
Abstract
Sigma factors of the extracytoplasmic function (ECF) subfamily are important regulators of stress responses in bacteria. This work described the characterization of ECF sigma factor SigT in Streptomyces coelicolor. We found the absence of sigT almost abolished the production of the antibiotics actinorhodin (Act) under nitrogen stress. Under nitrogen-limited conditions, significantly reduced Act production and linked actII-ORF4 transcription with respect to wild type were observed in the sigT-null mutant. Using reporter (xylE) fusion to sigT promoter, we demonstrated that sigT was induced by nitrogen limitation in a SigT-dependent manner. Transcriptional analyses showed that SigT controlled the expression of relA, the ppGpp synthetase gene, and consequently affected the Act production upon nitrogen starvation. Co-transcription analysis revealed that sigT was co-transcribed with rstB (gene upstream of sigT) but not with rstA (gene downstream of sigT). Phenotypic and transcriptional results suggested RstA may modulate the activity of SigT positively.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
During the complex life span, the soil-dwelling bacterial genus Streptomyces is challenged with diverse nutritional and environmental stresses, including high osmolarity, heat and cold shock, pH variation, and nutrient starvation, etc. (Chater 2001). Stresses lead to the reduction or cessation of growth, and also to a major switch in gene expression through the use of alternative sigma factors for altering the recognition specificity and binding strength of RNA polymerase to the target promoters (Ishihama 2000).
Among σ70 family, the extracytoplasmic function (ECF) sigma factors present the largest subfamily. They are involved in responses to a wide range of environmental stresses, such as metal homeostasis, starvation, high temperature, reactive oxygen species, and antibiotics (Helmann 2002; Anthony et al. 2005; Alvarez-Martinez et al. 2006, 2007; Bang et al. 2005). In most cases, genes encoding for ECF sigma factors are co-transcribed with a gene encoding their cognate regulator, an inner membrane anti-sigma factor (Helmann 2002).
The best-understood archetypes of ECF sigma factors include Streptomyces coelicolor SigR, SigE, and SigB, etc. SigE was characterized to control cell envelope stress response (Paget et al. 1999a, b), SigR controls disulphide stress response (Paget et al. 1998), and SigB is responsible for osmotic stress response (Kormanec et al. 2000). Nevertheless, up to date, the ECF sigma factor responding to nutrient starvation has not been identified in S. coelicolor. SigB of S. coelicolor may be a candidate as its structural homologue in Bacillus subtilis could respond to nutrient stress (Hecker and Völker 1998).
A very important aspect of Streptomyces biology is the ability of these bacteria, during stationary phase in liquid culture or at the onset of differentiation on solid medium, to synthesize a wide variety of secondary metabolites, including many antibiotics (Bibb 2005). Nutrition plays an important role in the onset and intensity of secondary metabolism. Control of antibiotics biosynthesis was demonstrated to be a multifunctional process in which limiting nutrients such as nitrogen and phosphate play different roles (Doull and Vining 1990). Two sigma factors in S. coelicolor had been reported to affect the production of the pH-reactive antibiotic actinorhodin (Act; red at pH below 8.5, blue at pH above) and of the red-pigmented tripyrrole antibiotic undecylprodigiosin (Red). One is principal and essential sigma factor HrdB, mutation of hrdB could abolish Act and Red production by reducing the ppGpp pool size (Wang et al. 2010). The other is SigB; the sigB mutant produces dramatically higher levels of Act precociously, while producing very low amounts of Red. This phenomenon results primarily from the absence of catalase B which is critically required for osmoprotection and proper differentiation of S. coelicolor cells (Cho et al. 2001). However, no ECF sigma factor in S. coelicolor has been characterized that could connect nutrient stress with antibiotics production.
In our previous work, we described that both ECF sigma factor SigT and its putative anti-sigma factor RstA negatively regulate morphological differentiation and secondary metabolism in S. coelicolor (Mao et al. 2009). Here, we show that SigT regulates the Act production of S. coelicolor upon nitrogen starvation, and this regulation role might result from the control over the transcription of relA by SigT. Promoter activity and real-time PCR analysis showed sigT gene expression was induced by nitrogen stress in SigT-dependent manner. Our findings suggested that SigT may function as a link between nitrogen stress and antibiotics production. Our results also revealed that the gene rstA encoding putative anti-sigma factor of SigT was not co-transcribed with sigT. The phenotypic and transcriptional analysis of rstA-null mutant coincided well with the proposal that RstA acts as a positive regulator of SigT.
Materials and methods
Bacterial strains and culture conditions
The S. coelicolor strains used in this study are listed in Table 1. Escherichia coli TG1 was used for plasmid construction. E. coli ET12567/pUZ8002 was used for DNA introduction into S. coelicolor by conjugation (MacNeil et al. 1992). E. coli BW25113/pIJ790 and DH5α/BT340 were used in λRED-mediated PCR-targeted mutagenesis of rstB (Gust et al. 2003).
S. coelicolor strains were grown and sporulated on solid R2YE and mannitol soya flour agar medium at 30°C. Transformations were performed as previously described (Kieser et al. 2000). SMMS (a solidified version of supplemented minimal medium, SMM) (Floriano and Bibb 1996; Takano et al. 1992), R5 and R2 (Kieser et al. 2000) were used to assess antibiotic production. RNA was isolated from liquid cultures grown in SMM (Takano et al. 1992; Hobbs et al. 1989) or tryptic soya broth (TSB). For the nitrogen-limited condition, a modification of R2 solid medium was used. The composition of this modified medium, per liter, was: sucrose 103 g; K2SO4 0.25 g; MgCl2·6H2O 10.12 g; glucose 10 g; KH2PO4 7.2 mM; CaCl2·2H2O 2.94 g; l-proline 1.3 mM; TES buffer 5.73 g; trace element solution 0.2 ml; total nitrogen: 1.3 mM; pH 7.2. For the nitrogen-rich condition, nitrogen-limited R2 medium was supplemented with Difco casamino acids of 2 g (BD, NJ, USA). For growth and antibiotics production in liquid, SMM was used. The composition of this medium, per liter, was: PEG6000 49.95 g; MgSO4·7H2O 0.6 g; TES buffer 5.73 g; glucose 10 g; trace element solution 0.1 ml; NaH2PO4 + K2HPO4 (50 mM each) 50 ml; Difco casamino acids 2 g; total phosphate 5 mM; total nitrogen 2 g; pH 7.2. For the analysis under nitrogen-limited or phosphate-limited condition, the S. coelicolor strains were grown in SMM medium until mid-exponential phase (OD450nm ≈ 0.5), and subjected to nitrogen or phosphate depletion by rapidly transferring the cultures to nitrogen-limited SMM (without Difco casamino acids) or phosphate-limited SMM (total phosphate concentration was reduced to 0.05 mM).
If necessary, 25 mg ml−1 kanamycin, 25 mg ml−1 thiostrepton or 25 mg ml−1 apramycin was added to the growth medium.
E. coli was grown in Luria broth at 37°C, supplemented with 100 μg ampicillin ml−1, 50 μg apramycin ml−1, 25 μg chloramphenicol ml−1, 50 μg kanamycin ml−1, and 50 μg streptomycin ml−1 if necessary.
Plasmid construction
The plasmids and cosmids used in this work are listed in Table 2. The primers used in this study are listed in Table 3.
The sigT promoter was cloned by PCR amplification with primers F3 and R3, and cloned into pTA2 to yield pMRB4, which was verified by sequencing. pMRB4 was digested with BglII and XbaI, and the resulting 140 bp DNA fragment was cloned into the vector pMRD279 containing the xylE reporter gene (encoding catechol 2,3-dioxygenase, which converts colorless catechol into the yellow-colored 2-hydroxymuconic semialdehyde), yielding pMRB3. pMRB3 was introduced into the S. coelicolor strains M145 and ∆sigT by conjugation, yielding S. coelicolor strains MRB4 and MRB5. As a positive control, plasmid pMRD278, containing the ermEp* (the strong constitutive promoter drives the expression of ermE gene, which is 23S rRNA dimethylase gene from Saccharopolyspora erythraea), was also conjugated into the S. coelicolor strain M145, yielding the S. coelicolor strain MRB3. As a negative control, pMRD279 was introduced into S. coelicolor strain M145 yielding MRB6.
To construct the complement rstB mutant, the rstB gene was amplified by PCR using primers F6 and R6 with NdeI sites and cloned into pTA2 to generate pMRB5. pMRB5 was digested using NdeI, and the resulting DNA fragment was subcloned into pLM1, yielding pMRB6.
Construction and complementation of the in-frame rstB deletion mutant
The disruption of rstB gene was achieved using the λRED-mediated PCR targeting system developed by Gust et al. (2003). Two primers, F1 and R1, were used to amplify the cassette aadA/oriT from plasmid pIJ778 to replace the rstB gene in the cosmid N03-16, giving the plasmid pRMB1. The cassette in pMRB1 was further removed by FLP recombinase, leaving an 81-bp “scar” sequence. The resulting plasmid, named pMRB2, was then conjugated into M145. The double-crossover replacements in S. coelicolor were verified by Southern blotting and PCR analysis (Fig. S1). One positive strain was designated MRB1.
To complement the mutant, pMRB6, a derivative of pIJ8630 containing the functional rstB gene was transferred into S. coelicolor strain MRB1 by conjugation. Insertion of complementation constructs was confirmed by PCR.
Fermentation and antibiotic quantification
For production of secondary metabolites, fermentation of S. coelicolor strains was carried out as described previously (Kieser et al. 2000). Antibiotic assays were performed as previously described (Kieser et al. 2000).
Isolation and manipulation of DNA
Streptomyces genomic DNA was isolated as described previously (Kieser et al. 2000). DNA restriction and modifying enzymes were used as recommended by the manufacturers (Sangon and Takara). Standard recombinant DNA techniques were used as previously described (Sambrook et al. 2000). DNA fragments were purified from agarose gels with the Gel Purification Kit (Axygen). Southern blotting was carried out as previously described (Sambrook et al. 2000).
RNA isolation
Total RNA was isolated from mycelia harvested from R2YE plates over cellophane or SMM and TSB liquid cultures. Total RNA was prepared as described by Mao et al. (2009). RNA concentrations were determined by absorbance at 260 nm. The integrity of the RNA was indicated by visualization of sharp rRNA bands after electrophoresis in agarose containing 2.2 M formaldehyde.
RT-PCR and quantitative real-time PCR
DNase I-treated RNA (1 μg) was used as a template for reverse transcription (RT) at 42°C with the Primer-Script RT-PCR kit (Takara) and random hexamer primers. The resulting cDNA was used for PCR amplification with sequence-specific primers under the following conditions: 95°C for 3 min, followed by 35 cycles of 95°C for 15 s, 55°C for 30 s, and 72°C for 30 s. DNase I-treated RNA samples that had not been subjected to RT were used as negative controls. Genomic DNA samples were used as positive controls.
Real-time PCRs were carried out on an iCycler MyiQ2 real-time PCR detection system (Bio-Rad Laboratories), and the data were analyzed using the software provided by the supplier. Assays were performed using 1 μl of template cDNA, 5 pmol of each primer, 0.5× SYBR Green (Takara), and 1 U of Taq polymerase (Takara) in a final volume of 25 μl. Thermal cycle conditions were as follows: 95°C for 3 min, followed by 40 cycles of 95°C for 15 s, 52°C for 20 s, and 72°C for 20 s. A final dissociation was run to generate a melting curve and verify the specificity of the amplification product. All samples were run in triplicate. The threshold cycle (CT) value was calculated for each sample. A ∆CT value was calculated by subtracting the mean CT value of the target gene from the mean CT value of the hrdB reference gene (Kelemen et al. 1996). Relative quantification of gene expression was determined using the 2–∆CT method (Livak and Schmittgen 2001).
5′ RACE experiment
Transcription start sites (TSS) were determined using the 3′/5′ RACE kit (Roche) according to manufacturer’s instructions. After treatment with DNase I (Takara), RNA was reverse-transcribed using a gene-specific primer, purified, and poly(dA) tailed at 3′ ends. The resulting cDNA was amplified by PCR using a poly(dT) primer and a gene-specific primer complementary to a region upstream of the original cDNA primer. Amplicons from the first PCR were submitted to a second PCR using the poly(dT) primer and a distinct gene-specific nested primer. PCR products were ligated into the pTA2 vector (Toyobo). Ten distinct clones were sequenced, generally resulting in the same TSS.
Promoter activity assay using the reporter xylE gene
The catechol-2,3-dioxygenase activity of the xylE reporter gene was measured as previously described to quantify promoter activity (Kieser et al. 2000).
Results
Transcriptional analysis of sigT, rstB, and rstA
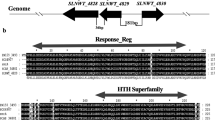
In the S. coelicolor chromosome, rstB (SCO3893), sigT (SCO3892), and rstA (SCO3891) are physically linked and have the same transcriptional direction (Fig. 1a). Amino acid homology analysis suggested that SigT is a structural homologue of SigM in other actinobacteria (data not shown). Domain prediction with COG (Tatusov et al. 2000) and Pfam databases revealed that rstB probably encode a serine/threonine protein kinase (STPK) (COG0515, pfam NO PF00069.18). Proteins of the STPK family are believed to be implicated in signal sensing and transduction (Barthe et al. 2010). RstA, being a member of zinc-associated anti-sigma factor (ZAS) family (Zdanowski et al. 2006; Dona et al. 2008), has been demonstrated to interact with SigT in vivo and in vitro, suggesting it being a putative anti-sigma factor against SigT (Mao et al. 2009).

Organization and transcription of three genes in sigT cluster of S. coelicolor. Physical map of the sigT cluster (sigT, rstA, and rstB) in S. coelicolor. The transcription directions of these three genes were indicated by filled arrowheads. b RT-PCR of transcripts overlapping the consecutive adjacent genes in the sigT cluster. RNA of strain M145 was isolated from transition phase cultures and reverse-transcribed into cDNA. The cDNA, and M145 chromosomal DNA were used as templates. Three paired primers (F4/R4, F8/R7, and F7/R7) were used for amplification of segments as indicated in (a). c Expression of sigT, rstA, and rstB in different S. coelicolor strains. S. coelicolor strains ΔsigT, ΔrstA, ΔrstB, and M145 were grown in TSB liquid medium, and total RNA was isolated from the transition phase cultures and transcribed into cDNA. Lanes 1, 2, 3, and 4 indicated the M145, ΔrstA, ΔsigT, and ΔrstB genetic backgrounds. Lane 5 is a positive control with genomic DNA from M145 as a template. The hrdB gene was used as an internal control
Although rstB, sigT, and rstA are closely linked, whether these three genes are co-transcribed as an operon has not been documented. To this end, RT-PCR analysis was performed with total RNA obtained from transition-phase cultures of wild-type strain. A 742-bp cDNA product covering the sigT-rstB intergenic region was obtained (Fig. 1b). Co-transcription analysis of rstA and sigT using primer pairs F8 and R7 failed to produce the expected 932 bp product. Nevertheless, the expected product was detected when using a forward primer F7, around 40 bp upstream of the sigT translation stop codon (Fig. 1b). These results suggested that sigT is likely not co-transcribed with rstA but with rstB, and one or more transcription start sites of rstA may lies within the coding region of sigT.
In most cases, genes in the vicinity of the genes coding for sigma factors of S. coelicolor were found functionally related to sigma factors (Helmann 2002). So, to obtain an overall picture of transcription of the genes probably related to sigT, we determined transcription of rstB, sigT and rstA in different genetic backgrounds such as the parental strain M145, and the deletion mutants, ∆sigT, ∆rstA, and ∆rstB. The expression of rstB was not detected in the ΔsigT mutant (Fig. 1c). Considering that rstB is cotranscribed with sigT, one reasonable explanation for the absence of transcript of rstB was that SigT might direct its own transcription. The rstA mutation as well as rstB mutation showed almost no effect on the expression of sigT under this normal condition (Fig. 1c).
Based on the co-transcription profile, we decided to map the TSS of sigT and rstA by 5′ RACE experiments. Two TSSs upstream of rstB, named sigTp1 and sigTp2, were identified (Fig. 2a, Fig. S3a and b). sigTp1 is more distal and sigTp2 is more proximal to the predicted translation initiation codon of rstB with relative positions of −28 and −74, respectively. The deduced −35 regions of these promoters lack a conserved AAC motif of most ECF sigma factor promoters (Helmann 2002).

Mapping of the 5′end of the sigT and rstA mRNA by 5′RACE analysis. RNA was isolated from transition phase cultures of strain M145 grown in TSB medium and equal amount (5 μg) of total RNA were used for 5′RACE analysis. a Nucleotide sequence of the promoter region of sigT and rstB. Transcription start sites of sigT are denoted as sigTp1 and sigTp2. The putative −10 and −35 regions and ribosome binding site are indicated with boxes. The translation initiation codon of rstB is underlined. b Nucleotide sequence of the promoter region of rstA. Transcription start sites of rstA are denoted by rstAp1, rstAp2, and rstAp3. The translation initiation codon of rstA and translation stop codon of sigT are underlined
Three TSSs, named rstAp1, rstAp2, and rstAp3, were identified for rstA (Fig. 2b, Fig. S3c, d and e). rstAp1, rstAp2 and rstAp3 are located at nucleotides −8, −79, and −165, relative to the translation start site of rstA, respectively. These results corroborated that rstA is not co-transcribed with sigT in a single operon. No distinct band was detected when using a reverse primer internal to the sigT ORF.
Phenotypic analysis of the S. coelicolor rstB, sigT, and rstA mutant upon nitrogen stresses
Mao et al. (2009) reported the over-production of antibiotics (Act and Red) and accelerated morphological development in ∆sigT and ∆rstA mutants on R2YE medium suggesting SigT and RstA were both involved in negative regulation on differentiation and secondary metabolism in S. coelicolor and these features appeared to be medium-dependent.
The fact that ECF sigma factor could respond to environmental stresses (Helmann 2002) led us to evaluate the potential role of SigT under various stress conditions. It was reported that a sigT mutant was sensitive to diamide, suggesting that SigT like SigR was also involved in the response to oxidative stress (Helmann 2002). However, no increased sensitivity with respect to wild type was observed when the ∆sigT mutant was exposed to diamide and hydrogen peroxide (data not shown) (Paget et al. 1998). To investigate if SigT was a functional homologue of SigM in B. subtilis, the ∆sigT mutant was exposed to high concentration salt, the strains was not impaired in growth compared to wild type (data not shown) (Horsburgh and Moir 1999; Thackray and Moir 2003).
Given that nitrogen stress plays an important role on production of antibiotics (Chakraburtty and Bibb 1997), we tested the ∆sigT mutant on nitrogen-limited and nitrogen-rich R2 medium. To exclude the effects of phosphate, excess phosphate (7.2 mM) was added into R2 medium as mentioned in methods. Under the nitrogen-limited condition, the ΔsigT mutant produced less Act than the parental strain M145 (Fig. 3c). While under the nitrogen-rich condition, the ΔsigT mutant exhibited the same phenotype as wild type (Fig. 3b). Notably, the ΔrstA mutant produced markedly less Act than the ΔsigT mutant (Fig. 3c).

Phenotype of S. coelicolor M145, ∆rstB, ∆sigT, and ΔrstA mutants and complemented rstA +, sigT +, and rstB + transformants on modified versions of R2 solid medium. a Plate legend, b growth of different S. coelicolor strains on nitrogen-rich R2 medium (Difco casamino acids: 2 g) for 4 days. c Growth of different S. coelicolor strains on nitrogen-limited R2 medium (total nitrogen: 1.3 mM, KH2PO4 7.2 mM) for 12 days
The production of the blue Act pigments (Bystrykh et al. 1996) or the development of aerial mycelium in the ΔsigT mutant could be restored to wild-type level by the introduction of the sigT gene into the ΔsigT mutant which gave rise to S. coelicolor strain sigT +, confirming that these phenotype was caused by mutation of the gene.
Growth and antibiotic production kinetics of ΔsigT, ΔrstA, and ΔrstB mutants upon nutritional downshift
To further assess the effect of the sigT mutation on antibiotic production under phosphate- and nitrogen-limited conditions, M145 and ΔsigT mutant were grown in SMM liquid medium until mid-exponential phase (optical density at 450 nm [OD450], 0.5) and subjected to amino acid shift-down (detail in methods) (Chakraburtty and Bibb 1997; Ryu et al. 2007). The level of antibiotics Act and Red was measured after nutritional depletion. Under nitrogen deprivation, the sigT deletion resulted in abolishment of Act production (Fig. 4a, b). This lowered Act production was also observed for the ΔrstA mutant but not for the ΔrstB mutant (Fig. 4c, d). The production of Red was barely detectable either in the wild type or in the ΔsigT mutant upon nitrogen deprivation.
Following nitrogen depletion, growth retardation was observed for the ΔsigT mutant throughout the course of experiment (Fig. 4c).

Growth and antibiotic synthesis kinetics of S. coelicolor strain ΔsigT and M145 grown in SMM after nitrogen or phosphate shiftdown. The S. coelicolor strains were grown in SMM until mid-exponential phase (OD450nm ≈ 0.5), and subjected to nitrogen shiftdown by rapidly transferring the cultures to nitrogen-limited SMM (without Difco casamino acids), a growth and actinorhodin level of S. coelicolor M145 and ΔsigT under nitrogen limitation. b Photograph showing the difference in antibiotic synthesis between the M145 and ΔsigT under nitrogen limitation. sigT + indicates the complementation of the ΔsigT mutant. c Growth and actinorhodin level of S. coelicolor mutants ΔrstA and ΔrstB under nitrogen limitation. d Photograph showing the difference in antibiotic synthesis between ΔrstA and ΔrstB under nitrogen limitation. Filled triangles, S. coelicolor mutant ΔsigT; open triangles, S. coelicolor M145; filled squares, S. coelicolor mutant ΔrstB; open squares, S. coelicolor mutant ΔrstA. Vertical error bars correspond to the standard error of the mean of four replicated cultures
To summarize, these data obtained from batch culture was in agreement with what observed in surface-grown culture, suggesting SigT was responsive to nitrogen starvation.
sigT gene expression is induced by nitrogen stress in auto-regulated manner
The roles of sigT on S. coelicolor development led us to investigate its expression during cell differentiation. The transcription of sigT was monitored by RT-PCR during differentiation of S. coelicolor M145 on solid R5 medium. This transcription was increased along with the development of aerial mycelium and peaked at 48 h, around the time when antibiotic production is most abundant (Bibb 1996). Then it decreased rapidly (Fig. S2), suggesting that SigT may participate in the regulation of antibiotic synthesis.
Using the reporter xylE gene coupled to the sigT promoter (Sola-Landa et al. 2005), the regulatory effects of phosphate or nitrogen on expression of sigT genes were studied in S. coelicolor M145 and the ΔsigT mutant in SMM. Nutrient deprivation was performed the same as described in the Materials and methods section. Results showed that nitrogen limitation caused a fivefold induction of the sigT promoter activity with respect to that under the nitrogen-rich condition during the transition stage (Fig. 5a). This xylE expression level paralleled that in the positive control strain MRB3 (M145/ermEp*::xylE) (data not shown). In contrast, no induction of sigT promoter activity was observed for phosphate stress (Fig. 5a). The SigT dependence of the nitrogen stress response at sigT promoter was tested by introducing the sigTp-xylE construct into the ∆sigT mutant, and induction was carried out under nitrogen starvation. The level of xylE expression in the sigT null mutant was not increased by nitrogen stress (Fig. 5a). The same induction of sigT upon nitrogen depletion was also observed in RT-PCR experiments using exponential-phase cultures of ΔsigT strain (Fig. 5b). Interestingly, the induction of sigT was also absent in the ΔrstA mutant upon nitrogen starvation as observed for the ΔsigT mutant in promoter activity assay (Figs. 5b and 6b). In contrast, the ΔrstB mutant displayed the similar induction of sigT with that in wild-type strain (Figs. 5b and 6b).

Expression of sigT was induced by growth in the nitrogen-limited SMM in a SigT-dependent manner. a Determination of sigT promoter activity using the xylE gene (encodes catechol dioxygenase) as reporter. For analysis of promoter activity in response to nitrogen or phosphate starvation, the S. coelicolor strains were grown in SMM until mid-exponential phase (OD450nm ≈ 0.5), and subjected to nitrogen or phosphate depletion as described in methods. The enzyme activity was measured at the indicated growth phases. Note that there is a drastic induction in sigT promoter activity under nitrogen limitation with respect to the negative control (MRB6) and that under nitrogen-rich condition. The promoter of the sigT gene was dependent on SigT as the above induction effect was abolished in the ΔsigT genetic background (MRB5). b qRT-PCR analysis of expression of sigT in M145 and different mutant strains upon different nutritional stress. The results confirmed that sigT was responsive to nitrogen stress. p (+) phosphate-replete condition, p (−) phosphate-limited condition, n (+) nitrogen-replete condition, n (−) nitrogen-limited condition. EX, TR, and ST indicate exponential, transition, and stationary phases of growth, respectively. Results described in (a) and (b) represent the mean value of at least three independent experiments, each performed in duplicate, and the vertical bars indicate the standard error

Relative mRNA levels measured by semi-quantitative and quantitative RT-PCR of differentially expressed genes upon nitrogen limitation in S. coelicolor M145, ΔsigT, ΔrstA, and ΔrstB strains. Total RNA was prepared from different S. coelicolor strains at exponential, transition, and stationary stages of growth in SMM after amino acid depletion then reverse-transcribed into cDNA. a–b show the expression of sigT under nitrogen-limited conditions and the control condition, respectively, c–d show the expression of actII-ORF4 and relA, e shows the expression of hrdB which was used as an internal control. f–g show the relative mRNA levels of the gene actII-ORF4 and relA in different S. coelocolor strains quantified by RT-PCR. EX, TR, ST indicate exponential, transition, and stationary phases of growth, respectively. Error bars correspond to the standard error of the mean of four culture replicates
SigT regulates relA to response nitrogen stress
Several studies have documented that highly phosphorylated guanine nucleotides ppGpp play a central role in triggering antibiotic biosynthesis in S. coelicolor under stringent response (Ochi 1990; Strauch et al. 1991). The mutation in the ppGpp synthetase gene relA would impair both antibiotic production and the morphological differentiation of S. coelocolor. The former was ascribed to a marked decline transcription of genes actII-ORF4 and redD regulating production of Act and Red, respectively (Chakraburtty and Bibb 1997). The coincidence of antibiotic production in the ΔsigT mutant with that in the ΔrelA mutant upon nitrogen starvation led us to probe a possible link between sigT and relA by RT-PCR analysis. Meanwhile, given that the ΔrstA mutant displayed a similar pattern of antibiotic production to the ΔsigT mutant upon nitrogen starvation, transcription analysis was also applied to the ΔrstA mutant and the ΔrstB mutant. The ΔsigT mutant displayed expression of relA and actII-ORF4 around eight times and 20 times less than the wild type as quantified by RT-PCR experiments (Fig. 6f, g). The ΔrstA mutant followed a similar transcription pattern to the ΔsigT mutant (around six times less than wild type both in expression of relA and actII-ORF4). Conversely, the transcription in the ΔrstB mutant was not affected by nitrogen starvation (Fig. 6f and g). We did not detect apparent difference on redD transcription between the ΔsigT mutant and wild-type strain (data not shown).
Discussion
The evidences presented in this study suggest that the ECF sigma factor SigT is responding to the nitrogen stress and essential for normal Act production under nitrogen limitation. ECF sigma factors of S. coelicolor typically regulate a response to environmental stress (es) (Missiakas and Raina 1998; Helmann 2002). In consequence, in most cases, the absence of ECF sigma factor gene usually does not result in obvious phenotypes under normal conditions. To identify which stress is SigT in responding to, we exposed the ΔsigT mutant to various stress conditions such as oxidative stress, osmotic stress, and nutrient stress. The ΔsigT mutant only displayed a marked mutant phenotype upon nitrogen starvation. Our data showed that the sigT mutation did indeed impair Act production upon nitrogen limitation. While under a nitrogen-replete condition, the production of Act in the ΔsigT mutant was restored to the level of wild-type M145, indicating that this deficiency in Act production was nitrogen-dependent. qRT-PCR combined with promoter activity assay revealed that sigT was only induced after nitrogen limitation. These data confirmed that SigT could respond to nitrogen stress. To our knowledge, it is the first report of an ECF sigma factor involved in response to nitrogen starvation in S. coelicolor.
The absence of the transcript of rstB (rstB is co-transcribed with sigT) combined with the abolishment of induction of sigT promoters upon nitrogen deprivation in the ΔsigT mutant background was indicative of the sigT promoter to be SigT-dependent and demonstrated that SigT could control its own expression by positively auto-regulating transcription, a feature observed with most ECF sigma factors (Helmann 1999, 2002). In 5′RACE experiments, two promoters were identified for the sigT gene. Pioneering works had uncovered many ECF sigma factor genes such as sigB and sigR were also transcribed from two promoters, in which one promoter is responsible for constitutive transcription, and the other appears to be induced by a stress condition and to be auto-regulated (Paget et al. 1998; Cho et al. 2001). So, it is conceivable that only one promoter of sigT could be induced by stress condition and auto-regulated in analogy to the sigR or sigB promoters. Then, the key issue is to determine which promoter is SigT-dependent.
The stringent response plays a central role in responding to nutrient stress in bacteria and mediates its effect through the alarmone ppGpp (Toulokhonov et al. 2001; Braeken et al. 2006). ppGpp can bind to the β-subunit of RNA polymerase (Toulokhonov et al. 2001) and thereby triggers the initiation of antibiotic production (Bibb 2005). So the disruption of ppGpp synthetase gene relA resulted in deficient production of antibiotics (Act and Red) of S. coelicolor upon nitrogen limitation (Chakraburtty and Bibb 1997). This defect in antibiotic production was due to the markedly reduced transcriptions of pathway-specific regulatory genes actII-ORF4 and redD, for Act and Red production respectively, in the ∆relA mutant (Chakraburtty and Bibb 1997). In this study, the ∆sigT mutant showed a phenotypic similarity with the ∆relA mutant, in that both are deficient in Act production and both display delayed aerial mycelium development. In addition, sigT is also induced by nitrogen deprivation similar to relA (Chakraburtty et al. 1996). These evidences suggested a link between SigT and RelA. Transcription analysis confirmed this link in that the expression of relA was dramatically declined in the ΔsigT mutant compared to the wild type upon nitrogen starvation. These results suggest that the observed phenotypes of the sigT mutant upon nitrogen starvation may result primarily from the absence of relA and accordingly expression of relA is SigT-dependent.
The genes neighboring to an ECF sigma factor gene often encode the regulatory proteins of the sigma factor, such as an anti-sigma factor or an anti-anti-sigma factor. For instance, in the case of SigB, rsbA and rsbB upstream of the sigB gene encode an anti-sigma factor and an anti-anti-sigma factor for SigB, respectively (Lee et al. 2004). Previous work suggested that RstA encoded by the gene downstream of sigT may be a putative anti-sigma factor for SigT of S. coelicolor (Mao et al. 2009). To probe the function of RstB encoded by the gene upstream of sigT, the rstB gene was disrupted. Act production and actII-ORF4 as well as relA transcription of the ∆rstA mutant under nitrogen limitation resembled well those of the ∆sigT mutant and were in contrast to those of ∆rstB mutant. In the transcriptional analysis under nitrogen starvation, we found the expression of sigT was reduced dramatically in the ∆rstA mutant in sharp contrast to that in the ∆rstB mutant. Giving that SigT is responsible for induction of its own structural gene as described above, the transcriptional profiles of sigT displayed in the ∆rstA mutant and the ∆rstB mutant may suggest that RstA regulates the activity of SigT positively and RstB is the negative regulator of SigT. Furthermore, co-transcriptional analysis and 5′RACE assay revealed that sigT was co-transcribed with rstB but not rstA. The data presented here were not in consistence with the notions that an anti-sigma factor gene is encoded with its cognate sigma factor gene in one operon and that an anti-sigma factor antagonizes the activity of sigma factor (Hughes and Mathee 1998). This raised the possibility that RstA may be just a positive regulator of SigT rather than an anti-sigma factor against SigT despite that it shares structural similarity with members of the ZAS family such as RsrA and that it could interact with SigT in vivo and in vitro (Kang et al. 1999; Mao et al. 2009). RstB was predicted to be a sensor STPK by bioinformatics analysis. We also found that RstA could interact with RstB in vitro (Mao et al. unpublished work). We propose from above evidences that RstA and RstB may constitute a partner pair in signal transduction for regulating expression of sigT transcriptionally in a similar manner with the CseB–CseC two-component signal transduction system in the SigE case, where CseC serve as a sensor histidine protein kinase and CseB serve as a response regulator (Hong et al. 2002; Paget et al. 1999b).
To conclude, our study demonstrated that sigT was induced by nitrogen stress in a SigT-dependent manner. SigT may activate the Act production under nitrogen limitation via activating the expression of relA gene directly or indirectly. Thus, SigT appears to link nutrient stress to antibiotic production by Streptomyces. Our results also suggested that the putative anti-sigma factor RstA may be a positive regulator of SigT at the transcriptional level. Although the detailed molecular mechanisms on regulation of Act production by SigT upon nitrogen stress remains unclear, our findings give a clue to understand the complex regulation system of antibiotic biosynthesis in S. coelicolor upon stringent response to nitrogen starvation. Further work will focus on defining the SigT regulon through in vitro transcription assays.
References
Alvarez-Martinez CE, Baldini RL, Gomes SL (2006) A Caulobacter crescentus extracytoplasmic function sigma factor mediating the response to oxidative stress in stationary phase. J Bacteriol 188(5):1835–1846
Alvarez-Martinez CE, Lourenco RF, Baldini RL, Laub MT, Gomes SL (2007) The ECF sigma factor σT is involved in osmotic and oxidative stress responses in Caulobacter crescentus. Mol Microbiol 66(5):1240–1255
Anthony JR, Warczak KL, Donohue TJ (2005) A transcriptional response to singlet oxygen, a toxic byproduct of photosynthesis. Proc Natl Acad Sci U S A 102(18):6502–6507
Bang IS, Frye JG, McClelland M, Velayudhan J, Fang FC (2005) Alternative sigma factor interactions in Salmonella: sigma and sigma promote antioxidant defences by enhancing sigma levels. Mol Microbiol 56(3):811–823
Barthe P, Mukamolova GV, Roumestand C, Cohen-Gonsaud M (2010) The structure of PknB extracellular PASTA domain from mycobacterium tuberculosis suggests a ligand-dependent kinase activation. Structure 18(5):606–615
Bibb M (1996) The regulation of antibiotic production in Streptomyces coelicolor A3(2). Microbiology 142:1335–1344
Bibb MJ (2005) Regulation of secondary metabolism in Streptomycetes. Curr Opin Microbiol 8(2):208–215
Braeken K, Moris M, Daniels R, Vanderleyden J, Michiels J (2006) New horizons for (p)ppGpp in bacterial and plant physiology. Trends Microbiol 14(1):45–54
Bystrykh LV, Fernandez-Moreno MA, Herrema JK, Malpartida F, Hopwood DA, Dijkhuizen L (1996) Production of actinorhodin-related “blue pigments” by Streptomyces coelicolor A3(2). J Bacteriol 178(8):2238–2244
Chakraburtty R, Bibb M (1997) The ppGpp synthetase gene (relA) of Streptomyces coelicolor A3(2) plays a conditional role in antibiotic production and morphological differentiation. J Bacteriol 179(18):5854–5861
Chakraburtty R, White J, Takano E, Bibb M (1996) Cloning, characterization and disruption of a (p)ppGpp synthetase gene (relA) of Streptomyces coelicolor A3(2). Mol Microbiol 19(2):357–368
Chater KF (2001) Regulation of sporulation in Streptomyces coelicolor A3(2): a checkpoint multiplex? Curr Opin Microbiol 4(6):667–673
Cho YH, Lee EJ, Ahn BE, Roe JH (2001) σB, an RNA polymerase sigma factor required for osmoprotection and proper differentiation of Streptomyces coelicolor. Mol Microbiol 42(1):205–214
Dona V, Rodrigue S, Dainese E, Palù G, Gaudreau L, Manganelli R, Provvedi R (2008) Evidence of complex transcriptional, translational, and posttranslational regulation of the extracytoplasmic function sigma factor σE in Mycobacterium tuberculosis. J Bacteriol 190(17):5963–5971
Doull JL, Vining LC (1990) Nutritional control of actinorhodin production by Streptomyces coelicolor A3(2): suppressive effects of nitrogen and phosphate. Appl Microbiol Biotechnol 32(4):449–454
Floriano B, Bibb M (1996) afsR is a pleiotropic but conditionally required regulatory gene for antibiotic production in Streptomyces coelicolor A3(2). Mol Microbiol 21(2):385–396
Gust B, Challis GL, Fowler K, Kieser T, Chater KF (2003) PCR-targeted Streptomyces gene replacement identifies a protein domain needed for biosynthesis of the sesquiterpene soil odor geosmin. Proc Natl Acad Sci U S A 100(4):1541–1546
Hecker M, Völker U (1998) Non-specific, general and multiple stress resistance of growth-restricted Bacillus subtilis cells by the expression of the σB regulon. Mol Microbiol 29(5):1129–1136
Helmann JD (1999) Anti-sigma factors. Curr Opin Microbiol 2(2):135–141
Helmann JD (2002) The extracytoplasmic function (ECF) sigma factors. Adv Microb Physiol 46:47–110
Hobbs G, Frazer CM, Gardner DC, Cullum JA, Oliver SG (1989) Dispersed growth of Streptomyces in liquid culture. Appl Microbiol Biotechnol 31(3):272–277
Hong HJ, Paget MS, Buttner MJ (2002) A signal transduction system in Streptomyces coelicolor that activates the expression of a putative cell wall glycan operon in response to vancomycin and other cell wall-specific antibiotics. Mol Microbiol 44(5):1199–1211
Horsburgh MJ, Moir A (1999) σM, an ECF RNA polymerase sigma factor of Bacillus subtilis 168, is essential for growth and survival in high concentrations of salt. Mol Microbiol 32(1):41–50
Hughes KT, Mathee K (1998) The anti-sigma factors. Annu Rev Microbiol 52:231–286
Ishihama A (2000) Functional modulation of Escherichia coli RNA polymerase. Annu Rev Microbiol 54:499–518
Kang JG, Paget MS, Seok YJ, Hahn MY, Bae JB, Hahn JS, Kleanthous C, Buttner MJ, Roe JH (1999) RsrA, an anti-sigma factor regulated by redox change. EMBO J 18(15):4292–4298
Kelemen GH, Brown GL, Kormanec J, Potuckova L, Chater KF, Buttner MJ (1996) The positions of the sigma-factor genes, whiG and sigF, in the hierarchy controlling the development of spore chains in the aerial hyphae of Streptomyces coelicolor A3(2). Mol Microbiol 21(3):593–603
Kieser T, Bibb M, Buttner M, Chater K, Hopwood D (2000) Practical Streptomyces genetics. The John Innes Foundation, Norwich, UK
Kormanec J, Sevcikova B, Halgasova N, Knirschova R, Rezuchova B (2000) Identification and transcriptional characterization of the gene encoding the stress-response sigma factor σH in Streptomyces coelicolor A3(2). FEMS Microbiol Lett 189(1):31–38
Lee EJ, Cho YH, Kim HS, Ahn BE, Roe JH (2004) Regulation of σB by an anti- and an anti-anti-sigma factor in Streptomyces coelicolor in response to osmotic stress. J Bacteriol 186(24):8490–8498
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔC(T) method. Methods 25(4):402–408
MacNeil DJ, Occi JL, Gewain KM, MacNeil T, Gibbons PH, Ruby CL, Danis SJ (1992) Complex organization of the Streptomyces avermitilis genes encoding the avermectin polyketide synthase. Gene 115(1–2):119–125
Mao XM, Zhou Z, Cheng LY, Hou XP, Guan WJ, Li YQ (2009) Involvement of σT and RstA in the differentiation of Streptomyces coelicolor. FEBS Lett 583(19):3145–3150
Missiakas D, Raina S (1998) The extracytoplasmic function sigma factors: role and regulation. Mol Microbiol 28(6):1059–1066
Ochi K (1990) A relaxed (rel) mutant of Streptomyces coelicolor A3(2) with a missing ribosomal protein lacks the ability to accumulate ppGpp, A-factor and prodigiosin. J Gen Microbiol 136(12):2405–2412
Paget MSB, Kang JG, Roe JH, Buttner MJ (1998) σR, an RNA polymerase sigma factor that modulates expression of the thioredoxin system in response to oxidative stress in Streptomyces coelicolor A3(2). EMBO J 17(19):5776–5782
Paget MS, Chamberlin L, Atrih A, Foster SJ, Buttner MJ (1999a) Evidence that the extracytoplasmic function sigma factor σE is required for normal cell wall structure in Streptomyces coelicolor A3(2). J Bacteriol 181(1):204–211
Paget MS, Leibovitz E, Buttner MJ (1999b) A putative two-component signal transduction system regulates σE, a sigma factor required for normal cell wall integrity in Streptomyces coelicolor A3(2). Mol Microbiol 33(1):97–107
Ryu YG, Kim ES, Kim DW, Kim SK, Lee KJ (2007) Differential stringent responses of Streptomyces coelicolor M600 to starvation of specific nutrients. J Microbiol Biotechnol 17(2):305–312
Sambrook J, MacCallum P, Russell D (2000) Molecular cloning: a laboratory manual, 3rd edn. Cold Spring Harbor Laboratory Press, NY, US
Sola-Landa A, Rodriguez-Garcia A, Franco-Dominguez E, Martin JF (2005) Binding of PhoP to promoters of phosphate-regulated genes in Streptomyces coelicolor: identification of PHO boxes. Mol Microbiol 56(5):1373–1385
Strauch E, Takano E, Baylis HA, Bibb MJ (1991) The stringent response in Streptomyces coelicolor A3(2). Mol Microbiol 5(2):289–298
Sun J, Kelemen GH, Fernandez-Abalos JM, Bibb MJ (1999) Green fluorescent protein as a reporter for spatial and temporal gene expression in Streptomyces coelicolor A3(2). Microbiology 145:2221–2227
Takano E, Gramajo HC, Strauch E, Andres N, White J, Bibb MJ (1992) Transcriptional regulation of the redD transcriptional activator gene accounts for growth-phase-dependent production of the antibiotic undecylprodigiosin in Streptomyces coelicolor A3(2). Mol Microbiol 6(19):2797–2804
Tatusov RL, Galperin MY, Natale DA, Koonin EV (2000) The COG database: a tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res 28(1):33–36
Thackray PD, Moir A (2003) σM, an extracytoplasmic function sigma factor of Bacillus subtilis, is activated in response to cell wall antibiotics, ethanol, heat, acid, and superoxide stress. J Bacteriol 185(12):3491–3498
Toulokhonov II, Shulgina I, Hernandez VJ (2001) Binding of the transcription effector ppGpp to Escherichia coli RNA polymerase is allosteric, modular, and occurs near the N terminus of the β′-subunit. J Biol Chem 276(2):1220–1225
Wang G, Tanaka Y, Ochi K (2010) The G243D mutation (afsB mutation) in the principal sigma factor σHrdB alters intracellular ppGpp level and antibiotic production in Streptomyces coelicolor A3(2). Microbiology 156:2384–2392
Zdanowski K, Doughty P, Jakimowicz P, O’Hara L, Buttner MJ, Paget MSB, Kleanthous C (2006) Assignment of the zinc ligands in RsrA, a redox-sensing ZAS protein from Streptomyces coelicolor. Biochem 45(27):8294–8300
Acknowledgments
This work was supported by the National Sciences Foundation of China (31070040), Science Foundation of Chinese University (2009QNA6006) and National Basic Research Program of China(2012CB721005). We thank Dr. Yi-Ling Du for the many helpful discussions and technical guides. We also especially thank Prof. Zhong-Jun Qin at Institute of Plant Physiology and Ecology, Chinese Academy of Science, for his kind gift of cosmid N03-16 for rstB disruption.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 1752 kb)
Rights and permissions
About this article
Cite this article
Feng, WH., Mao, XM., Liu, ZH. et al. The ECF sigma factor SigT regulates actinorhodin production in response to nitrogen stress in Streptomyces coelicolor . Appl Microbiol Biotechnol 92, 1009–1021 (2011). https://doi.org/10.1007/s00253-011-3619-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-011-3619-2