reduced metabolic endotoxemia in mice fed a high-fat diet (Cani et al. 2008) and Membrez et al. found that modulation of gut microbiota by antibiotics enhanced glucose tolerance in mice (Membrez et al. 2008). Similarly, Carvalho and colleagues found that gut microbiota modulation with antibiotics improved insulin signaling in mice fed a high-fat diet (Carvalho et al. 2012). These studies indicate that antibiotic-induced perturbations of gut microbiota might protect the host from metabolic diseases. The difference in the results may be associated with type and the age of animals used in the studies. Rats were used for the current study, while mice were used in the previous studies. Furthermore, in previous studies, animals were treated with antibiotics beginning at 12 weeks (Cani et al. 2008), 8 to 10 weeks (Membrez et al. 2008), and 6 weeks of age (Carvalho et al. 2012). In the current study, rats were treated with antibiotics beginning at 4 weeks of age. In addition, the types and dosage of antibiotics used might have contributed to differences in the results. Cho et al. reported that antibiotics in early life (3 weeks old) altered the murine colonic microbiome and increased adiposity (Cho et al. 2012). This suggests that early-life imbalances of gut microbiota modulated by antibiotics preceded the metabolic disease pathogenesis. The gut microbiota present in early life impacts our immune development, and dysbiosis in early life plays an important role in disease pathogenesis (Arrieta et al. 2014). The above results suggest that the gut microbiota plays an important role in host health and disease status, whether antibiotic-induced perturbations of gut microbiota precede the onset of metabolic disease or protect the host from disease pathogenesis. In any case, either way, further research is needed.
Dietary factors are considered to be important determinants of liver disease progression (Ioannou et al. 2009). High-cholesterol diets usually cause liver injuries, liver inflammation, and hepatocyte apoptosis (Kainuma et al. 2006; Sumiyoshi et al. 2010; Vergnes et al. 2003). In this study, histopathological analysis suggested that hepatic steatosis and liver inflammation occurred in rats fed a high-cholesterol diet. Antibiotic treatment aggravated liver injuries that occurred as a result of the high-cholesterol diet. Analysis of mRNA expression also demonstrated that antibiotic treatment further increased the expression of the pro-inflammatory factors, TNF-α, IL-1β, IL-6, iNOS, and COX-2, which were induced by the high-cholesterol diet. Similar results were also observed in the alterations of the mRNA concentrations of chemokine MCP-1, the marker specific for macrophage infiltration, F4/80, and the apoptotic related Bax and Bcl-2. These results suggest that liver inflammation, macrophage infiltration, and hepatic apoptosis are induced by a high-cholesterol diet and aggravated by treatment with antibiotics. Gut microbiota might play a role in liver injuries induced by a high-cholesterol diet.
Bacterial lipopolysaccharide (LPS) is considered to be a triggering factor for metabolic diseases. High-fat diets chronically increase plasma LPS concentration and lead to metabolic endotoxemia (Cani et al. 2007). In the current study, the high-cholesterol diet increased the level of plasma LPS and this was aggravated by antibiotic treatment. This may be associated with the high prevalence of Gram-negative gut bacteria (Proteobacteria). In addition, increased intestinal permeability also contributed to increased LPS absorption. Antibiotic treatment induced a more significant reduction in expression of occludin, an intestinal epithelial tight junction protein. These results indicate that gut microbiota may participate in modulating intestinal epithelial tight junction protein expression in HCD rats, influencing intestinal permeability, leading to metabolic endotoxemia.
TLR4 is an essential receptor for the recognition of LPS (Poltorak et al. 1998). Previous studies have shown that TLR4 participates in the progression of chronic liver disease (Mencin et al. 2009) and that TLR4 signaling is pivotal during the early stages of liver injuries induced by high-fat diets (Li et al. 2011). HMGB1 is an early mediator of injury and inflammation (Scaffidi et al. 2002) and serves as a positive component, mediating TLR4 activation (Li et al. 2011). HMGB1 interacts with TLR4 to induce production of pro-inflammatory cytokines and chemokines by inducing NF-κB signaling (Park et al. 2006; van Zoelen et al. 2009). In this study, antibiotic treatment increased high-cholesterol diet-induced HMGB1, TLR4, and NF-κB expression. These results indicate that modulation of gut microbiota via antibiotic treatment significantly increased the level of plasma LPS and activated TLR4 signaling in the HCD rats, which could lead to a more serious degree of hepatic steatosis in the liver tissues of these rats.
Taken together, the high-cholesterol diet induced alterations in the gut microbiota profile and increased intestinal permeability, which led to an increase in plasma LPS. Increased LPS absorption might activate TLR4 signaling and promote the synthesis of pro-inflammatory cytokines and chemokines in the liver tissues, eventually leading to liver injuries. The cholesterol metabolic disorders observed in the current study were all aggravated by antibiotic-induced perturbation of gut microbiota in HCD rats. These results suggest that gut microbiota modulation might be a novel target for disease prevention and that modification of gut microbiota may be beneficial for preventing cholesterol metabolic disorders.
References
Andersson A, Covacu R, Sunnemark D, Danilov AI, Dal Bianco A, Khademi M, Wallstrom E, Lobell A, Brundin L, Lassmann H, Harris RA (2008) Pivotal advance: HMGB1 expression in active lesions of human and experimental multiple sclerosis. J Leukoc Biol 84(5):1248–1255
Arrieta MC, Stiemsma LT, Amenyogbe N, Brown EM, Finlay B (2014) The intestinal microbiome in early life: health and disease. Front Immunol 5:427
Batista Jr ML, Neves RX, Peres SB, Yamashita AS, Shida CS, Farmer SR, Seelaender M (2012) Heterogeneous time-dependent response of adipose tissue during the development of cancer cachexia. J Endocrinol 215(3):363–373
Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, Neyrinck AM, Fava F, Tuohy KM, Chabo C, Waget A, Delmee E, Cousin B, Sulpice T, Chamontin B, Ferrieres J, Tanti JF, Gibson GR, Casteilla L, Delzenne NM, Alessi MC, Burcelin R (2007) Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 56(7):1761–1772
Cani PD, Bibiloni R, Knauf C, Waget A, Neyrinck AM, Delzenne NM, Burcelin R (2008) Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 57(6):1470–1481
Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, Fierer N, Pena AG, Goodrich JK, Gordon JI, Huttley GA, Kelley ST, Knights D, Koenig JE, Ley RE, Lozupone CA, McDonald D, Muegge BD, Pirrung M, Reeder J, Sevinsky JR, Tumbaugh PJ, Walters WA, Widmann J, Yatsunenko T, Zaneveld J, Knight R (2010) QIIME allows analysis of high-throughput community sequencing data. Nat Methods 7(5):335–336
Carvalho BM, Guadagnini D, Tsukumo DM, Schenka AA, Latuf-Filho P, Vassallo J, Dias JC, Kubota LT, Carvalheira JB, Saad MJ (2012) Modulation of gut microbiota by antibiotics improves insulin signalling in high-fat fed mice. Diabetologia 55(10):2823–2834
Cho I, Yamanishi S, Cox L, Methe BA, Zavadil J, Li K, Gao Z, Mahana D, Raju K, Teitler I, Li HL, Alekseyenko AV, Blaser MJ (2012) Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature 488(7413):621–626
Claus SP, Ellero SL, Berger B, Krause L, Bruttin A, Molina J, Paris A, Want EJ, de Waziers I, Cloarec O, Richards SE, Wang Y, Dumas ME, Ross A, Rezzi S, Kochhar S, Van Bladeren P, Lindon JC, Holmes E, Nicholson JK (2011) Colonization-induced host-gut microbial metabolic interaction. Mbio 2(2):e00271-10
Clemente JC, Ursell LK, Parfrey LW, Knight R (2012) The impact of the gut microbiota on human health: an integrative view. Cell 148(6):1258–1270
Frank DN, Amand ALS, Feldman RA, Boedeker EC, Harpaz N, Pace NR (2007) Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc Natl Acad Sci U S A 104(34):13780–13785
Hill MJ (1997) Intestinal flora and endogenous vitamin synthesis. Eur J Cancer Prev 6:S43–S45
Hooper LV, Gordon JI (2001) Commensal host-bacterial relationships in the gut. Science 292(5519):1115–1118
Hu M, Wang Z, Rao J, Cao Y, Jiang W, Zhang F, Li X, Wang X (2010) Inhibition of inducible nitric oxide synthase worsens liver damage regardless of lipopolysaccharide treatment in small-for-size liver transplantation. Transpl Immunol 23(1–2):6–11
Ioannou GN, Morrow OB, Connole ML, Lee SP (2009) Association between dietary nutrient composition and the incidence of cirrhosis or liver cancer in the United States population. Hepatology 50(1):175–184
Kainuma M, Fujimoto M, Sekiya N, Tsuneyama K, Cheng C, Takano Y, Terasawa K, Shimada Y (2006) Cholesterol-fed rabbit as a unique model of nonalcoholic, nonobese, non-insulin-resistant fatty liver disease with characteristic fibrosis. J Gastroenterol 41(10):971–980
Ley RE, Turnbaugh PJ, Klein S, Gordon JI (2006) Microbial ecology—human gut microbes associated with obesity. Nature 444(7122):1022–1023
Li L, Chen L, Hu L, Liu Y, Sun HY, Tang J, Hou YJ, Chang YX, Tu QQ, Feng GS, Shen F, Wu MC, Wang HY (2011) Nuclear factor high-mobility group Box1 mediating the activation of toll-like receptor 4 signaling in hepatocytes in the early stage of nonalcoholic fatty liver disease in mice. Hepatology 54(5):1620–1630
Long JP, Tong HH, Shannon PA, DeMaria TF (2003) Differential expression of cytokine genes and inducible nitric oxide synthase induced by opacity phenotype variants of Streptococcus pneumoniae during acute otitis media in the rat. Infect Immun 71(10):5531–5540
Mazmanian SK, Liu CH, Tzianabos AO, Kasper DL (2005) An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 122(1):107–118
Membrez M, Blancher F, Jaquet M, Bibiloni R, Cani PD, Burcelin RG, Corthesy I, Mace K, Chou CJ (2008) Gut microbiota modulation with norfloxacin and ampicillin enhances glucose tolerance in mice. FASEB J 22(7):2416–2426
Mencin A, Kluwe J, Schwabe RF (2009) Toll-like receptors as targets in chronic liver diseases. Gut 58(5):704–720
Millan J, Pinto X, Munoz A, Zuniga M, Rubies-Prat J, Pallardo LF, Masana L, Mangas A, Hernandez-Mijares A, Gonzalez-Santos P, Ascaso JF, Pedro-Botet J (2009) Lipoprotein ratios: physiological significance and clinical usefulness in cardiovascular prevention. Vasc Health Risk Manag 5:757–765
Miyake Y, Yamamoto K (2013) Role of gut microbiota in liver diseases. Hepatol Res 43(2):139–146
Nicholson JK, Holmes E, Wilson ID (2005) Gut microorganisms, mammalian metabolism and personalized health care. Nat Rev Microbiol 3(5):431–438
Owyang C, Wu GD (2014) The gut microbiome in health and disease. Gastroenterology 146(6):1433–1436
Park JS, Gamboni-Robertson F, He QB, Svetkauskaite D, Kim JY, Strassheim D, Sohn JW, Yamada S, Maruyama I, Banerjee A, Ishizaka A, Abraham E (2006) High mobility group box 1 protein interacts with multiple toll-like receptors. Am J Physiol Cell Physiol 290(3):C917–C924
Poltorak A, He XL, Smirnova I, Liu MY, Van Huffel C, Du X, Birdwell D, Alejos E, Silva M, Galanos C, Freudenberg M, Ricciardi-Castagnoli P, Layton B, Beutler B (1998) Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science 282(5396):2085–2088
Qiao Y, Sun J, Ding YY, Le GW, Shi YH (2013) Alterations of the gut microbiota in high-fat diet mice is strongly linked to oxidative stress. Appl Microbiol Biotechnol 97(4):1689–1697
Qin JJ, Li YR, Cai ZM, Li SH, Zhu JF, Zhang F, Liang SS, Zhang WW, Guan YL, Shen DQ, Peng YQ, Zhang DY, Jie ZY, Wu WX, Qin YW, Xue WB, Li JH, Han LC, Lu DH, Wu PX, Dai YL, Sun XJ, Li ZS, Tang AF, Zhong SL, Li XP, Chen WN, Xu R, Wang MB, Feng Q, Gong MH, Yu J, Zhang YY, Zhang M, Hansen T, Sanchez G, Raes J, Falony G, Okuda S, Almeida M, LeChatelier E, Renault P, Pons N, Batto JM, Zhang ZX, Chen H, Yang RF, Zheng WM, Li SG, Yang HM, Wang J, Ehrlich SD, Nielsen R, Pedersen O, Kristiansen K, Wang J (2012) A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 490(7418):55–60
Qin N, Yang FL, Li A, Prifti E, Chen YF, Shao L, Guo J, Le Chatelier E, Yao J, Wu LJ, Zhou JW, Ni SJ, Liu L, Pons N, Batto JM, Kennedy SP, Leonard P, Yuan CH, Ding WC, Chen YT, Hu XJ, Zheng BW, Qian GR, Xu W, Ehrlich SD, Zheng SS, Li LJ (2014) Alterations of the human gut microbiome in liver cirrhosis. Nature 513(7516):59–64
Savage DC (1977) Microbial ecology of the gastrointestinal tract. Annu Rev Microbiol 31:107–133
Scaffidi P, Misteli T, Bianchi ME (2002) Release of chromatin protein HMGB1 by necrotic cells triggers inflammation. Nature 418(6894):191–195
Schnabl B, Brenner DA (2014) Interactions between the intestinal microbiome and liver diseases. Gastroenterology 146(6):1513–1524
Sumiyoshi M, Sakanaka M, Kimura Y (2010) Chronic intake of a high-cholesterol diet resulted in hepatic steatosis, focal nodular hyperplasia and fibrosis in non-obese mice. Br J Nutr 103(3):378–385
van Zoelen MAD, Yang H, Florquin S, Meijers JCM, Akira S, Arnold B, Nawroth PP, Bierhaus A, Tracey KJ, van der Poll T (2009) Role of toll-like receptors 2 and 4, and the receptor for advanced glycation end products in high-mobility group box 1-induced inflammation in vivo. Shock 31(3):280–284
Vergnes L, Phan J, Strauss M, Tafuri S, Reue K (2003) Cholesterol and cholate components of an atherogenic diet induce distinct stages of hepatic inflammatory gene expression. J Biol Chem 278(44):42774–42784
Vrieze A, Out C, Fuentes S, Jonker L, Reuling I, Kootte RS, van Nood E, Holleman F, Knaapen M, Romijn JA, Soeters MR, Blaak EE, Dallinga-Thie GM, Reijnders D, Ackermans MT, Serlie MJ, Knop FK, Holst JJ, van der Ley C, Kema IP, Zoetendal EG, de Vos WM, Hoekstra JBL, Stroes ES, Groen AK, Nieuwdorp M (2014) Impact of oral vancomycin on gut microbiota, bile acid metabolism, and insulin sensitivity. J Hepatol 60(4):824–831
Wang JC, Zhao Y, Chen SJ, Long J, Jia QQ, Zhai JD, Zhang Q, Chen Y, Long HB (2013) AOPPs induce MCP-1 expression by increasing ROS-mediated activation of the NF-kappaB pathway in rat mesangial cells: inhibition by sesquiterpene lactones. Cell Physiol Biochem 32(6):1867–1877
Xia XM, Wang FY, Wang ZK, Wan HJ, Xu WA, Lu H (2010) Emodin enhances alveolar epithelial barrier function in rats with experimental acute pancreatitis. World J Gastroenterol 16(24):2994–3001
Xiao J, Ching YP, Liong EC, Nanji AA, Fung ML, Tipoe GL (2013) Garlic-derived S-allylmercaptocysteine is a hepato-protective agent in non-alcoholic fatty liver disease in vivo animal model. Eur J Nutr 52(1):179–191
Zhang TJ, Hang J, Wen DX, Hang YN, Sieber FE (2006) Hippocampus bcl-2 and bax expression and neuronal apoptosis after moderate hypothermic cardiopulmonary bypass in rats. Anesth Analg 102(4):1018–1025
Zhang W, Wang LW, Wang LK, Li X, Zhang H, Luo LP, Song JC, Gong ZJ (2013) Betaine protects against high-fat-diet-induced liver injury by inhibition of high-mobility group box 1 and toll-like receptor 4 expression in rats. Dig Dis Sci 58(11):3198–3206
Zhu LX, Baker SS, Gill C, Liu WS, Alkhouri R, Baker RD, Gill SR (2013) Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: a connection between endogenous alcohol and NASH. Hepatology 57(2):601–609
Acknowledgments
We thank LetPub (www.letpub.com) for its linguistic assistance during the preparation of this manuscript.
Grants
The study was granted by NS Bio Japan and NS Bio Guangzhou.
Conflicts of interest
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Additional information
Xu Hu and Tao Wang contributed to the work equally and should be regarded as co-first authors.
Rights and permissions
About this article

Cite this article
Hu, X., Wang, T., Liang, S. et al. Antibiotic-induced imbalances in gut microbiota aggravates cholesterol accumulation and liver injuries in rats fed a high-cholesterol diet. Appl Microbiol Biotechnol 99, 9111–9122 (2015). https://doi.org/10.1007/s00253-015-6753-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-015-6753-4