
Abstract
In the course of our screening program for anti-Mycobacterium bovis bacillus Calmette-Guérin (BCG) and anti-Mycobacterium tuberculosis H37Rv (MTB H37Rv) agents from our marine natural product library, a newly isolated actinomycete strain, designated as MS449, was picked out for further investigation. The strain MS449, isolated from a sediment sample collected from South China Sea, produced actinomycin X2 and actinomycin D in substantial quantities, which showed strong inhibition of BCG and MTB H37Rv. The structures of actinomycins were elucidated by nuclear magnetic resonance and mass spectrometric analysis. The strain MS449 was taxonomically characterized on the basis of morphological and phenotypic characteristics, genotypic data, and phylogenetic analysis. The 16S rRNA gene sequence of the strain was determined and a database search indicated that the strain was closely associated with the type strain of Streptomyces avermitilis (99.7 % 16S rRNA gene similarity). S. avermitilis has not been previously reported to produce actinomycins. The marine-derived strain of Streptomyces sp. MS449 produced notably higher quantities of actinomycin X2 (1.92 mg/ml) and actinomycin D (1.77 mg/ml) than previously reported actinomycins producing strains. Thus, MS449 was considered of great potential as a new industrial producing strain of actinomycin X2 and actinomycin D.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Tuberculosis is a contagious disease, caused by Mycobacterium tuberculosis (MTB). MTB infection results in more than two million deaths per year and it is considered the leading cause of mortality in people co-infected with HIV/AIDS (World Health Organization 1999). About one third of the world’s population has been infected with MTB and new infections occur at a rate of one individual per second (Dye et al. 1999). Tuberculosis is becoming a serious threat for disease control and its deadly synergy with HIV/AIDS and a surge in drug-resistant strains make it a primary objective for our team in recent years to discover entirely novel agents that can markedly shorten the time to cure tuberculosis and simplify the treatment.
To date, a lot of work has been done in our lab to accelerate the lead discovery process (Song et al. 2010; Yu et al. 2010; You et al. 2010) and to improve the yields of some bioactive metabolites by biodiversity and taxonomy-guided natural product library construction (Ashforth et al. 2010), high throughput screening (Liu et al. 2010), and genetic engineering project (Zhang et al. 2005; Zhuo et al. 2010). Also, these remarkable efforts facilitated our screening process for anti-Mycobacterium bovis bacillus Calmette-Guérin (BCG) and anti-M. tuberculosis H37Rv (MTB H37Rv) compounds and enhanced the efficiency of the discovery of bioactive agents. The construction of a large marine natural product library (MNPL) (Bian et al. 2008) afforded a strong support for all screening work based on different high throughput screening models, such as synergic anti-fungal screening model (Zhang et al. 2007), anti-methicillin-resistant Staphylococcus aureus, anti-BCG, and anti-fungal screening models, and some other antimicrobial or antitumor screening models at the cellular and molecular levels.
We have developed a convenient high throughput screening model to identify inhibitors in an aerobic, logarithmic growth screen of BCG (Cowley and Av-Gay 2001). The BCG used here was transformed with green fluorescent protein (GFP) constitutive expression plasmid Puv3583 with direct readout of fluorescence as a measure of bacterial growth. In our screening for anti-BCG compounds from our MNPL, a number of positive strains were picked out and their secondary metabolites were analyzed by HPLC-diode array detector. These organisms proved to be a rich source of bioactive compounds. Strain MS449, which was isolated from a sediment sample collected from South China Sea, gained our special interest because of the potential activity against BCG and diversity of metabolites in its fermentation broth. The 16S rRNA gene sequence from the strain was determined and proved to have high identity (99.7 %) with Streptomyces avermitilis. As the type S. avermitilis strain which produces avermectins was available, we compared its features with MS449 and analyzed the HPLC-UV–Vis characteristic of its secondary metabolites and found that it evidently differed from the strain MS449 in morphological and metabolic characteristics. Therefore, MS449 was picked out for further investigation and resulted in three actinomycins with potent bioactivities.
Actinomycins are a family of chromopeptide lactone antibiotics, among which actinomycin D has been studied extensively and widely used clinically for the treatment of several types of malignant tumors, notably Wilms’ tumor (Green 1997) and childhood rhabdomyosarcoma (Womer 1997). Since the discovery of actinomycin D, it has become an important tool in clinical, molecular, and cell biology (Kurosawa et al. 2006) because of its unique structure and biological properties by intercalating into duplex DNA, which results in inhibition of DNA-dependent RNA polymerase and thus protein synthesis (Wadkins et al. 1998). It was reported to inhibit the coxsackievirus B3 (Saijets et al. 2003) and was proposed for the treatment of AIDS because of its effectiveness in inhibiting the minus-strand transfer process in HIV-1 (Rill and Hecker 1996). By contrast, the anti-tuberculosis activity of actinomycins was not paid much attention, and little research has been reported. The literature review indicated that subsequent research mainly focused on the structure modification and biofunction of actinomycins with no more attention to actinomycin X2 and X0β especially in the anti-tuberculosis aspect (Sun and Yang 2011). The strain with high production of actinomycin X2, actinomycin D, and relatively less actinomycin X0β may be valuable for researchers’ investigation.
In this paper, we describe the marine-derived Streptomyces strain MS449, which exhibits potent activities against BCG and MTB H37Rv and is a stable high yield producer of actinomycin X2, actinomycin D, and actinomycin X0β even under non-optimized culture conditions. The strain MS449 was identified as S. avermitilis based on taxonomic experiments and it is a novel source for actinomycins.
Materials and methods
Microbial strains
An actinomycete strain MS449, used as the producing strain, was isolated from a sediment sample collected from South China Sea (longitude 110:22:47.058 and latitude 17:33:35.661) and grown on an GT medium agar slant consisting of soluble starch 2.0 %, l-asparagine 0.05 %, KNO3 0.1 %, K2HPO4·H2O 0.05 %, NaCl 0.05 %, and MgSO4⋅7H2O 0.05 % (pH 7.5) at 28 °C. MS449 was deposited at the China General Microbiological Culture Collection Center (CGMCC) with the accession number CGMCC No. 5452.
Mycobacterium phlei (ATCC 11758), used as an indicator organism during the isolation process, was grown on an agar plate consisting of peptone 1.0 %, malt extract 0.5 %, yeast extract 0.5 %, casein acid hydrolysate 0.5 %, beef extract 0.2 %, glycerol 0.2 %, Tween-80 0.005 %, MgSO4⋅7H2O 0.1 %, and agar 1.2 %, pH 7.2 at 37 °C. M. bovis BCG (Pasteur 1173P2) and Mycobacterium tuberculosis H37Rv (ATCC27294) were used here for bioactivity screening and assays.
Enterococcus faecalis HH22 (Shiojima et al. 1997), carrying bifunctional modifying enzyme genes in plasmid PBE10, was clinically isolated and resistant to gentamycin, penicillin, erythromycin, and tetracycline. Proteus mirabilis PM631 SE (Neuwirth et al. 2001) was a clinical isolate producing TEM-3 β-lactamase. E. faecalis HH22 and PM631 SE were preserved in the Institute of Medicinal Biotechnology, Chinese Academy of Medical Sciences & Peking Union Medical College. E. faecalis HH22 and PM631 SE, together with Pseudomonas aeruginosa (ATCC 27853), Escherichia coli (ATCC 25922), S. aureus (ATCC 29213), Mycobacterium smegmatis (ATCC 700044), and M. phlei (ATCC 11758), were used to test antimicrobial properties of the fermentation broth using paper disk-agar diffusion assay (Raahave 1974; Fosto et al. 2008).
Cultural and morphological properties of strain MS449
A set of cultural and phenotypic characteristics was examined using media and the International Streptomyces Project procedures recommended by Shirling and Gottlieb (1966). Mature aerial mycelium and substrate mycelium pigmentation were recorded on GT agar media following incubation at 28 °C for 28 days. Morphological features of spores and mycelia were observed by scanning electron microscopy (Quanta 200) after growth on GT, desiccation, and coating the cells with gold.
16S rRNA gene sequence and phylogenetic analysis
Extraction of MS449 genomic DNA was done as described by the TINAamp Bacteria DNA Kit. Universal primers (27f: 5′-GAGAGTTTGATCCTGGCTCAG-3′; 1492r: 5′-CTACGGCTACCTTGTTACGA-3′) were used to amplify the 16S rDNA. PCR amplification [25 μl final volume—0.4 μl 20 μM of each primer, 2.5 μl 10× buffer (TaKaRa, Dalian, China), 2.5 μl 2.5 nM dNTP (TaKaRa), 2 U rTap polymerase (TaKaRa), and 1 μl DNA template] of the 16S rDNA was performed on TaKaRa PCR Thermal Cycler with the initial denaturation at 94 °C for 5 min, 30 cycles of denaturation (94 °C, 1 min), annealing (55 °C, 1 min), and elongation (72 °C, 1 min 15 s), and a final elongation at 72 °C for 10 min. Multiple alignments with sequences of most closely related Streptomyces and calculations of levels of sequence similarity were carried out using CLUSTAL W (Thompson et al. 1994). The phylogenetic tree was constructed using the neighbor-joining method (Saitou and Nei 1987) by MEGA 4.0 (Tamura et al. 2007). The topology of phylogenetic tree was evaluated by bootstrap resampling method with 1,000 replicates (Felsenstein 1985).
Nucleotide sequence accession number
The nucleotide sequence of 16S rRNA gene reported in this article was assigned to the GenBank accession number JN104388.
Production of actinomycins by MS449 in different producing media
A stock culture of the strain was grown and maintained on a GT agar slant. The stock culture was transferred into 250-ml Erlenmeyer flasks containing 40 ml of the seed medium with the same components as the agar slant medium. The culture was incubated on a rotary shaker (220 rpm) at 28 °C for 96 h. Four milliliters of the seed culture was transferred into 250-ml Erlenmeyer flasks containing 40 ml of the producing medium. Three kinds of producing media were used: (a) M21 medium consisting of glucose 0.5 %, lactose 4 %, cottonseed protein 3 %, Bacto Peptone 0.5 %, K2HPO4 0.05 %, MgSO4⋅7H2O 0.05 %, and KCl 0.03 % (pH 7.0); (b) 9A medium consisting of yeast extract 1.0 %, saccharose 5.0 %, K2HPO4 1.0 %, NaNO3 0.3 %, KCl 0.05 %, MgSO4 0.05 %, and FeSO4 0.001 % (pH 6.5); and (c) MPG medium consisting of glucose 1.0 %, millet meal 2.0 %, cotton seed gluten meal 2.0 %, and MOPS 2.0 %, pH 7.0. The cultures were incubated at 28 °C for 17 days on a rotary shaker at 220 rpm. After incubation for 5 days, aliquots (2 ml) of the cultures were collected every 48 h and prepared for HPLC analysis as follows: After centrifugation (8,000 rpm, 3 min), the mycelia were extracted twice with acetone to give the organic fraction. The supernatants were added 0.5 ml HP-20 resins, rocked for 2 h on a shaker, and filtrated, and the resins were washed with acetone three times. The acetone fractions from the mycelia and the supernatants were combined and dried to give crude extracts. Each crude extract was then redissolved in 2 ml acetone, filtered through a 0.45-μm membrane filter, and analyzed by RP-HPLC equipped with an Agilent XDB-C8, 5 μm column (4.6 × 150 mm), and 70 % methanol in water as the mobile phase. External standard method was used in the quantitative analysis with actinomycin X2 (Alexis), actinomycin D (Sigma), and actinomycin X0β (Alexis) as standards and 446 nm as the detection wavelength.
Susceptibility test of fermentation broth with MPG medium
P. aeruginosa, E. coli, S. aureus, M. smegmatis, M. phlei, E. faecalis HH22, and PM631 SE were grown for 24 h at 37 °C in LB medium; 1 % (v/v) microorganism cultures were mixed with sterilized LB agar medium to make uniform microorganism suspensions for pouring plates before the medium solidified. Six-millimeter filter paper disks impregnated with testing samples were allowed to dry and then placed on the solidified agar plates and incubated at 37 °C overnight. The area of no growth around the disk was known as a “zone of inhibition”.
Purification and characterization of actinomycins
The fermentation broth of MS449 grown in MPG medium (200 ml) at 28 °C for 15 days was fractionated by centrifugation to give the supernatant and mycelium. The mycelium was soaked with acetone and filtered. The filtrate was diluted to five times volume with the supernatant or water and then the mixture was charged on a column of HP-20 resins, which was washed with distilled water and then eluted with 50, 65, and 80 % acetone, respectively. The active fractions eluted with 65 % acetone were combined and removed acetone in vacuo, and then the water residue was extracted twice with ethyl acetate. The organic fractions were combined, dried (over Na2SO4), and concentrated under reduced pressure. The residue was then resuspended in acetone and gave a pure active constituent (1) using preparative RP-HPLC equipped with an Eclipse XDB-C18, 5 μm column (9.4 × 250 mm, Agilent), and 70 % MeOH in water as the mobile phase. The active fractions eluted with 80 % acetone from HP-20 resins were also separated into pure constituents using RP-HPLC and gave two pure compounds (2 and 3). Antibiotic activity of the culture broth was determined and the bioactive fractions were tracked by conventional paper disk-agar diffusion assay using M. phlei, and the zones of inhibition were measured after 18 h of incubation at 37 °C.
The actinomycins were characterized using spectroscopic analyses. Electrospray ionization mass spectra (ESI-MS) were recorded on a Bruker Esquire3000 plus spectrometer and various nuclear magnetic resonance (NMR) spectra were recorded on a Varian Inova 600 MHz spectrometer and the solvent was acetone-d 6.
Anti-BCG and MTB assay
The anti-BCG and anti-MTB H37Rv assays utilize constitutive GFP expression (pUV3583c-GFP) with direct readout of fluorescence as a measure of bacterial growth (Cowley and Av-Gay 2001). The in vitro activity of compounds against BCG and MTB H37Rv was determined in a 96-well plate as previously described (Wang et al. 2010). BCG and MTB H37Rv were grown at 37 °C to mid-log phase in Middlebrook 7H9 broth supplemented with 10 % OADC enrichment (Becton Dickinson), 0.05 % Tween-80, and 0.2 % glycerol. The cultures were then diluted to a bacterial suspension with OD600 values of 0.025 with culture media. For primary screening, aliquots (80 μl) of the bacterial suspension were added to each well of the clear flat-bottom 96-well microplates, followed by adding 2 μl of sample solutions (1 mg/ml). Isoniazid served as the positive control while DMSO served as the blank control. The plates incubated at 37 °C for 3 days for BCG or 10 days for MTB H37Rv GFP fluorescence were measured with a multi-label plate reader (Perkin-Elmer Envision 2103) using the bottom read mode, with excitation at 485 nm and emission at 535 nm. Hits were defined as compounds that could inhibit more than 90 % of bacterial growth compared to the blank control at a concentration of 64 μg/ml or less.
For MIC determination of the compounds, bacterial suspensions were prepared and dispensed as described above, and compounds were serially diluted twofold from 25 to 0.19 μg/ml in each column. Isoniazid as positive control (800 to 6.25 ng/ml) and DMSO as blank control were also included in each plate. The plate was incubated at 37 °C for an appropriate time, and after incubation, the plate was bottom-read to record the GFP fluorescence. Here, MIC is defined as the minimum concentration of drug that inhibits more than 90 % of bacteria growth reflecting by fluorescence values. Anti-BCG and anti-MTB H37Rv assays were performed in Biosafety level (BSL) 2 and BSL3 settings, respectively.
Results
Characterization and identification of isolated strain MS449
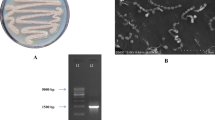
A number of actinomycete strains were isolated from a sediment sample collected from South China Sea and investigated with small-scale fermentation. The strains with their crude extracts were deposited in our MNPL and evaluated for their ability to produce antibiotics by high throughput screening. Strain MS449 showed strong anti-BCG activity and was picked out to be further investigated. The colonial (Fig. 1) and morphological properties (Fig. 2) suggested that MS449 was a member of the genus Streptomyces. Substrate mycelium is yellow in color. Aerial mycelium, where produced, on medium such as GT is white to yellowish white with spinous surfaces. Soluble yellow pigments are produced. Aerial hyphae differentiate into short, straight chains of smooth-surfaced, cylindrical-like spores (0.5–0.7 μm × 1.5–2.0 μm) after 28 days of cultivation on GT plates.

Colony characteristics of MS449 grown on GT agar at 28 °C for 28 days

Scanning electron micrograph of MS449 grown on GT agar at 28 °C for 28 days. Bar, 5 μm
The comparison between the 16S rRNA gene sequence of strain MS449 (1448 nt) and sequence in GenBank indicated that the novel isolate was closely related to the genus Streptomyces and has the highest similarity with S. avermitilis (99.70 %). Phylogenetic analysis based on 16S rRNA gene sequence analysis revealed that strain MS449 formed a distinct phylogenetic cluster with S. avermitilis in the phylogenetic tree (Fig. 3) with a bootstrap value above 95 %.

Neighbor-joining phylogenetic tree of strain MS449 based on 16S rRNA gene sequence generated by Mega4.0. Numbers at nodes indicate levels of bootstrap support (percent) based on a neighbor-joining analysis of 1,000 resampled datasets; only values >50 % are given. NCBI accession numbers are given in parentheses. Bar, 0.01 nucleotide substitutions per site. Also, Kutzneria albida was chosen as the outgroup
Production of actinomycins by MS449 in different producing media
Actinomycin X0β, actinomycin X2, and actinomycin D (1–3) (Fig. 4) were detected in three crude extracts and the three major components were eluted by HPLC at 8.6, 14.7, and 16.9 min, respectively (Fig. 5). The relative proportion of the three identified antibiotics remained consistent in the three media. The highest actinomycin productivity was observed in MPG medium at 15 days (Fig. 6). The yields of actinomycin X0β, actinomycin X2, and actinomycin D from MPG medium were 0.15, 1.92, and 1.77 mg/ml, respectively.

The structures of isolated compounds (1–3)

HPLC analysis of crude extracts of MS449 cultured in MPG medium. Extracts were analyzed using Shimadzu 20A HPLC with an Agilent XDB-C8 column (5 μm, 4.6 × 150 mm), 70 % methanol as the mobile phase, 1.0 ml/min as the flow rate, and 254 nm as the detection wavelength

Time course production of actinomycin X2 (a), actinomycin D (b), and actinomycin X0β (c) by MS449 cultured in MPG (square), M21 (circle), and 9A (triangle) media. Results represent the means ± standard errors of the means (SEM) of three independent experiments
Purification and characterization of actinomycins produced by MS449
Active components were isolated from MPG medium culture broth (200 ml) and three bioactive compounds were obtained (1–3). Their structures were elucidated by UV, 1D NMR, 2D NMR (1H-1H COSY, 1H-13C HSQC, 1H-13C HMBC), and MS analysis as well as by comparison with the previously reported NMR data of actinomycins. All isolated compounds showed typical UV–Vis spectra with maximal absorbance at 241 nm (shoulder) and 446 nm similar to those of known actinomycins (Fig. 5). ESI-MS of isolated compounds revealed molecular ion peaks at m/z 1,271.3 [M+H]+ for 1, 1,269.1 [M+H]+ and 1,291.1 [M+Na]+ for 2, and 1,255.1 [M+H]+ and 1,277.1 [M+Na]+ for 3 (Supplementary materials, Fig. S1), which were identical to those of actinomycin X0β, actinomycin X2, and actinomycin D, respectively. 1H, 13C (Supplementary materials, Table S1) and 2D NMR spectra of isolated compounds in acetone-d 6 showed a great similarity. From 1D, 2D NMR, and by comparison with the previously reported data (Yu and Tseng. 1992; Zhang et al. 2009; Lackner et al. 2000), the structures of 1, 2, and 3 were confirmed to be actinomycin X0β, actinomycin X2, and actinomycin D (Fig. 4).
Susceptibility test of fermentation broth
Susceptibility of fermentation broth of MS449 in MPG medium against a variety of microorganisms was performed by paper disk diffusion assay. The relevant comparisons between the zones of inhibition are shown in Table 1. The fermentation broth showed no activity against P. aeruginosa and PM631 SE. According to the test results, we choose one of the positive microorganisms, M. phlei, which also belongs to the genus Mycobacterium, as the indicator organism (Supplementary materials, Fig. S2) to track the bioactive components during the isolation process.
Anti-BCG and anti-tuberculosis activities of actinomycins
The pure compounds 1–3 all showed activities against BCG with MIC values of 0.25–0.5 μg/ml and MTB H37Rv with MIC values of 8.0, 1.0, and 8.0 μg/ml, respectively. Actinomycin X2 showed the best activity against tuberculosis among these three compounds.
Discussion
MS449 is the first reported naturally occurring strain of S. avermitilis producing a notably high yield of actinomycin X2 and actinomycin D. To the best of our knowledge, approximately 30 species of Streptomyces and Micromonospora were reported capable of producing actinomycins (Kurosawa et al. 2006; Praveen and Tripathi 2009). However, only a few strains were reported to produce relatively large quantities of one major component of actinomycins, such as Streptomyces parvulus (0.152 mg/ml actinomycin D under optimized conditions) (Praveen et al. 2008a), Streptomyces griseoruber (0.21 mg/ml actinomycin D) (Praveen and Tripathi 2009), a mutant strain of Streptomyces sindenensis (0.85 mg/ml actinomycin D) (Praveen et al. 2008b), Streptomyces MITKK-103 (0.11 mg/ml actinomycin X2) (Kurosawa et al. 2006), Streptomyces spp. JAU4234 (0.62 mg/ml actinomycin X2 under optimized conditions) (Xiong et al. 2008), and Streptomyces nasri strain YG62 (0.15 mg/ml actinomycin X2) (EI-Naggar 1998), whereas strain MS449 exhibited a unique ability to produce large quantities of actinomycin X2 (2), actinomycin D (3), and actinomycin X0β (1) simultaneously with a production of 1.92, 1.77, and 0.15 mg/ml, respectively, under non-optimized conditions. Actinomycin X2 production in MPG medium reached a sub-maximum peak after 9 days and increased slowly afterwards. The production of actinomycin D and actinomycin X0β in MPG increased markedly after 11 days. Maximum levels of actinomycin productivity were reached after 15 days (Fig. 6). Optimization of the culture conditions was expected to be done to exploit the full potential for industrial application of this strain.
Actinomycins, which were firstly discovered by Waksman and Tishler (1942), represent a family of chromopeptide antibiotics which differ solely in the peptide portions of the molecules. To date, more than 30 analogues have been described. Actinomycins have become an important tool in molecular and cellular biology. Among actinomycins, actinomycin D has been extensively studied and used clinically. The exciting potentialities of actinomycin D have stimulated a great deal of research into its chemical, physical, and biological characteristics (Hollstein 1974; Henry 1985). Although little research about actinomycin X2 and actinomycin X0β was reported, actinomycin X2 showed better activity against MTB H37Rv than actinomycin D and actinomycin X0β in our research. It was also reported that actinomycin X2 showed higher cytotoxicity toward HL-60 cells than actinomycin D (Kurosawa et al. 2006). It is affirmative that actinomycin X2 will be more and more attractive. As a new source of actinomycin X2, MS449 is considered of great potential for industrial application.
References
Ashforth EJ, Fu CZ, Liu XY, Dai HQ, Song FH, Guo H, Zhang LX (2010) Bioprospecting for antituberculosis leads from microbial metabolites. Nat Prod Rep 27:1709–1719
Bian J, Song FH, Zhang LX (2008) Strategies on the construction of high-quality microbial natural product library—a review. Acta Microbiol Sin 48:1132–1137
Cowley SC, Av-Gay Y (2001) Monitoring promoter activity and protein localization in Mycobacterium spp using green fluorescent protein. Gene 264:225–231
Dye C, Scheele S, Dolin P, Pathania V, Raviglione MC (1999) Consensus statement. Global burden of tuberculosis: estimated incidence, prevalence, and mortality by country. WHO Global Surveillance and Monitoring Project. J Am Med Assoc 282:677–686
EI-Naggar MYM (1998) Synergistic effect of actinomycin X2 produced by Streptomyces nasri YG62 with other antibiotics. Biomed Lett 58:169–173
Felsenstein J (1985) Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39:783–791
Fosto S, Mahmud T, Zabriskie TM, Santosa DA, Proteau PJ (2008) Rearranged and unrearranged angucyclinones from Indonesian Streptomyces spp. J Antibiot 61:449–456
Green DM (1997) Wilm’s tumour. Eur J Cancer 33:409–418
Henry MS (1985) Actinomycin and DNA transcription. Proc Natl Acad Sci 82:5328–5331
Hollstein U (1974) Actinomycin, chemistry and mechanism of action. Chem Rev 74:625–652
Kurosawa K, Buim VP, VanEssendelft JL, Willis LB, Lessard PA, Ghiviriga I, Sambandan TG, Rha CK, Sinskey AJ (2006) Characterization of Streptomyces MITKK-103, a newly isolated actinomycin X2-producer. Appl Microbiol Biotechnol 72:145–154
Lackner H, Bahner I, Shigematsu N, Pannell LK, Mauger AB (2000) Structure of five components of the actinomycin Z complex from Streptomyces fradiae, two of which contain 4-chlorothreonine. J Nat Prod 63:352–356
Liu XY, Ashforth E, Ren B, Song FH, Dai HQ, Liu M, Wang J, Xie Q, Zhang LX (2010) Bioprospecting microbial natural product libraries from the marine environment for drug discovery. J Antibiot 63:415–422
Neuwirth C, Madec S, Siebor E, Pechinot A, Duez JM, Pruneaux M, Fouchereau-Peron M, Kazmierczak A, Labia R (2001) TEM-89β-lactamase produced by a Proteus mirabilis clinical isolate: new complex mutant (CMT 3) with mutations in both TEM-59 (IRT-17) and TEM-3. Antimicrob Agents Chemother 45:3591–3594
Praveen V, Tripathi CKM (2009) Studies on the production of actinomycin-D by Streptomyces griseoruber—a novel source. Lett Appl Microbiol 49:450–455
Praveen V, Tripathi CKM, Bihari V (2008a) Studies on optimum fermentation conditions for actinomycin-D production by two new strains of Streptomyces spp. Med Chem Res 17:114–122
Praveen V, Tripathi CKM, Bihari V, Srivastava SC (2008b) Production of actinomycin-D by the mutant of a new isolate of Streptomyces sindenensis. Braz J Microbiol 39:689–692
Raahave D (1974) Paper disk-agar diffusion assay of penicillin in the presence of streptomycin. Antimicrob Agents Chemother 6:603–605
Rill RL, Hecker KH (1996) Sequence-specific actinomycin-D binding to single-stranded DNA inhibits HIV reverse transcriptase and other polymerases. Biochem 35:3525–3533
Saijets S, Ylipaasto P, Vaarala O, Hovi T, Roivainen M (2003) Enterovirus infection and activation of human umbilical vein endothelial cells. J Med Virol 70:430–439
Saitou N, Nei M (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Shiojima M, Tomita H, Tanimoto K, Fujimoto S, Ike Y (1997) High-Level plasmid-mediated gentamicin resistance and pheromone response of plasmids present in clinical isolates of Enterococcus faecalis. Antimicrob Agents Chemother 41:702–705
Shirling EB, Gottlieb D (1966) Methods for characterization of Streptomyces species. Int J Syst Bacteriol 16:313–340
Song FH, Dai HQ, Tong YJ, Ren B, Chen CX, Sun N, Liu XY, Bian J, Liu M, Gao H, Liu HW, Chen XP, Zhang LX (2010) Trichodermaketones A-D and 7-O-methylkoninginin D from the marine fungus Trichoderma koningii. J Nat Prod 73:806–810
Sun ZY, Yang XP (2011) The research progress of actinomycin. J Capital Normal Univ (Nat Sc Ed) 32:54–57 (in Chinese)
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol 24:1596–1599
Thompson JD, Higgins DG, Gibson TJ (1994) ClustalW-improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22:4673–4680
Wadkins RM, Vladu B, Tung CS (1998) Actinomycin D binds to metastable hairpins in single-stranded DNA. Biochem 37:11915–11923
Waksman SA, Tishler M (1942) The chemical nature of actinomycin, an anti-microbial substance produced by Actinomyces antibioticus. J Biol Chem 142:519–528
Wang JF, Dai HQ, Wei YL, Zhu HJ, Yan YM, Wang YH, Long CL, Zhong HM, Zhang LX (2010) Antituberculosis agents and an inhibitor of the para-aminobenzoic acid biosynthetic pathway from Hydnocarpus anthelminthica seeds. Chem Biodivers 7:2046–2053
Womer RB (1997) Soft tissue sarcomas. Eur J Cancer 33:2230–2234
World Health Organization (1999) Removing obstacles to healthy development. World Health Organization, Geneva
Xiong ZQ, Tu XR, Tu GQ (2008) Optimization of medium composition for actinomycin X2 production by Streptomyces spp JAU4234 using response surface methodology. J Ind Microbiol Biotechnol 35:729–734
You JL, Dai HQ, Chen ZH, Liu GJ, He ZX, Song FH, Yang X, Fu HA, Zhang LX, Chen XP (2010) Trichoderone, a novel cytotoxic cyclopentenone and cholesta-7, 22-diene-3β, 5α, 6β-triol with new activities from the marine-derived fungus Trichoderma sp. J Ind Microbiol Biotechnol 37:245–252
Yu C, Tseng Y (1992) NMR study of the solution conformation of actinomycin D. Eur J Biochem 209:181–187
Yu K, Ren B, Wei JL, Chen CX, Song FH, Dai HQ, Zhang LX (2010) Verrucisidin A and B, two prone-type polyketides isolated from a marine derived fungus, Penicillium aurantiogriseum. Marine Drugs 8:2744–2754
Zhang LX, An R, Wang JP, Sun N, Zhang S, Hu JC, Kuai J (2005) Exploring novel bioactive compounds from marine microbes. Curr Opin Microbiol 8:276–281
Zhang LX, Yan KZ, Zhang Y, Huang R, Bian J, Zheng CS, Sun HX, Sun N, An R, Min FG, Zhang WB, Zhuo Y, You JL, Song YJ, Yu ZY, Liu ZH, Yang HK, Gao H, Dai HQ, Zhang XL, Wang J, Fu CZ, Pei G, Liu JT, Zhang S, Goodfellow M, Jiang YY, Kuai J, Zhou GC, Chen XP (2007) High throughput synergy screening identifies microbial metabolites as combination agents for the treatment of fungal infections. Proc Natl Acad Sci 104:4606–4611
Zhang Z, Peng G, Yang G, Xiao CL, Hao XQ (2009) Isolation, purification, identification of structures and study of bioactivity of anti-TB active component 9005B. Chin J Antibiot 34:399–402
Zhuo Y, Zhang WQ, Chen DF, Gao H, Tao J, Liu M, Gou ZX, Zhou XL, Wang Q, Ye B, Zhang Q, Zhang S, Zhang LX (2010) Reverse biological engineering of hrdB to enhance the production of avermectins in an industrial strain of Streptomyces avermitilis. Proc Natl Acad Sci 107:11250–11254
Acknowledgments
This research was partly supported by the National Natural Science Foundation of China (30973665, 30911120483, 30911120484, 81102356, and 81102369), the CAS Pillar Program (XDA04074000), the Ministry of Science and Technology of China (2007DFB31620, 2011ZX09102-011-11, 2012CB721006), and China Postdoctoral Science Foundation (49th). L. Z. is an Awardee for National Distinguished Young Scholar Program in China.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Caixia Chen, Fuhang Song, and Qian Wang contributed equally to this paper.
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(PDF 72 kb)
Rights and permissions
About this article
Cite this article
Chen, C., Song, F., Wang, Q. et al. A marine-derived Streptomyces sp. MS449 produces high yield of actinomycin X2 and actinomycin D with potent anti-tuberculosis activity. Appl Microbiol Biotechnol 95, 919–927 (2012). https://doi.org/10.1007/s00253-012-4079-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-012-4079-z