
Abstract
Most screening approaches produce compounds that target survival genes and are likely to generate resistance over time. Simply having more drugs does not address the potential emergence of resistance caused by target mutation, drug efflux pumps over-expression, and so on. There is a great need to explore new strategies to treat fungal infections caused by drug-resistant pathogens. In this study, we found that azole-resistant Candida albicans with CaCDR1 and CaCDR2 over-expression is hypersensitive against amphotericin B (AmB) by our high throughput synergy screening (HTSS). In contrast, Δcdr1 and Δcdr2 knockout strains were resistant to AmB. Moreover, clinical isolates with increased expression of CaCDR1 and CaCDR2 demonstrated susceptibility to AmB, which can also synergize with the efflux pumps inducer fluphenazine (FPZ). Finally, the increased drug susceptibility to AmB in azole-resistant C. albicans with drug efflux pumps over-expression was consistent with the elevated expression of CaERG11 and its associated ergosterols in clinical isolates. Our data implies that the level of ergosterol contents determines the susceptibility to azoles and AmB in C. albicans. Deep understanding of the above mechanisms would offer new hope to treat drug-resistant C. albicans.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The opportunistic pathogen Candida albicans is one of the leading causes of fungal infections in humans, especially in immunocompromised patients (Zhang et al. 2007). C. albicans infections are usually treated with antifungal agents such as azoles, echinocandins, and polyene drugs. Widespread over-use of azole drugs results in the occurrence of drug-resistant isolates (Gallagher et al. 1992; Redding et al. 1994; White et al. 1998), often due to the over-expression of membrane efflux pumps (Goffeau 2008). There is a need to discover novel drugs that are less likely to be made ineffective by fungal pathogen evolution, which can counteract the drug efflux pumps, enhance the useful lifespan of anti-fungal agents, and reduce the frequency of treatment failures.
The drug efflux pumps CaCDR1 (Prasad R et al. 1995) and CaCDR2 (Sanglard et al. 1997)—C andida Drug Resistance (CDR)—are from the ATP-Binding Cassette (ABC) family, while CaMDR1 [Multi Drug Resistance (MDR); Ben-Yaacov et al. 1994] is a major facilitator in Candida. The drug efflux pumps CaCDR1 and CaMDR1 are distributed on the plasma membrane (Pasrija et al. 2005). The composition of membrane, particularly, ergosterol and sphingolipid, determines the localization and function of ABC transporters which are only accumulated in the cell plasma when the biosynthesis pathway of ergosterol or sphingolipid is disrupted (Mukhopadhyay et al. 2002, 2004; Pasrija et al. 2005). By screening our proprietary natural product library (Zhang et al. 2007), some natural products, such as berberine and AmB, are identified to specifically target the azole-resistant strains caused by drug efflux pump over-expression. In this study, we try to explore the potential mechanism of AmB on azole-resistant strains. We found that inhibitors of ergosterol biosynthesis (azoles) lost activity due to over-expression of drug efflux pumps, but the cells unexpectedly increased their ergosterol contents, and the polyenes which bind to ergosterol become more effective to kill the resistant pathogens.
Material and methods
Anti-fungal agents
Ketoconazole (KTC) and fluconazole (FLC) were purchased from the local chemical pharmacy. Amphotericin B (AmB), fluphenazine (FPZ), filipin and ergosterol (98 % purity) were purchased from Sigma-Aldrich, Germany. All compounds were dissolved in dimethyl sulfoxide (DMSO) and stored at −20 °C.
Strains and culture
All strains used in this study are listed in Table 1. The C. albicans wild type (WT) is identical to SC5314 also known as ATCC MYA-2876 (Gillum et al. 1984) except that ura3 is deleted (Fonzi and Irwin 1993). Strains Δcdr1 and Δcdr2 each lack specific pumps (Sanglard et al. 1996, 1997). Two ergosterol biosynthesis disruption mutants erg11/erg11 and erg11/erg11 erg3/erg3 were used in this study (Sanglard et al. 2003). The clinical strains #1, #4, and #17 were isolated from the same HIV patient over 2 years of increased FLC usage, which were selected for azole-resistant progeny due to an increased expression in drug efflux pumps (White 1997). Saccharomyces cerevisiae strain AD1-8u- is the parental strain with several transporter deletions (Decottignies et al. 1998). The strains AD/CaCDR1 and AD/CaCDR2 were heterologously expressed with CaCDR1 and CaCDR2, respectively (Lamping et al. 2007). All strains (Table 1) were maintained on YEPD agar (1 % yeast extract/2 % peptone/2 % dextrose/2 % agar) medium unless specified and incubated at 35 °C overnight.
Spot assays
The starting concentration of cells was 1 × 107 cells/ml counted by hemocytometer and serially diluted to 1 × 103 cells/ml. 2 μl of each suspension was spotted on YEPD plates supplemented with either different concentrations of test drugs or solvent controls. Cultures were incubated at 35 °C for 24 h (C. albicans) or 48 h (S. cerevisiae).
Anti-fungal susceptibility test and growth measurement
Drug susceptibility measurements were carried out in flat bottom, 96-well microtiter plates (Greiner, Germany), using a broth microdilution protocol modified from the Clinical and Laboratory Standards Institute M-27A methods (National Committee for Clinical Laboratory Standards 2002). Minimal inhibition concentrations (MIC) and the growth under different concentrations of antifungal agents were measured. Representative aliquot of well-mixed 2 μl of samples from each well was spotted on YEPD media to monitor cells recovery after the drug treatment. All experiments were done in triplicate. Drug interactions from FLC and AmB with FPZ were calculated using a fractional inhibitory concentration index (FICI; Zhang et al. 2007). The FICI was interpreted as follows: a synergistic effect is indicated by FICI ≤ 0.5, an additive or indifferent effect when FICI > 0.5 and <4, and an antagonistic effect when FICI > 4.
Relative quantification of differentially expressed genes by real-time PCR
All primer sequences are listed in Table S1. RNA isolation, complementary DNA synthesis, and polymerase chain reaction (PCR) amplification was carried out as previously described (Xu et al. 2006). Real-time PCRs of triplicate samples were performed using the LightCycler System (Roche Diagnostics). The gene expression level relative to the calibrator was expressed as 2-ΔΔCT.
Ergosterol measurement
Ergosterol extraction was based on a previously described procedure with modification (Breivik and Owades 1957). A single C. albicans or S. cerevisiae colony from an overnight SD agar (0.67 % yeast nitrogen base, 2 % glucose, and 2 % agar) culture was used to inoculate 50 ml of SD Broth (0.67 % yeast nitrogen base and 2 % glucose) for 16 h at 35 °C. Cells were harvested by centrifugation at 5,000 rpm for 5 min. The weight of the cell pellet was determined after lyophilization. 3 ml of freshly prepared 25 % alcoholic potassium hydroxide solution was added to each pellet and vortex for 1 min. Cell suspensions were transferred to sterile screw-cap tubes and were incubated in a water bath at 85 °C for 1 h. After the tubes were cooled, ergosterol was extracted by the addition of a mixture containing 1 ml of sterile distilled water and 3 ml of heptane and vigorously vortex for 3 min. The heptane layer was carefully transferred to a clean screw-cap tube and stored at −20 °C. A Shimadzu 20A HPLC was used to analyze ergosterol with a mobile phase of 1 ml methanol/min. Ergosterol levels were calculated as the peak area compared to cell weight (abs/mg; Breivik and Owades 1957).
Filipin-binding assay
Single colonies of WT and Δcdr1 strains from overnight SD agar plates were used to inoculate 50 ml SD Broth cultures. The liquid cultures were grown for 16 h at 35 °C, and the cells were harvested by centrifugation at 5,000 rpm for 5 min. Cells were then rinsed three times with autoclaved phosphate buffered saline (PBS) buffer. After that, the cells were suspended in PBS buffer to make the final cell concentration of 1 × 107 cells/ml. Filipin was then added at a concentration of 10 μg/ml and incubated for 30 min at 35°C 200 rpm. Finally, cells were harvested and rinsed twice with PBS buffer, and the fluorescence was measured (exCitation at 340 nm and emission at 480 nm).
Results
Elevated expression of ABC transporters contributes to azole resistance and AmB susceptibility
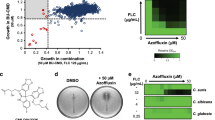
KTC and FLC were used to monitor azole susceptibility of the C. albicans and S. cerevisiae strains (Table 1). In isogenic laboratory C. albicans, WT strain was resistant to azoles due to the over-expression of drug efflux pumps (Fig. S1A), while Δcdr1 and Δcdr2 mutants were sensitive to azoles compared to WT strain (Fig. 1a). No growth of Δcdr1 strain occurred on KTC or FLC plates. Only at a high cell density, Δcdr2 strain was able to grow on azole plates (at 1 × 107 cells/ml on KTC plate and 1 × 106 cells/ml on FLC plate, respectively). Interestingly, in contrast to the susceptibility to azoles, the Δcdr1 mutant was resistant to AmB compared to WT strain. Specifically, growth of Δcdr1 strain on AmB agar occurred after 104-fold dilution (at 1 × 103 cells/ml), while the WT strain could only grow at a higher cell density (1 × 107 cells/ml; Fig. 1a). A growth increase of the Δcdr2 mutant was also observed on AmB plate compared to WT strain as its growth occurred after a 103-fold dilution (at 1 × 104 cells/ml; Fig. 1a).

Over-expression of drug efflux pumps responsible for azole resistance but AmB susceptibility. Spots assay: The strains were grown in YEPD overnight at 35 °C and then 2 μl ten-fold serial dilutions were spotted onto YEPD plates with solvent control or containing one of the drugs (a: 0.5 μg/ml KTC, 1 μg/ml FLC, 7 μg/ml AmB; b: 0.5 μg/ml KTC, 2 μg/ml FLC, 20 μg/ml AmB; c: 4 μg/ml KTC, 8 μg/ml FLC, 7 μg/ml AmB), and plates were incubated at 35 °C for 24 h for C. albicans or 48 h for S. cerevisiae
To exclude the effect of unknown factors, isogenic strains of S. cerevisiae AD/CaCDR1 and AD/CaCDR2 (Table 1; Lamping et al. 2007), which contain over-expressed CaCDR1 and CaCDR2 genes from C. albicans, respectively, were used to investigate the relationship between drug efflux pumps and AmB susceptibility. Strains AD/CaCDR1 and AD/CaCDR2 exhibited the increase of resistance to KTC and FLC due to the over-expression of ABC transporters, while no growth occurred for parent strain AD1-8u- on azole plates (Fig. 1b). However, AD/CaCDR1 and AD/CaCDR2 strains were sensitive to AmB (no growth occurred on the AmB plate), but strain AD1-8u- can still grow at a low cell density (1 × 104 cells/ml; Fig. 1b). This indicates that the elevated expression of drug efflux pumps (both CaCDR1 and CaCDR2) contributes to the increase of susceptibility to AmB.

Interactions of FLC and AmB with drug efflux pumps inducer FPZ. C. albicans SC5314 was treated by twofold dilution drugs: FLC (from 16 to 0.031 μg/ml), AmB (from 4 to 0.0078 μg/ml) and FPZ (from 100 to 0.20 μg/ml) alone or the diluted FLC (from 16 to 0.031 μg/ml) and AmB (from 4 to 0.0078 μg/ml) supplemented with different concentration of FPZ (12, 6 and 3 μg/ml) and DMSO as control. The experiment was performed in the 96-well plate in RPMI 1640 medium. After the incubation at 35 °C overnight, plates were carefully shaken, a representative aliquot of 2 μl of each well was spotted on fresh YEPD recovery plates. Recovery plates were incubated at 35 °C overnight. a: Antagonistic interactions of FLC and FPZ; b: Synergistic interactions of AmB and FPZ
The counterintuitive phenomenon led us to test whether AmB can kill the clinical azole-resistant isolates with over-expressed drug efflux pumps. Isogenic clinical isolates (#1, #4, and #17) from an HIV-infected patient (White 1997) were employed. In agreement with previous results, azole resistance gradually increased (Fig. 1c) in strains #1, #4, and #17, which is consistent with the elevated expression of drug efflux pumps (Fig. S1B; White 1997). The growth of strain #1 only occurred at a high cell density on azole plates (at 1 × 107 cells/ml on KTC and FLC plates; Fig. 1c). Similarly, for strain #4, growth only occurred at 1 × 106 cells/ml on KTC plate and at 1 × 104 cells/ml on FLC plate. Strain #17 was the most azole-resistant one as its growth occurred at 1 × 104 cells/ml on KTC plate and at 1 × 103 cells/ml on FLC plate (Fig. 1c). In contrast, the serial clinical strains showed decreasing resistance to AmB (Fig. 1c). Strain #17 could only grow at a higher cell density (1 × 106 cells/ml) on AmB plate, while strain #1 grew at 1 × 104 cells/ml and #4 grew at 1 × 105 cells/ml (Fig. 1c).
AmB synergized with drug efflux pumps inducer FPZ
To further decipher whether the susceptibility of AmB is directly related to the expression of efflux pumps, we tested the drug sensitivity of AmB or FLC in combination with the ABC transporter inducer fluphenazine (FPZ; Micheli et al. 2002) on C. albicans SC5314. The sensitivity to FLC was significantly reduced when supplemented with FPZ (at 3, 6, and 12 μg/ml) with an FICI > 4, indicating the antagonistic effect (Fig. 2a). Yet AmB could synergize with FPZ (at 3, 6, and 12 μg/ml; Fig. 2b), and the FICI values were 0.31, 0.13, and 0.25, respectively.
Over-expression of efflux pumps is coupled with elevated levels of ergosterol
AmB binds to ergosterol to form an AmB/sterol complex, which forms cell pores and changes the permeability of the plasma membrane leading to cell death (Baginski et al. 2005; Bossche et al. 1987; Brajtburg and Bolard 1996; Brajtburg et al. 1990; Schultz et al. 1984). In line with a previous report, the disruption of ergosterol biosynthesis (such as CaERG3 and CaERG11) reduced the sensitivity to AmB (Fig. 3a; Sanglard et al. 2003). The CaERG11 and CaERG3 null strain increased the MICs of AmB over 20-fold compared to the WT strain (Fig. 3a). The susceptibility to AmB from azole-resistant strains is possibly due to the over-expression of drug efflux pumps coupled with an increase of ergosterol content. To test this hypothesis, ergosterol levels from strains with different drug efflux pump expression levels were measured. Ergosterol levels from the Δcdr1 and Δcdr2 mutants were significantly decreased compared to WT strain (P < 0.01), consistent with the increased resistance to AmB (Figs. 3b and 1a). Over-expression of CaCDR1 and CaCDR2 in S. cerevisiae significantly increased the ergosterol levels (P < 0.01) compared with their parent strain AD1-8u- (Fig. 3b), which directly confirmed the relationship between ergosterol and drug efflux pumps. Due to the over-expression of CaCDR1 and CaCDR2 (Fig. S1B), clinical strain #17, which was most sensitive to AmB (Fig. 1c), had significantly increased ergosterol level (P < 0.01) compared to the strain #1 and #4 (Fig. 3b). We also measured CaERG11 expression level in this serial clinical azole-resistant strains. CaERG11 expression gradually increased in #1, #4, and #17, consistent with the increase of ergosterol content (Fig. 3b and c). This demonstrates that over-expression of drug efflux pumps was coupled with the increase of the ergosterol biosynthesis which led to azole resistance but AmB susceptibility. In order to prove that elevated ergosterol can bind more polyene antibiotics, we use filipin, the fluorescent analog of AmB, to measure their bindings on the WT strain and Δcdr1 mutant. As shown in Fig. 3d, Δcdr1 mutant significantly reduced filipin binding compared to the WT strain (P < 0.05; Fig. 3d). This is in consistent with the reduction of ergosterol due to that knock out of drug efflux pumps (Fig. 3b), indicating that elevated ergosterol increases polyene antibiotic binding and makes azole-resistant strains sensitive to AmB.

Ergosterol biosynthesis and content related to AmB susceptibility. a: The AmB and FLC MICs from the ergosterol biosynthesis (CaERG3 or CaERG11) disruption strains; b: The ergosterol levels of different strains. The bar indicates the fold change of ergosterol content compared to WT in laboratory strains, the drug efflux pumps heterologous expression S. cerevisiae strains compared to the parent strain AD1-8u-, and the drug efflux pumps over-expressed clinical isolates (#4 and #17) compared to #1. c: Quantitative real-time PCR analysis of CaERG11 gene expression in clinical isolates. Mean values from three independent experiments are shown. Error bars indicate standard deviation. d: The percentage of filipin binding from WT and Δcdr1 strains. Mean values from three independent experiments are shown. Error bars indicate standard deviation
Discussion
Here, we report that azole-resistant strains are sensitive to AmB. Our results reveal a possible mechanism underlying the reversed drug susceptibility, which was directly associated with the major ABC transporters (CaCDR1 and CaCDR2), especially in clinical isolates (Fig. 1). We also prove that the different susceptibilities are correlated with ergosterol level resulting from an increase of ergosterol biosynthesis (Fig. 3). The reverse susceptibility might shed light on the evolution of C. albicans drug resistance resulting from azole selectivity. Azole drugs cause an accumulation of toxic sterols in cells by targeting CaERG11. In response, fungal pathogens can use significant amounts of cellular resources to over-express drug efflux pumps and up-regulate ergosterol biosynthesis genes (Fig. 1 and Fig. 3c) in order to decrease the azole damage. However, the increase of the ergosterol content (Fig. 3b) may lead to susceptibility to polyenes (Fig. 1).
The localization of CaCDR1 within the cell membrane is proved to be associated with ergosterol and sphingolipid content when it is heterologously expressed in S. cerevisiae (Mukhopadhyay et al. 2004; Pasrija et al. 2005, 2008). Depletion of ergosterol and sphingolipid biosynthesis genes altered the localization of CaCdr1p resulting in reduced efflux of Rhodamine 6G, a fluorescent substrate of ABC transporters. This suggests that CaCdr1p has lost efflux function due to a lack of ergosterol and sphingolipid which are the main components of lipid rafts (Pasrija et al. 2008). The over-expressed efflux pumps may need more ergosterol and sphingolipid for CaCdr1p localization. Sanglard et al. analyzed the gene expression profiles of azole-resistant C. albicans isolates treated with azole drugs and showed CaCDR-specific up-regulation of CaERG3 and CaERG6 which are involved in ergosterol biosynthesis (Karababa et al. 2004). Guan et al. (2010) observed that loss of pdr5p function due to mutations causes azole susceptibility and AmB resistance. In our study, we directly demonstrate the relationship between ergosterol and efflux pumps in C. albicans as well as in S. cerevisiae. The drug-resistant pump over-expression up-regulates the ergosterol biosynthesis genes (such as CaERG11 in clinical strains, Fig. 3c; Karababa et al. 2004), then causes an increase in ergosterol levels (Fig. 3b) which may help localize the efflux pumps and perform the efflux function (Pasrija et al. 2008). This series of changes rendered C. albicans vulnerable to AmB (Fig. 1; Hsuchen and Feingold 1973). Our results from CaCDR2 suggest that both CaCdr2p and CaCdr1p require ergosterol for their localization and function. Other drug efflux pump activities might be involved in AmB susceptibility. For example, ABC transporters are general phospholipid translocators (Dogra et al. 1999; Prasad and Panwar 2004) which can cause the asymmetric distribution of phospholipids between the two monolayers of the plasma membrane. We propose that the asymmetric distribution of phospholipids may expose ergosterol, which can increase the affinity of AmB and ergosterol.
Another observation is that the strain #17, which has the highest efflux pump expression levels (Fig. S1B), is most sensitive to AmB (Fig. 1c). This is accompanied by an increase in CaERG11 expression (Fig. 3c) and ergosterol levels (Fig. 3b). This result provides clear evidence for AmB reverse action in the clinical strains, providing an important consideration in treating azole-resistant strains. Moreover, we used the drug efflux pump inducer FPZ to stimulate the development of ABC transporters and to accelerate azole resistance as indicated by the antagonistic activity of the FLC and FPZ combination (Fig. 2a). The synergistic activity from the combination of FPZ and AmB (Fig. 2b) indicates that we may even first induce C. albicans drug resistance and then kill the pathogens with AmB.
Some researchers have observed that a combination of azoles (such as posaconazole and itraconazole) and AmB can decrease the burden of fungal infection (Larsen et al. 2004; Lewis et al. 2002; Rodriguez et al. 2008). Yet others have observed no difference using a combination of KTC or FLC with AmB (Lewis et al. 1998; Martin et al. 1994; Schaffner and Frick 1985). Our results provide a novel approach of using AmB alone in dealing with azole-resistant fungal pathogen instead of increasing the dosage of azoles.
References
Baginski M, Sternal K, Czub J, Borowski E (2005) Molecular modelling of membrane activity of amphotericin B, a polyene macrolide antifungal antibiotic. Acta Biochim Pol 52:1–4
Ben-Yaacov R, Knoller S, Caldwell GA, Becker JM, Koltin Y (1994) Candida albicans gene encoding resistance to benomyl and methotrexate is a multidrug resistance gene. Antimicrob Agents Chemother 38(4):648–652
Bossche HV, Willemsens G, Marichal P (1987) Anti-Candida drugs—the biochemical basis for their activity. Crit Rev Microbiol 15(1):57–72
Brajtburg J, Bolard J (1996) Carrier effects on biological activity of amphotericin B. Clin Microbiol Rev 9(4):512–531
Brajtburg J, Powderly WG, Kobayashi GS, Medoff G (1990) Amphotericin B: current understanding of mechanisms of action. Antimicrob Agents Chemother 34(2):183–188
Breivik ON, Owades JL (1957) Spectrophotometric semimicrodetermination of ergosterol in yeast. J Agric Food Chem 5:360–363
Decottignies A, Grant AM, Nichols JW, de Wet H, McIntosh DB, Goffeau A (1998) ATPase and multidrug transport activities of the overexpressed yeast ABC protein Yor1p. J Biol Chem 273(20):12612–12622
Dogra S, Krishnamurthy S, Gupta V, Dixit BL, Gupta CM, Sanglard D, Prasad R (1999) Asymmetric distribution of phosphatidylethanolamine in C. albicans: possible mediation by CDR1, a multidrug transporter belonging to ATP binding cassette (ABC) superfamily. Yeast 15:111–121
Fonzi WA, Irwin MY (1993) Isogenic strain construction and gene mapping in Candida albicans. Genetics 134(3):717–728
Gallagher PJ, Bennett DE, Henman MC, Russell RJ, Flint SR, Shanley DB, Coleman DC (1992) Reduced azole susceptibility of oral isolates of Candida albicans from HIV-positive patients and a derivative exhibiting colony morphology variation. J Gen Microbiol 138(9):1901–1911
Gillum A, Tsay EH, Kirsch D (1984) Isolation of the Candida albicans gene for orotidine-5rsch D (1984) Isolation of the ing colony mo of S. cerevisiae ura3 and E. coli pyrF mutations. Mol Gen Genet 198(1):179–182. doi:10.1007/bf00328721
Goffeau A (2008) Drug resistance: the fight against fungi. Nature 452(7187):541–542
Guan W, Jiang H, Guo X, Mancera E, Xu L, Li Y, Steinmetz L, Li Y, Gu Z (2010) Antagonistic changes in sensitivity to antifungal drugs by mutations of an important ABC transporter gene in a fungal pathogen. PLoS ONE 5(6):e11309
Hsuchen C-C, Feingold DS (1973) Selective membrane toxicity of the polyene antibiotics: studies on lecithin membrane models (liposomes). Antimicrob Agents Chemother 4(3):309–315
Karababa M, Coste AT, Rognon B, Bille J, Sanglard D (2004) Comparison of gene expression profiles of Candida albicans azole-resistant clinical isolates and laboratory strains exposed to drugs inducing multidrug transporters. Antimicrob Agents Chemother 48(8):3064–3079
Lamping E, Monk BC, Niimi K, Holmes AR, Tsao S, Tanabe K, Niimi M, Uehara Y, Cannon RD (2007) Characterization of three classes of membrane proteins involved in fungal azole resistance by functional hyperexpression in Saccharomyces cerevisiae. Eukaryot Cell 6(7):1150–1165
Larsen RA, Bauer M, Thomas AM, Graybill JR (2004) Amphotericin B and fluconazole, a potent combination therapy for Cryptococcal Meningitis. Antimicrob Agents Chemother 48(3):985–991
Lewis RE, Lund BC, Klepser ME, Ernst EJ, Pfaller MA (1998) Assessment of antifungal activities of fluconazole and amphotericin B administered alone and in combination against Candida albicans by using a dynamic in vitro mycotic infection model. Antimicrob Agents Chemother 42(6):1382–1386
Lewis RE, Prince RA, Chi J, Kontoyiannis DP (2002) Itraconazole preexposure attenuates the efficacy of subsequent amphotericin B therapy in a murine model of acute invasive pulmonary aspergillosis. Antimicrob Agents Chemother 46(10):3208–3214
Martin E, Maier F, Bhakdi S (1994) Antagonistic effects of fluconazole and 5-fluorocytosine on candidacidal action of amphotericin B in human serum. Antimicrob Agents Chemother 38(6):1331–1338
Micheli MD, Bille J, Schueller C, Sanglard D (2002) A common drug-responsive element mediates the upregulation of the Candida albicans ABC transporters CDR1 and CDR2, two genes involved in antifungal drug resistance. Mol Microbiol 43(5):1197–1214. doi:10.1046/j.1365-2958.2002.02814.x
Mukhopadhyay K, Kohli A, Prasad R (2002) Drug susceptibilities of yeast cells are affected by membrane lipid composition. Antimicrob Agents Chemother 46(12):3695–3705
Mukhopadhyay K, Prasad T, Saini P, Pucadyil TJ, Chattopadhyay A, Prasad R (2004) Membrane sphingolipid–ergosterol interactions are important determinants of multidrug resistance in Candida albicans. Antimicrob Agents Chemother 48(5):1778–1787
National Committee for Clinical Laboratory Standards (2002) Reference method for broth dilution antifungal susceptibility testing of yeasts approved standard, 2nd edn M27-A2. Approved Standard-2nd edn, Pennsylvania, USA
Pasrija R, Prasad T, Prasad R (2005) Membrane raft lipid constituents affect drug susceptibilities of Candida albicans. Biochem Soc Trans 33:1219–1223
Pasrija R, Panwar SL, Prasad R (2008) Multidrug transporters CaCdr1p and CaMdr1p of Candida albicans display different lipid specificities: both ergosterol and sphingolipids are essential for targeting of CaCdr1p to membrane rafts. Antimicrob Agents Chemother 52(2):694–704
Prasad R, Panwar S (2004) Physiological functions of multidrug transporters in yeast. Curr Sci 86(1):62–73
Prasad R, Wergifosse DP, Goffeau A, Balzi E (1995) Molecular cloning and characterization of a novel gene of Candida albicans, CDR1, conferring multiple resistance to drugs and antifungals. Curr Genet 27(4):320–329
Redding S, Smith J, Farinacci G, Rinaldi M, Fothergill A, Rhine-Chalberg J, Pfaller M (1994) Resistance of Candida albicans to fluconazole during treatment of oropharyngeal Candidiasis in a patient with AIDS: documentation by in vitro susceptibility testing and DNA subtype analysis. Clin Infect Dis 18(2):240–242
Rodriguez MM, Serena C, Marine M, Pastor FJ, Guarro J (2008) Posaconazole combined with amphotericin B, an effective therapy for a murine-disseminated infection caused by Rhizopus oryzae. Antimicrob Agents Chemother 52:3786–3788
Sanglard D, Ischer F, Monod M, Bille J (1996) Susceptibilities of Candida albicans multidrug transporter mutants to various antifungal agents and other metabolic inhibitors. Antimicrob Agents Chemother 40(10):2300–2305
Sanglard D, Ischer F, Monod M, Bille J (1997) Cloning of Candida albicans genes conferring resistance to azole antifungal agents: characterization of CDR2, a new multidrug ABC transporter gene. Microbiology 143(2):405–416
Sanglard D, Ischer F, Parkinson T, Falconer D, Bille J (2003) Candida albicans mutations in the ergosterol biosynthetic pathway and resistance to several antifungal agents. Antimicrob Agents Chemother 47(8):2404–2412. doi:10.1128/aac.47.8.2404-2412.2003
Schaffner A, Frick PG (1985) The effect of ketoconazole on amphotericin B in a model of disseminated aspergillosis. J Infect Dis 151(5):902–910
Schultz SG, Thompson SM, Hudson R, Thomas SR, Suzuki Y (1984) Electrophysiology of Necturus urinary bladder: II. Time-dependent current–voltage relations of the basolateral membranes. J Membrane Biol 79(257–269):257
White TC (1997) Increased mRNA levels of ERG16, CDR, and MDR1 correlate with increases in azole resistance in Candida albicans isolates from a patient infected with human immunodeficiency virus. Antimicrob Agents Chemother 41(7):1482–1487
White TC, Marr KA, Bowden RA (1998) Clinical, cellular, and molecular factors that contribute to antifungal drug resistance. Clin Microbiol Rev 11(2):382–402
Xu Z, Zhang LX, Zhang JD, Cao YB, Yu YY, Wang DJ, Ying K, Chen WS, Jiang YY (2006) cDNA microarray analysis of differential gene expression and regulation in clinically drug-resistant isolates of Candida albicans from bone marrow transplanted patients. Int J Med Microbiol 296(6):421–434
Zhang L, Yan K, Zhang Y, Huang R, Bian J, Zheng C, Sun H, Chen Z, Sun N, An R, Min F, Zhao W, Zhuo Y, You J, Song Y, Yu Z, Liu Z, Yang K, Gao H, Dai H, Zhang X, Wang J, Fu C, Pei G, Liu J, Zhang S, Goodfellow M, Jiang Y, Kuai J, Zhou G, Chen X (2007) High-throughput synergy screening identifies microbial metabolites as combination agents for the treatment of fungal infections. Proc Natl Acad Sci U S A 104(11):4606–4611
Acknowledgment
This work was supported in part by grants from the Ministry of Science and Technology of China (2013CB734001) and National Natural Science Foundation of China (30873129, 30911120484, and 81011120046). We thank Joachim Morschhäuser, Dominique Sanglard, Theodore C. White, and Richard D. Cannon for kindly providing fungal isolates. We also would like to thank Dongmei Li, John Wallace, Yongquan Li, Haoping Liu, Pinghui Gao, Yuanying Jiang, Zhenglong Gu, Chris Vavricka, and Elizabeth Ashforth for critical reading and helpful discussions. L.Z. is an Awardee for National Distinguished Young Scholar Program in China.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(PDF 78 kb)
Rights and permissions
About this article
Cite this article
Ren, B., Dai, HQ., Pei, G. et al. ABC transporters coupled with the elevated ergosterol contents contribute to the azole resistance and amphotericin B susceptibility. Appl Microbiol Biotechnol 98, 2609–2616 (2014). https://doi.org/10.1007/s00253-013-5425-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-013-5425-5