
Abstract
The bacterium Gordonia alkanivorans RIPI90A has been previously reported as dibenzothiophene-desulfurizing strain. The present study provides a complete investigation of the dsz operon including dsz promoter analysis from desulfurization competent strain belonging to the genus Gordonia. PCR was used to amplify the dszABC genes and adaptor ligation-based PCR-walking strategy used to isolate the dsz promoter. Unlike the dsz operon of Rhodococcus erythropolis, the operon of RIPI90A was located on chromosome. Despite the remarkably high homology between dsz genes of G. alkanivorans RIPI90A and R. erythropolis IGST8, promoter sequences of the strains were not very similar. The dsz promoter of G. alkanivorans RIPI90A shows only 52.5% homology to that of R. erythropolis IGTS8 and Gordonia nitida. Deletion analysis of the dsz promoter from RIPI90A using luciferase as a reporter gene revealed that the dsz promoter was located in regions from −156 to −50.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Combustion of sulfur-containing compounds in fossil fuels emits sulfur oxides, which can cause adverse effects on health, environment, and economy. As a result, regulations for sulfur level in diesel oil have become increasingly strict (Monticello 2000; Kilbane 2006). The current industrial method used for the removal of sulfur from fuels is hydrodesulfurization, which requires high temperature and high pressure. Biodesulfurization is an environmentally friendly method that can remove sulfur from refractory organic compounds under ambient temperature and pressure without lowering the calorific value of the fuel (McFarland 1999; Chen et al. 2008). In the last decade, tremendous advances have been made in the understanding of the biochemistry and genetics of biodesulfurization, particularly the 4S pathway that metabolizes dibenzothiophene (DBT) model molecule via the sequential formation of sulfoxide, sulfone, sulfonate, and finally 2-hydroxybiphenyl (2-HBP; Oldfield et al. 1997; Mohebali and Ball, 2008). 4S pathway plays a central role in biological sulfur removal concept, and the pathway does not result in ring breakage of the carbon–carbon backbone and specifically emits sulfur from heterocycle (Soleimani et al. 2007). Much of this information is derived from the prototype DBT-biodesulfurizing strain, Rhodococcus erythropolis IGTS8 (Gupta et al. 2005). The DBT desulfurization genes of the bacterium were identified, and three desulfurization genes (dszA, dszB, and dszC) were confirmed to constitute a single operon. These genes, when cloned on a DSZ− phenotype, confer the ability to desulfurize DBT to 2-HBP. The dsz operon was found on a large plasmid of 150 kb in R. erythropolis IGTS8. Promoter and regulatory regions of the dsz operon were also studied, and it was found that enzymes are strongly repressed in the presence of readily bioavailable sulfur, i.e., sulfate, sulfide, methionine, and cysteine (Denome et al. 1994; Piddington et al. 1995; Denis-Larose et al. 1998).
Although R. erythropolis is the most extensively studied example, several other microorganisms have also been shown to catalyze DBT desulfurization; these include strains of Corynebacterium, Gordonia, Nocardia, Mycobacterium, and a thermophilic strain of Paenibacillus (Caro et al. 2007; Mohebali and Ball 2008). Since the isolation and identification of R. erythropolis IGTS8, considerable research has been conducted on molecular biology of this strain, in order to obtain a better control over the machinery of specific sulfur removal (Gallardo et al. 1997; Hirasawa et al. 2001; Yu et al. 2006). However, there has been a little report on such studies on other desulfurizing strains. We recently reported the isolation of DBT-desulfurizing bacterium, Gordonia alkanivorans strain RIPI90A. The growing and resting cells of the strain were able to desulfurize DBT to 2-HBP via the 4S pathway in aqueous and biphasic systems (Mohebali et al. 2007). Herein, we report the cloning and identification of the DNA sequence of the dsz operon of G. alkanivorans RIPI90A. The dszABC genes of G. alkanivorans were amplified by PCR and sequenced. In addition, we cloned and sequenced the dsz regulatory region of G. alkanivorans RIPI90A by PCR-walking method. The minimal promoter region was determined by deletion analysis and using the luciferase as a reporter gene.
Materials and methods
Bacterial strains, plasmids, and cultivation conditions
G. alkanivorans RIPI90A (DSMZ 01-1151) was used as the source of total DNA for cloning of dsz genes. G. alkanivorans RIPI90A was cultivated at 30°C in minimal salt (MS) medium supplemented with 0.5 mM DBT (Shavandi et al. 2009). E. coli DH5α cells were used as host for general cloning and were cultivated at 37°C on medium containing ampicillin (50 mg l−1) and kanamycin (50 mg l−1) when required.
Recombinant DNA techniques
Recombinant DNA techniques were performed as described by Sambrook and Russell (2001) or according to manufacturer’s protocols. Total DNA of Gordonia was prepared as previously described (Shavandi et al. 2009). Plasmid DNA was prepared using high pure plasmid isolation kit (Roche). PCR products were cloned with instant T/A cloneTM PCR product cloning kit (Fermentas).
PCR Amplification and cloning of dsz genes
PCR amplification was performed using a programmable DNA thermal cycler (Light Cycler, BioRad). PCR reactions and conditions were as previously described (Shavandi et al. 2009). Primers were purchased from MWG Biotech. The primers used in this work are listed in Table 1. To amplify the DNA fragment carrying the dsz gene cluster from genomic DNA of strain RIPI90A, primers were designed based on known sequences from R. erythropolis IGTS8 (GenBank U08850). Primers AF and BR were used to amplify dszA and dszB genes of strain RIPI90A. Primers BCF and C1R were used to amplify dszC gene. The dsz genes sequences of the strain RIPI90A were determined using the primer walking technique.
Isolation and sequencing of the dsz promoter
To clone the 5′ flanking sequence of the dszA gene (dsz promoter), PCR-walking strategy was used (Rishi et al. 2004). According to digestion results, restriction endonucleases, PstI, and EcoRI were used to generate several DNA fragment libraries, which were separately ligated to the corresponding adaptors. Adaptor-ligated genomic DNA fragments were subjected to an initial round of PCR amplification with the adaptor-specific primer (ASP-E and ASP-P) and an outer gene-specific primer (GSPR1). A gene-specific inner reverse primer (GSPR2) and adaptor-specific primer were used for the second round of PCR amplification. The fragments amplified by PCR were gel-purified and screened in a third PCR reaction using a gene-specific primer pair, GSPR2 and GSPF, which were amplifying the initial 100 bp of the dszA gene. Confirmed PCR fragments were ligated into pTZ57R/T vector (Fermentas), and the transformants were selected and confirmed using standard methods. Recombinant plasmid DNA was prepared, and the inserts were sequenced using pUC/M13 forward and reverse primers.
Deletion analysis of the dsz promoter
Deletion analysis of the 350-bp dsz promoter fragment was performed to define the extent of sequence necessary for promoter function (Vasicova et al. 1999). PCR was used to create truncated promoter fragments of various lengths, with appropriate restriction sites attached to the ends for cloning. The promoter fragments were digested, gel-purified, and ligated into the pRSL21 vector (Fig. 1), which was constructed by insertion of the luciferase reporter gene to the pRSG43 vector (Matsui et al. 2007; Shavandi et al. 2009), and resulted constructs were used to transform electrocompetent RIPI90A cells. The expression of the luciferase reporter gene from each construct in comparison to the cell containing promoterless reporter gene (pRSL21 plasmid) was used as a content of promoter activity.

Schematic map of the plasmid pRSL21, which was derived from pRSG43 vector by inserting a promoterless luciferase gene. Restriction sites and structural genes are indicated. Km kanamycin resistance, Luc luciferase gene, RepA and RepB Gordonia origin of replication, Ori ColE1 origin of replication
Transformation of G. alkanivorans RIPI90A
Plasmids were introduced into G. alkanivorans by electroporation using Gene pulser X-cell electroporator (BioRad). The preparation of Gordonia electrocompetent cells and electroporation conditions was as described previously (Arenskötter et al. 2003; Shavandi et al. 2009).
Measurement of luciferase activity
Activity of the full-length promoter and its deletion derivatives was evaluated by luciferase assay. Activity of promoter fragments cloned in pRSL21 vector was measured in MS medium supplemented with 0.5 mM DBT. For quantitative liquid assay, 10-ml samples of cell cultures were centrifuged, washed with 20 mM potassium phosphate buffer (pH 7.2), and the pellets were collected. All cell suspensions were prepared in potassium phosphate buffer with final optical density of 1 (OD660 = 1). After disruption of the cells by sonication, the bioluminescence emission spectrum of the luciferase reporter gene was recorded using Perkin-Elmer luminescence spectrophotometer LS 50B apparatus. A volume of 50 μl of substrate mixture consisting of 1 mM luciferin, 50 mM Tris buffer (pH 7.8), 2 mM ATP, and 10 mM MgSO4 was added to 100 μl of luciferase solution in a quartz cell. Data were collected over the wavelength range 400–700 nm (Emamzadeh et al. 2006). The spectra were automatically corrected for photosensitivity of the equipment.
Sequence analysis and bioinformatics
All sequencing was carried out in both directions to eliminate read errors. Sequencing reactions were carried out by MWG Biotech. The nucleotide sequence of the dsz operon from G. alkanivorans RIPI90A has been submitted to GenBank with assigned accession number EU364831. The DNA nucleotide sequence of the dsz genes was translated into protein sequence, and the deduced amino acid sequence was analyzed. The web-based NCBI BLAST program was used for sequence identity determination of protein and DNA sequences (Altschul et al. 1997). Multiple sequence alignments were done by CLUSTALW program.
Results
Cloning and sequencing of the DBT-desulfurization genes from G. alkanivorans RIPI90A
DBT-desulfurization genes, dszABC, have been previously cloned and sequenced from different DBT-desulfurizing strains such as Rhodococcus sp. IGTS8 (Denome et al. 1994), Gordonia nitida (Park et al. 2003), and Paenibacillus sp. A11-2 (Ishii et al. 2000). Based on the known nucleotide sequences of R. erythropolis IGTS8 (GenBank U08850), primers were synthesized and used for PCR with total DNA prepared from G. alkanivorans as template. dszAB genes were successfully PCR-amplified using the AF and BR primers. However, the dszC gene could not be amplified using the primers designed based on the R. erythropolis sequences.
A 2.5-kb amplified DNA was cloned into pTZ57R vector, and the nucleotide sequence of this PCR product was determined. Subsequently, new primers were synthesized based on these nucleotide sequences and used for PCR with G. alkanivorans RIPI90A total DNA. The dszABC genes of G. alkanivorans RIPI90A were amplified and sequenced using these Gordonia-specific primers. The overall nucleotide sequence of dszABC genes of G. alkanivorans shows 83.1% homology to dszABC of Rhodococcus sp. IGTS8 (GenBank U08850) and 83.2% to dszABC of G. nitida (GenBank AY714057). The dszA gene encodes a protein of 474 amino acids, and the corresponding gene product, DszA, shows 91% homology to DszA of R. erythropolis and 90.7% to DszA of G. nitida. The dszB gene encodes a protein of 365 amino acids, which shows 86.6% and 86.3% homology to DszB of R. erythropolis and DszB of G. nitida, respectively. The dszC gene encodes a protein of 416 amino acids, and the corresponding gene product, DszC, shows 90.2% homology to DszC of R. erythropolis and 89% to DszC of G. nitida. In the dsz operon structure of G. alkanivorans RIPI90A, the end of dszA has 64-bp overlap with the translational initiation site of dszB gene. Furthermore, there is a 13-bp gap between dszB and dszC genes of G. alkanivorans RIPI90A.
Isolation and sequencing of the dsz promoter
In an effort to isolate and sequence the dszA flanking region (dsz promoter), we used a PCR-walking technique. The G. alkanivorans RIPI90A genomic DNA was partially digested with EcoRI and PstI restriction enzymes and ligated to the corresponding adaptor. An outwarding gene-specific primer and an adaptor-specific primer were used to PCR amplify upstream flanking sequence that resulted in amplification of fragments up to 1,500 bp in two adaptor-ligated libraries that were created using EcoRI and PstI. The products of the first PCR were used as a template in second round PCR by a nested gene-specific primer and adaptor-specific primers. Fragments of 200 to 1,000 bp length were amplified in EcoRI library, but no amplification was observed in the PstI library. No amplification in the PstI library could be due to the absence of a restriction site close to the gene-specific primer (Rishi et al. 2004). The PCR-amplified fragments were cloned into pTZ57R/T plasmid and sequenced using vector-specific primers.
dsz promoter sequence analysis
Figure 2 presents the pairwise alignments of the promoter regions of G. alkanivorans RIPI90A and R. erythropolis IGTS8. While there is a complete homology between the upstream sequences of the dsz operons of R. erythropolis IGTS8 and G. nitida, the 200-bp upstream sequence of dsz operon of G. alkanivorans RIPI90A shows only 52.5% homology to corresponding region in dsz operons of R. erythropolis IGTS8 and G. nitida. The sequence similarity analysis of the 350-bp dszA upstream fragment using NCBI BLAST tool indicates that there is a transposase-like sequence in regions from −350 to −170. Several other desulfurization competent strains including Gordonia sp. CYKS2 (GenBank AY396519), G. alkanivorans strain 1B (GenBank AY678116), and Rhodococcus sp. (GenBank U08850) also have the same transposase sequences near their dsz operon.

CLUSTALW alignment of G. alkanivorans RIPI90A and R. erythropolis IGTS8 dsz promoter sequences; 200-bp sequences upstream to the starting ATG of the dszA genes were aligned. The nucleotides identical in both sequences are indicated by an asterisk
Deletion analysis of P-dsz
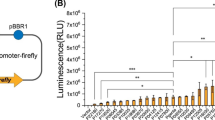
To locate the dsz promoter and determine the minimal promoter sequence and analyze the promoter activity, a series of 5′ progressive deletions was performed on the 350-bp sequenced fragment immediately upstream of the dszA gene. PCR produced successively truncated promoter fragments were fused to the promoterless luciferase reporter gene (pRSL21 plasmid). To determine the activity of the promoter fragments, these constructs were used to transform G. alkanivorans RIPI90A, and subsequently luciferase activity was measured. A schematic picture of the deletion analysis process and the luciferase activity of each promoter fragment were presented in Fig. 3. The position of the 5′ primers used for amplification of each fragment was indicated by a number at the beginning of each fragment. Luciferase activity of each clone was expressed relative to that measured with full-length fragment. The presented amounts of luciferase activity are averages from three independent measurements.

Deletion mapping of the dsz promoter of G. alkanivorans RIPI90A. The horizontal lines represent the relative lengths of DNA fragments carrying the promoter studied. The numbers at the beginning of the lines indicate the starting point of the promoter containing fragment. PCR generated fragments were cloned into pRSL21 to construct luciferase transcriptional fusions for determination of promoter activity. Luciferase activity is expressed relative to that measured with full-length fragment
Two sets of deletions were defined according to the levels of luciferase activity in the different clones. In the first set of clones, the luciferase activity in promoter fragments, 233, 190, and 156 bp lengths, with little deviations is equal to the promoter activity of the full-length promoter fragment. This result suggests that the regions from −335 to −156 have no dramatic effect on the dsz promoter activity and indicates that these regions may be not containing any part of the dsz promoter.
Deletions at the 5′ end as far as position −156 did not affect P-dsz strength, but in the second set of clones, further deletions has shown a progressive decrease in promoter activity. A remarkable reduction of activity (to 37.5%) was caused by the deletion of nucleotides between positions −115 and −75. A further deletion that removed the region −75 to −51 abolished promoter activity nearly completely (less than 6% luciferase activity).
Discussion
G. alkanivorans RIPI90A is a DBT-desulfurizing strain which converts DBT to 2-hydroxybiphenyl via the 4S pathway (Mohebali et al. 2007). In this study, the dsz operon of the strain has been PCR-amplified, cloned, and sequenced. Application of the primers designed based on R. erythropolis IGTS8 dsz sequences result in amplification of dszA and dszB genes while the dszC could not be successfully amplified. Other researchers also reported that the efforts to PCR amplify the dszC gene from Gordonia strains by primers designed using dsz gene sequences of R. erythropolis were unsuccessful. Park et al. could not amplify the dszC gene of G. nitida using Rhodococcus-based primers (Park et al. 2003). Alves et al. (2007) amplified the dszA and dszB genes of a Gordonia strain by primers based on R. erythropolis IGTS8 but they employed Gordonia-specific primers for the amplification of the dszC gene. Santos et al. (2006) also have reported the same results of PCR amplification using Gordonia sp. strain F.5.25.8 genomic DNA as template.
Recently, Kilbane and Robbins (2007) performed a comprehensive comparison of the known desulfurization proteins from three divergent desulfurizing strains namely R. erythropolis IGTS8, Bacillus subtilis WU-S2B, and Paenibacillus sp. A11-2 in order to detect conserved regions for PCR primer designing. The results indicate that there were conserved sequences at the beginning of the dszA gene and at the end of dszB gene which can be useful in primer designing and PCR amplification of the dszA and dszB gene, but there is no such conserved sequence at the beginning of the dszC gene. Lack of the conserved sequences at the beginning regions of the dszC gene could be explanation of the unsuccessful efforts for PCR amplification of the dszC gene of Gordonia strains using rhodococcal primers.
In the dsz operon structure of G. alkanivorans RIPI90A, the 64-bp overlap of the end of dszA gene and the beginning of dszB gene is similar to the dsz operon of G. nitida (Park et al. 2003) and in contrast with the dszA-dszB junction of R. erythropolis IGTS8 which has four-base overlapping (Denis-Larose et al. 1998). The 13-bp gap between dszB and dszC genes is similar to G. nitida and R. erythropolis IGTS8. The structure of the dsz operon has shown to have serious effects on dsz gene expression (Li et al. 2007, 2008). Li et al. (2007) achieved a 12-fold increase in desulfurization activity by the rearrangement of the dszABC operon into dszBCA and removing the gene overlap within the operon and reconstructing the ribosome-binding site of dszB gene. The engineered operon was expressed from the plasmid vector pRHK1 in R. erythropolis 4.1491 (Li et al. 2007).
Several researchers working with Rhodococcus-desulfurizing strains reported that DBT-desulfurizing genes of the strains are located on large megabase pair plasmids (Monticello 2000; Gray et al. 2003). We have investigated the location of G. alkanivorans RIPI90A dsz operon using plasmid curing and pulsed field gel electrophoresis followed by Southern blot analysis (data not shown). Plasmid curing via heat treatment of the strain has shown no impact on loss of biodesulfurization trait; furthermore, the pulsed field gel electrophoresis and Southern blot analysis results indicate that, unlike the Rhodococcus dsz operon, the dsz operon of G. alkanivorans RIPI90A is located on bacterial chromosome. These results are in accordance to that of Santos et al. (2006) who reported that genes involved in DBT desulfurization by Gordonia sp. strain F.5.25.8 are located on the chromosome.
To isolate the G. alkanivorans RIPI90A dsz promoter, adaptor-ligated libraries were constructed by digestion of genomic DNA with EcoRI and PstI restriction enzymes and ligation to the specially designed adaptors, and the dsz promoter was PCR-amplified using gene-specific and adaptor-specific primers. The sequence similarity analysis of the 350-bp dszA upstream fragment using BLAST tool revealed a transposase-like sequence similar to several other desulfurization competent bacterial species. This transposase immediately near to the dsz operon could be responsible for horizontal transfer of the desulfurization operon between bacterial strains and its transfer from plasmid to the chromosome.
We used deletion analysis to delineate the sequences critical for promoter activity and determine the minimal promoter sequence. The deletion of first 180 bp (−335 to −156) of the G. alkanivorans RIPI90A promoter fragment has no consequence on the promoter activity and may be not containing any part of the dsz promoter; these results are in accordance to the findings of Li et al. (1996) who reported that deletion of first 145 bp from the 5′ end of the promoter containing fragment (regions −385 to −240) of dsz operon of R. erythropolis IGTS8 has no detectable effect on dsz promoter activity. As previously mentioned, this region contains a transposase-like sequence, and the presence of the transposase sequence at this region could be explanation for the ineffectiveness of deletion of the region on promoter activity.
Deletions of sequences farther than −156 dramatically decrease the promoter activity, and the activity has been completely abolished when region from −75 to −51 was eliminated. These results are comparable to the findings of Li et al. (1996) who has shown that the deletion of region from −160 to −80 eliminates the dsz promoter activity of the R. erythropolis IGTS8. From these results, we conclude that the sequence from −156 to −50 is necessary for full P-dsz activity and the dsz promoter localized at this region.
In summary, we have isolated and sequenced the dszABC genes and its regulatory region from DBT-desulfurizing strain, G. alkanivorans RIPI90A. In contrast to the dsz operon of R. erythropolis IGTS8 which is located on a large megaplasmid, the G. alkanivorans RIPI90A operon is located on bacterial chromosome. Despite the remarkably high homology between dsz genes of RIPI90A and R. erythropolis IGST8, promoter sequences of the strains were not very similar. We have utilized luciferase reporter gene and deletion analysis to localize and determine the minimal promoter sequence. The results indicate that the location of the dsz promoter of G. alkanivorans RIPI90A is somehow similar to R. erythropolis IGST8 and regions from −156 to −50 are responsible for full promoter activity.
References
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25:3389–3402
Alves L, Melo M, Mendonca D, Simoes F, Matos J, Tenreiro R, Girio FM (2007) Sequencing, cloning and expression of the dsz genes required for dibenzothiophene sulfone desulfurization from Gordonia alkanivorans strain 1B. Enzyme Microb Technol 40:1598–1603
Arenskötter M, Baumeister D, Kalscheuer R, Steinbüchel A (2003) Identification and application of plasmids suitable for transfer of foreign DNA to members of the genus Gordonia. Appl Environ Microbiol 69:4971–4974
Caro A, Boltes K, Calvo LP (2007) Dibenzothiophene biodesulfurization in resting cell conditions by aerobic bacteria. Biochem Eng J 35:191–197
Chen H, Zhang W, Cai Y, Zhang Y, Li W (2008) Elucidation of 2-hydroxybiphenyl effect on dibenzothiophene desulfurization by Microbacterium sp. strain ZD-M2. Bioresour Technol 99:6928–6933
Denis-Larose C, Bergeron H, Labbe D, Greer CW, Hawari J, Grossman MJ, Sankey BM, Lau PCK (1998) Characterization of the basic replicon of Rhodococcus plasmid pSOX and development of a Rhodococcus–Escherichia coli shuttle vector. Appl Environ Microbiol 64:4363–4367
Denome SA, Oldfield C, Nash LJ, Young KD (1994) Characterization of the desulfurization genes from Rhodococcus sp. strain IGTS8. J Bacteriol 176:6707–6716
Emamzadeh A, Hosseinkhani S, Sadeghizadeh M, Nikkhah M, Chaichi MJ, Mortazavi M (2006) cDNA cloning, expression and homology modeling of luciferase from the firefly Lampyroidea maculata. J Biochem Mol Biolo 39:578–585
Gallardo ME, Ferrandez A, Lorenzo VD, Garcia JL, Diaz E (1997) Designing recombinant Pseudomonas strains to enhance biodesulfurization. J Bacteriol 179:7156–7160
Gray KA, Mrachko GT, Squires CH (2003) Biodesulfurization of fossil fuels. Curr Opin Microbiol 6:229–235
Gupta N, Roychoudhury PK, Deb JK (2005) Biotechnology of desulfurization of diesel: prospects and challenges. Appl Microbiol Biotechnol 66:356–366
Hirasawa K, Ishii Y, Kobayashi M, Koizumi K (2001) Improvement of desulfurizing activity in Rhodococcus erythropolis KA2-5-1 by genetic engineering. Biosci Biotechnol Biochem 65:239–246
Ishii Y, Konishi J, Okada H, Hirasawa K, Onaka T, Suzuki M (2000) Operon structure and functional analysis of the genes encoding thermophilic desulfurizing enzymes of Paenibacillus sp. A11-2. Biochem Biophys Res Commun 270:81–88
Kilbane JJ (2006) Microbial biocatalyst development to upgrade fossil fuels. Curr Opin Biotechnol 17:305–314
Kilbane JJ, Robbins J (2007) Characterization of the dszABC genes of Gordonia amicalis F.5.25.8 and identification of conserved protein and DNA sequences. Appl Microbiol Biotechnol 75:843–851
Li GQ, Ting MA, Li SS, Li H, Liang F, Liu R (2007) Improvement of dibenzothiophene biodesulfurization activity by removing the gene overlap in the dsz operon. Biosci Biotechnol Biochem 71:849–854
Li GQ, Li SS, Li H, Zhang ML, Wang J, Zhu L, Liang F, Liu R, Ting MA (2008) Genetic rearrangement strategy for optimizing the dibenzothiophene biodesulfurization pathway in Rhodococcus erythropolis. Appl Environ Microbiol 74:971–976
Li ZM, Charles HS, Monticello DJ, Childs JD (1996) Genetic analysis of the dsz promoter and associated regulatory regions of Rhodococcus erythropolis IGTS8. J Bacteriol 178:6409–6418
Matsui T, Saeki H, Shinzato N, Matsuda H (2007) Analysis of the 7.6-kb cryptic plasmid pNC500 from Rhodococcus rhodochrous B-276 and construction of Rhodococcus–E. coli shuttle vector. Appl Microbiol Biotechnol 74:169–175
McFarland BL (1999) Biodesulfurization. Curr Opin Microbiol 2:257–264
Mohebali G, Ball AS (2008) Biocatalytic desulfurization (BDS) of petrodiesel fuels. Microbiology 154:2169–2183
Mohebali G, Ball AS, Rasekh B, Keytash A (2007) Biodesulfurization potential of a newly isolated bacterium, Gordonia alkanivorans RIPI90A. Enzyme Microb Technol 40:578–584
Monticello DJ (2000) Biodesulfurization and the upgrading of petroleum distillates. Curr Opin Biotechnol 11:540–546
Oldfield C, Pogrebinsky O, Simmonds J, Olson ES, Kulpa CF (1997) Elucidation of the metabolic pathway for dibenzothiophene desulphurization by Rhodococcus sp. strain IGTS8 (ATCC 53968). Microbiology 143:2961–2973
Park SJ, Lee I, Chang YK, Lee SY (2003) Desulfurization of dibenzothiophene and diesel oil by metabolically engineered E. coli. J Microbial Biotechnol 13:578–583
Piddington CS, Kovacevich BR, Rambosek J (1995) Sequence and molecular characterization of a DNA region encoding the dibenzothiophene desulfurization operon of Rhodococcus sp. strain IGTS8. Appl Environ Microbiol 61:468–475
Rishi AS, Nelson ND, Goyal A (2004) Genome walking of large fragments: an improved method. J Biotech 111:9–15
Sambrook J, Russell DW (2001) Molecular cloning, a laboratory manual, 3rd edn. Cold Spring Harbor Laboratory, Cold Spring Harbor
Santos SCC, Alviano DS, Alviano CS, Pádula M, Leitao AC, Martins OB, Ribeiro CMS, Sassaki MYM, Matta CPS, Bevilaqua J, Sebastián GV, Seldin L (2006) Characterization of Gordonia sp. strain F.5.25.8 capable of dibenzothiophene desulfurization and carbazole utilization. Appl Microbiol Biotechnol 71:355–362
Shavandi M, Sadeghizadeh M, Zomorodipour A, Khajeh K (2009) Biodesulfurization of dibenzothiophene by recombinant Gordonia alkanivorans RIPI90A. Bioresour Technol 100:475–479
Soleimani M, Bassi A, Margaritis A (2007) Biodesulfurization of refractory organic sulfur compounds in fossil fuels. Biotechnol Adv 25:570–596
Vasicova P, Patek M, Nesvera J, Sahm H, Eikmanns B (1999) Analysis of the Corynebacterium glutamicum dapA promoter. J Bacteriol 181:6188–6191
Yu B, Xu P, Zhu S, Cai X, Wang Y, Li L, Li F, Liu X, Ma C (2006) Selective biodegradation of S and N heterocycles by a recombinant Rhodococcus erythropolis strain containing carbazole dioxygenase. Appl Environ Microbiol 72:2235–2238
Acknowledgments
This work was supported by a fund from Iranian Research Institute of Petroleum Industry (R.I.P.I.). We gratefully thank M. Takeo for kindly donating the pRSG43 plasmid. The authors are grateful to Dr. S. Hoseinkhani for providing us with luciferase reporter gene and members of his laboratory for technical assistance in luciferase activity measurement.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Shavandi, M., Sadeghizadeh, M., Khajeh, K. et al. Genomic structure and promoter analysis of the dsz operon for dibenzothiophene biodesulfurization from Gordonia alkanivorans RIPI90A. Appl Microbiol Biotechnol 87, 1455–1461 (2010). https://doi.org/10.1007/s00253-010-2605-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-010-2605-4