
Abstract
The essential general features required for lipid membranes of extremophilic archaea to fulfill biological functions are that they are in the liquid crystalline phase and have extremely low permeability of solutes that is much less temperature sensitive due to a lack of lipid-phase transition and highly branched isoprenoid chains. Many accumulated data indicate that the organism’s response to extremely low pH is the opposite of that to high temperature. The high temperature adaptation does not require the tetraether lipids, while the adaptation of thermophiles to acidic environment requires the tetraether polar lipids. The presence of cyclopentane rings and the role of polar heads are not so straightforward regarding the correlations between fluidity and permeability of the lipid membrane. Due to the unique lipid structures and properties of archaeal lipids, they are a valuable resource in the development of novel biotechnological processes. This microreview focuses primarily on structural and physicochemical properties of polar lipids of (hyper)thermophilic archaea.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Classification of archaea
Archaea are regarded as one of the three main domains of life, its concept being proposed on the basis of phylogenetic analysis of ribosomal RNA (rRNA) sequences (Woese et al. 1990). This classification has been confirmed by extensive phylogenetic studies on a variety of macromolecules found in the patrimony of these microbes. Recently, the concept that Archaea and Bacteria were differentiated based on their membrane lipids has been advocated (Koga et al. 1998; Peretó et al. 2004; Wachtershauser 2003; William and Michael 2003). Archaeal domain, based on the evolutionary relationships, is divided into two phyla, the Euryarchaeota (from the Greek “euryos” meaning diversity) and the Crenarchaeota (from the Greek “crenos” meaning spring or origin; Fox et al. 1980; Woese and Fox 1977; Woese et al. 1990; Woese 1987). With the rise of the molecular ecology and the discovery of many novel lineages of mesophilic and psychrophilic archaea, a new, third archaeal phylum Thaumarchaeota (from the Greek ‘thaumas’, meaning wonder) was proposed (Brochier-Armanet et al. 2008).
Cultivated archaea are divided, based on their physiological and metabolic characteristics, into six groups. The first group includes extremely halophilic archaea that thrive in environments with salt concentrations up to 200 g/l. In the second group are those extremely thermophilic archaea that grow optimally at 80ºC or more, with a current record of growth at 121ºC (Kashefi and Lovley 2003). The third group is comprised of acidophilic archaea that thrive in acidic environment often also at elevated temperature. To maintain a near neutral pH, these organisms must withstand proton gradients up to five orders of magnitude (Macalady et al. 2004). The fourth group includes ammonia-oxidizing archaea or nitrifiers that are abundant and important in the open ocean (Wuchter et al. 2006) and in soils (Leininger et al. 2006). They mostly inhabit surficial sediments that are likely to have low concentrations of O2, and the open ocean, in which ammonia concentrations are typically an order or more lower than in more replete environments. The fifth group includes methanogens, which are a group of strictly anaerobic archaea characterized by the unique ability to produce methane. These organisms thrive across a broad range of temperature, salinity, and pH. The sixth group is composed of anaerobic methane oxidizers that are closely related to the methanogens and are thought to form syntropic associations with sulfate- or nitrate- reducing bacteria (Raghoebarsing et al. 2006).
Special biochemical adaptations of archaea
Archaea and Bacteria differ in many genetic and biochemical properties. One important distinction between Archaea and Bacteria is the chemical structure of lipids composing the cytoplasmic membrane, which will be discussed later in the chapter. Valentine (2007) hypothesized in his article that the adaptation to chronic energy stress is the crucial factor that distinguishes the Archaea from the Bacteria and that the primary biochemical basis for this adaptation are low-permeability membranes and specific catabolic pathways. Catabolic specificity of Archaea (aerobiosis, fermentation, nitrification, methanogenesis, phototrophy, and numerous forms of lithotrophy) could be interpreted in terms of adaptation to chronic energy stress, conditions in which cells are consistently faced with an insufficient supply of cellular energy (high rates of futile ion cycling driven by extreme temperature, acidity or salinity, as well as low rates of cellular energy generation due to limited substrate availability and/or unfavorable thermodynamic conditions; Valentine 2007). From a metabolic perspective in competition between Archaea and Bacteria, the dominance of Archaea is achieved through several successful modes. Halophiles, hyperthermophiles, and thermoacidophiles are groups adapted to cope with a level of energy stress that disables all but few selected bacterial species. As conditions become more moderate, this environmental exclusivity gives way to metabolic exclusivity, which is in the case of methanogens and methane oxidizers. Hence, their dominance is achieved with exclusion of Bacteria by the use of unique catabolic pathways. In moderate environmental conditions, the level of competition between archaea and bacteria intensifies and successful archaea maintain a singularity of catabolism. This allows them to exclude or out-compete bacteria in specific niches with low energy availability. Methanogens, methane oxidizers, and possibly nitrifiers each seemingly display such singularity, with their catabolic emphasis on a single well-defined pathway (Valentine 2007).
Archaeal lipids
The molecular adaptations responsible for archaeal ability to survive and grow in harsh environments have clearly emphasized the key role of membrane lipid components in overcoming the destabilizing conditions encountered in such extreme environments as hot acidic springs and submarine volcanic fields. Since the first archaeal diether polar lipids were discovered in 1960s, more than 100 new lipids have been found in various archaea with great variety of structures.
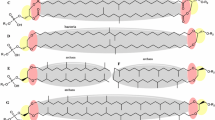
Polar archaeal lipids are generally composed of a core lipid and a phosphodiester bonded polar head groups or glycosides that are linked to one of the core lipids. Profiles of archaeal lipid core are quite remarkable in their own right even though they are not as diverse as those found in Bacteria and Eukarya. However, four characteristics unique to archaeal lipids compared with bacterial lipids have been identified (Fig. 1) (Koga and Morii 2006). (1) Glycerophosphate backbone of archaeal lipids is composed of sn-glycerol-1-phosphate (G-1-P), which is an enantiomer of sn-glycerol-3-phosphate (G-3-P) in bacterial phospholipids. Hydrocarbon chains are though bound at the sn-2 and -3 positions of the glycerol moiety in archaeal lipids, whereas bacterial lipids have sn-1,2-radyl chains. (2) Hydrocarbon chains are bonded to the glycerol moiety exclusively by ether linkages in archaeal polar lipids in contrast to bacterial lipids, most of which have ester linkages. (3) The hydrocarbon chains of polar lipids in Archaea are highly methyl-branched isoprenoids and isopranoids. Bacterial counterparts are mostly straight chain fatty acids. (4) Significant number of archaeal species contain bipolar lipids with a tetraether core that span through membranes (bolaform amphiphilic), which are rarely found in Bacteria.

Typical structures of archaeal phospholipid compared with bacterial phospholipid. Four differences are shown. –R stands for polar head group
Core lipid structure
Basically, major core lipid structures consist of the ubiquitous standard archaeol (2,3-di-O-diphytanil-sn-glicerol) consisting mainly of C20,20 alkyl chains and/or the standard caldarchaeol (2,2′,3,3′-tetra-O-dibiphytanil-sn-diglycerol) with C40,40 carbon chains (Fig. 2). Variations to these standard lipid cores include archaeols with increased number of C5 units or caldarchaeols with cyclopentane rings within the C40-phytanil chains (Fig. 2; Patel and Sprott 1999). Recently, a numerous new core lipid structures have been elucidated (Koga and Morii 2005). Those include isocaldarchaeol from Methanothermobacter marburgensis (mixture of regioisomers with antiparallel and parallel arrangements of two glycerol moieties), H-shaped caldarchaeol from Methanothermus fervidus (Morii et al. 1998), C40 isoprenoid hydrocarbons with cyclopentane rings at unusual positions isolated from sedimentary organic matter samples of water column from several marine environments (Hoefs et al. 1997), macrocyclic archaeol with one or two cyclopentane rings in the byphytanediyl chain (Stadnitskaia et al. 2002), alkyl ether type core lipids from Methanopyrus kandleri and Methanococcoides burtonii (Sprott et al. 1997a), and many more new lipids that are minor modifications of known lipids of archaea (Koga and Morii 2005).

Typical archaeal polar lipid cores. a Archaeol (C20,20), b archaeol (C25,25), c macrocyclic arhaeol, d caldarchaeol, e isocaldarchaeol, f crenarchaeol, g H-shaped caldarchaeol containing three cyclopentane rings
Polar head groups
The polar head groups of archaeal lipids are phospholipids or glycosides that are linked to one of the core lipids. The most common phospholipids are phosphoserin, phosphoinositol, phosphoglycerol, phosphoetanolamin, and many phosphoglyicolipids; among them, the most common carbohydrates found among archaeal lipids are glucose, gulose, mannose, galactose, inositol, and N-acetylglucosamine, which can form mono-, di-, or oligosaccharides on one or both sides of caldarchaeol. Phosphoglycolipids with two polar head groups on both sides of the caldarchaeol may have glycerophosphate as the phosphoester moiety on one side and gulose alone or glucose and mannose, which form mono-, di-, or oligosaharides as the sugar moiety on the other side as is in the case of Thermoplasma acidophilum (Shimada et al. 2008). Replacement of one glycerol moiety of the core lipid backbone by a nonitol has also been observed (De Rosa and Gambacorta 1988). The occurrence of unusual carbohydrate ß-d-galactofuranosyl units has been found in methanogens (Gambacorta et al. 1995), but it has not been found in thermoacidophiles (Tables 1 and 2), since the five-member rings in such environments are rapidly hydrolyzed.
Biosynthesis of the polar lipids in thermophilic archea
An excellent review, which focused on in vitro studies of archaeal lipid biosynthesis, has been published by Koga and Morii (2007). Here we present the brief overview of the biosynthesis of the main compounds of archaeal dieter lipids.
-
1.
The hydrocarbon portions of archaeal dieter lipids are exclusively isoprenoids (C20 phytanyl, C25 sesterterpenyl, or farnesylgeranyl groups). The isoprenoid biosynthetic pathway is divided into two sections. The first half is the synthesis from acetyl-coenzyme A (CoA) to isopentyl diphosphate (IPP) and dimethylallyl diphosphate (DMAPP), and the second half is the synthesis of polyprenyl diphosphate from the two C5 units (IPP and DMAPP; Fig. 3). For IPP synthesis, two independent pathways are known: the classical mevalonate (MVA) pathway and more recently discovered MVA-independent (1-deoxy-d-xylulose-5-phosphate pathway; Koga and Morii 2007 and reference cited therein). The archaeal MVA pathway is a mosaic composed of the enzymes common to Archaea and Eucarya, along with the enzymes unique to Archea and Bacteria.
Fig. 3 
MVA pathway for synthesis of isopentenyl diposphate (IPP) and dimethylallyl diposphatehyl (DMAPP) and further formation of geranylgeranyl-PP (GGPP) (Koga and Morii 2007)
-
2.
G-1-P is the enantiomer of bacterial G-3-P and it is formed from d-glycerolaldehyde-3-phosphate (D-GAP) by three possible reactions. The first possible reaction for G-1-P formation is the reduction of GAP at the position 1. Because D-GAP has the same configuration as G-1-P at the position 2, the simple reduction of its aldehyde group can form G-1-P. The second possibility for the G-1-P formation is the reduction of dihydroxyacetonephosphate at the position 2 catalyzed by G-1-P dehydrogenase, the enzyme responsible for the formation of enantiomeric glycerophosphate backbone structure in archaeal phospholipids. The third possibility for G-1-P formation is the direct phosphorylation of glycerol at the sn-1 position by adenosine triphosphate (Koga and Morii 2007 and reference cited therein).
-
3.
The pathway for biosynthesis of the ether-type phospholipids in Archaea is depicted in Fig. 4. The results of in vivo studies suggested that the ether bond is formed from unsaturated prenyl precursors. The bond formation is catalyzed by two prenyl transferases: One is responsible for formation of the first ether bond between the sn-3 hydroxyl group of G-1-P and geranylgeranyl diphosphate (GGPP; GGGP synthase), and the other catalyzes the formation of ether bond at the sn-2 position to form di-geranylgeranyl-G-1-P (DGGGP or unsaturated archaetidic acid; DGGGP synthase). For details, see the recent review published by Koga and Morii (2007).
Fig. 4 
The proposed biosynthetic pathway for phospholipids synthesis in archaea (Koga and Morii 2007). Enzymes confirmed by in vitro experiments are as follows: 1 G-1-P dehydrogenase; 2 GGGP synthase; 3 DGGGP synthase; 4 CDP-archaeol synthase
-
4.
The final product of thermophilic archaea are fully saturated chains. Hydrogenation or saturation reaction may take place somewhere after attachment of polar head groups in the presence of reductase, which requires nicotinamide adenine dinucleotide (reduced form) as a hydrogen donor via flavin adenine dinucleotide (oxidized). If unsaturated archaetidic acid were saturated, it could not react with CDP-archaeol synthase, which reacts preferentially with unsaturated archaetidic acid.


Physicochemical properties of archaeal lipids
The existence of such a large variety of unusual lipid structures in Archaea raises a question of how these lipids function in membranes of these organisms. Ether linkages are more stable than ester over a wide range of pH, and the branching methyl groups help to reduce both crystallization (membrane lipids in the liquid crystalline phase at ambient temperature) and membrane permeability (steric hindrance of the methyl side group). The saturated alkyl chains would impair stability toward oxidative degradation particularly in halophiles that are exposed to air and sunlight (Benvegnu et al. 2008). The membranes of methanogens and thermoacidophiles essentially consist of bipolar monolayer structures (Table 2). The high proportions of glycosylated lipids presented in membranes of thermoacidophiles and methanogenes may further stabilize their membrane structures through the interglycosyl headgroup hydrogen bonding. The presence of large sugar heads toward the convex surface of the membrane is likely to promote an asymmetric orientation, thus making the monolayer organization easier. Furthermore, the flux of small molecules and protons through archaeal bipolar tetraether lipid membranes is considerably reduced as the result of the particular physical structure of the lipid monolayer. Finally, the presence of the cyclic diether structures in species isolated from deep-sea hydrothermal vents may be related to the high pressures under which these archaea live.
Thermal stability
The primary and fundamental physiological function of polar lipids is to form a cell membrane, which encloses a cell and makes a permeability barrier for various essential solutes for life. One of the essential general features required for lipid membrane to fulfill biological functions is that they are in the liquid crystalline phase. Archaeal lipid membranes have, in general, much lower phase transition temperature than fatty acyl ester lipids (Yamauchi et al. 1993). While membranes made of fatty acyl ester lipids are in the gel phase or in the liquid crystalline phase depending mostly on their fatty acid composition, archaeol- and caldarchaeol-based polar lipid membranes of archaea are assumed to be in the liquid crystalline phase at a wide temperature range of 0–100°C. Recently, it has been shown in vivo by applying the electron paramagnetic resonance and fluorescence emission spectroscopy that the membrane fluidity of hyperthermophilic archaeon Aeropyrum pernix consisted solely of 2,3-di-O-sesterterpanyl-sn-glycerol core lipid (C25,25-archaeol; Morii et al. 1999), gradually increases with the temperature up to 65°C, the lower temperature limit of A. pernix growth (Ulrih et al. 2007). Similar results were observed for in vitro study on the total polar lipids (TPL) isolated from thermophilic archaeon Sulfolobus acidocaldarius, which become fluid at temperature close to minimum growth temperature (~50°C) of thermoacidophilic archaeon (Khan and Chong 2000). Additionally, differential dynamic calorimetry (DSC) and pressure perturbation calorimetry (PPC) were used to characterize thermal phase transition, membrane packing, and volumetric properties of multilamellar vesicles prepared from TPL isolated from S. acidocaldarius (Chong et al. 2005). In the temperature range at ~47–50°C and ~69°C, two phase transitions were detected by DSC, which involve very small volume changes, the first one is ascribed to lamellar-to-lamellar phase transition, while the second one with significant enthalpy changed to lamellar-to-cubic phase transition. These phase transitions are followed with very low volume changes, indicating that the polar headgroup region of TPL may still be rigid and tightly packed through hydrogen-bond network at elevated temperatures.
Permeability
The transport of small molecules across lipid bilayers is a fundamental biological process. Given the extreme environmental conditions in which these archaea thrive, their membranes have to fulfill two conditions required for a biological membrane, liquid crystalline phase and low permeability, with almost the same composition in a wide temperature range. It has been observed that archaeal cultures grown at different conditions exhibit different lipid membrane ratios as is the case of Archaeoglobus fulgidus, where change in tetraether/diether lipid ratio and number of pentacycles in the hydrocarbon chains was observed between cultures grown at 70°C or 89°C, respectively (Lai et al. 2008). Here, we focus on hyperthermophiles and acidothermophiles (Tables 1 and 2). The membrane of acidothermophiles is composed of 100% tetraether lipids, while for (neutro)-hyperthermophiles, the ratios between tetraether and diether lipids is strain dependent (Tables 1 and 2). In vitro study using the liposomes (archeosomes) made from caldarchaeol lipids of S. acidocaldarius, which has an optimal growth temperature of 85°C at pH 2.0, at temperatures below 40°C, no proton permeability was observed (Elferink et al. 1994). In the similar study of the major polar lipids from S. acidocaldarius, it was shown that membrane surface charge was responsible for low 5,6-carboxylfluorescence permeability but not for proton permeability (Elferink et al. 1994). More examples are listed in following paragraphs describing archaeosomes.
It is known that the structural differences between lipids affect solute permeation through the membranes. In the polar headgroup regions of bipolar tetraether liposomes, there is an extensive network of hydrogen bonds, which should generate a high electrical dipole potential, thus hindering solute permeability through membranes. Unlike non-archaeal monopolar diester/diether lipids, archaeal bipolar tetraether lipids do not have the hydrocarbon terminal methyl groups; instead, the biphytanyl hydrocarbon chains are linked covalently from one polar end to the other, lacking the midplace spacing. Furthermore, there are cyclopentane rings in the phytanyl chains, which provide additional rigidity to the membrane. Because of the stiffness in the molecule, bipolar tetraether lipids should exhibit rather limited gauche-trans isomerization in their hydrocarbon chains. Conceivably, solutes cannot easily move in the hydrocarbon region of the bipolar membrane via “hopping.” From the viewpoints of penetration and “hopping” solute permeation across archaeal bipolar tetraether lipid membranes should be less than that for archaeal dieter lipids and non-archaeal monopolar diester or dieter lipid membrane (Gliozzi et al. 2002 and reference cited therein).
Artificial archaeal lipid membranes—archaeosomes
Archaeal lipids were shown to be an excellent source for the formation of liposomes (archaeosomes) with remarkable thermostability and tightness against solute leakage (Gambacorta et al. 1995). Ether lipids are also resistant to enzymatic degradation by phospholipases, and archaeal liposomes are exceptionally stable; they do not fuse or aggregate during storage at 4°C over a period of 4 months (De Rosa 1996). Membrane stability can be conveniently monitored by determining the release of fluorescent dyes originally trapped in the intravesicular compartment of the liposomes. In particular, several studies have focused on investigation of membranes made solely from bipolar lipid fractions. Different physicochemical properties (structure, dynamics, and polymorphism and thermal and mechanical stability) of bipolar lipid fractions extracted from several archaeal species have been investigated. However, there are different views on the specific structural factors responsible for the high thermostability. Chang and Elferink (Chang 1994; Elferink et al. 1994) suggested that the high thermostability is not due to the phytanyl chain alone, in contrast to the perception of Yamauchi and his coworkers (Yamauchi et al. 1993).
Arakawa et al. (1999) investigated the polymorphism and physicochemical properties of the macrocyclic lipids by synthetic 72-membered macrocyclic tetraether lipids. DSC, 31P NMR and electron microscopy analysis studied the physicochemical features of diphospholipids. The cyclic tetraetherlipids appeared to show lower phase transition temperature (T c). Fluorescence studies have shown that the passive proton permeability in bipolar tetraether liposomes isolated from S. acidocaldarius is lower and less temperature sensitive than that in liposomes composed of monopolar diester lipids, although the permeability increases with temperature in all liposomes. It has been proposed that low proton permeability is due to the chemical structure of tetraether lipids and their monolayer organization, especially the cyclopentane rings and the network of hydrogen bonds between the sugar residues exposed at the outer face of the membrane (Chong et al. 2003, Mathai et al. 2001).
In general, it was shown that, at a given temperature, the bolaform lipid chains are more ordered and less flexible than in conventional bilayer membranes. Only at elevated temperatures (80°C) does the flexibility of the chain environment in tetraether lipid assemblies approach that of fluid bilayer membranes (Bartucci et al. 2005). By examination of water, solute (urea and glycerol), proton, and ammonia permeability of archaeosome, it was shown that macrocyclic archaeol and caldarchaeol lipids reduced the water, ammonia, urea, and glycerol permeability significantly (6–120-fold) compared to dipalmitoylphosphatidylcholine (DPPC) liposomes (Mathai et al. 2001). The presence of an ether bond reduced the apparent proton permeability for threefold compared to DPPC liposomes. The presence of the ether bond and phytanyl chains did not significantly affect these permeabilities. The presence of macrocycllic archaeol and caldarchaeol structure further reduced apparent proton permeability by 10–17-fold (Mathai et al. 2001). Komatsu and Chong have suggested that the low leakage rates of anionic probe at neutral pH are due to the negative charges of polar tetraether lipid membrane surface in addition to the tight and rigid lipid packing (Komatsu and Chong 1998). Some studies have shown that the leakage rate of entrapped dyes increases with decreasing the content of tetraether lipids and increases in the presence of fusogenic agents such as calcium ions and polyethylene glycol. These observations and many others suggest that the degree of hydration and conformation of the lipid polar headgroup are crucial for the tetraether membrane integrity. Furthermore, the release of entrapped dyes in bipolar tetraether liposomes is less temperature sensitive than that in non-archaeal monopolar diester liposomes (Mathai et al. 2001), suggesting a low thermal expansibility for tetraether liposomes (Chong et al. 2003). This general property is important for thermophiles, since they may experience temperature fluctuations in their native environment.
Biotechnological applications of archeosomes
Both in vitro and in vivo studies indicate that archaeosomes are safe. Their pH and thermal stability, tissue distribution profiles, as well as the adjuvant activity of archaeosome formulations indicate that they may offer a superior alternative for several biotechnological applications, including delivery systems for drugs, genes, or cancer imaging agents (Table 3).
As an ideal vector in drug and gene delivery should be highly efficient in delivering the drug in a target-specific manner (various molecules can be incorporated into archaeosomes or associated with the vesicles to target-specific tissues), stable in vitro as well as in vivo, nontoxic and non-immunogenic, archaeosomes delivery systems answer most of these conditions. The two important observations are (1) archaeosomes promote strong humoral and cell-mediated (Th1/Th2) immune reactions to soluble-entrapped proteins. More importantly, archaeosomes induce CD8+ cytotoxic T lymphocyte responses (critical for protection against cancer and intracellular infections) to entrapped proteins. Thus, archaeosomes are self-adjuvanting delivery vesicles (Sprott et al. 1997b; Sprott et al. 1999) and (2) better immunization with archaeosomes is achieved (Sprott et al. 2003). Recently, two excellent reviews were published covering the archaeosome adjuvants from immunological capabilities to mechanism(s) of action (Krishnan and Sprott 2008), and a synthetic route for the preparation of symmetrical and unsymmetrical archaeal tetraether-like analogs has been described for use in these purposes (Brard et al. 2007). Archaeal lipids have also been proposed as monomers for bioelectronics for several reasons, the most important being that their bipolar tetraether structure offers novel opportunities for protein–lipid interactions, which are of interest for the assembly of electronic devices based on redox proteins or enzymes (De Rosa et al. 1994). Some other applications include use of archaeal lipids as taxonomic markers (Gambacorta et al. 1995) and as novel lubricants for engines, since they are chemically stable at high temperatures, lubricate, either with or without a carrier lubricant, and provide a friction coefficient of less than 0.1 (Chang 1992).
Conclusions
The organism’s response to extremely low pH is the opposite of that to high temperature. The literature data indicate that the presence of bipolar tetraether lipids is not necessary for thermal adaptation. A significant number of hyperthermophilic archaea does not contain tetraether lipids at all (Tables 1 and 2). Some neutrophilic hyperthermophiles contain C25,25 diether lipids. The chain length of the C25-isoprenoid hydrocarbon is 20% longer than those of the C20-isoprenoid and C18 straight-chain fatty acids. Therefore, with it composed of only the C25,25-archaeol-based lipids, the thickness of the membrane of A. pernix is assumed to be 20% greater than the membranes of the C20,20-archaeol-based lipids of other archaea (Morii et al. 1999). Furthermore, the presence of variety of polar head groups in an archaeal membrane is not necessary for adaptation to thermal environments. The adaptation of thermophiles to acidic environments requires the presence of tetraether polar lipids in addition to polar heads to maintain the low permeability of the membrane (Tables 1 and 2). The higher temperature adaptation might or might not require the higher number of cyclopentane rings compared to low pH. The number of sugar lipids units of glyco(phospho)lipids increase under high temperature and low pH conditions. The sugar moiety of lipids may interact with each other via hydrogen bonding. The network with increased sugar units of polar lipids at low pHs may have decreased membrane fluidity. The number of cyclopentane rings may have decreased to increase membrane fluidity at low pH and to counteract the effect of increased amount of sugar moieties and vice versa at higher temperature. All this characteristics of unique lipid structures and properties of archaeal lipids are important when development of novel biotechnological processes/products or their application is considered. It is obvious that application will be further developed on the basis of our understanding of archaeal lipids functioning in natural environment, which will open new possibilities in new technologies such as biotechnology, nanotechnology, and also in farma and nutria applications.
References
Arakawa K, Kano H, Eguchi T, Nishiyama Y, Kakinuma K (1999) Significance of the 72-membered macrocyclic structure found in archaeal membrane lipids: model studies of the macrocyclic tetraether diphospholipids by calorimetric, P-31 NMR, and electron microscopic analyses. Bull Chem Soc Jpn 727:1575–1581
Bartucci R, Gambacorta A, Gliozzi A, Marsh D, Sportelli L (2005) Bipolar tetraether lipids: chain flexibility and membrane polarity gradients from spin-label electron spin resonance. Biochemistry 4445:15017–15023
Bauer S, Heckmann K, Six L, Strobl C, Blöcher D, Henkel B, Garbe T, Ring K (1983) Hyperfiltration through crosslinked monolayers II. Desalination 461–3:369–378
Benvegnu T, Lemiegre L, Cammas-Marion S (2008) Archaeal lipids: innovative materials for biotechnological applications. Eur J Org Chem 28:4725–4744
Berzina TS, Troitsky VI, Vakula S, Riccio A, Morana A, De Rosa M, Dante S, Maccioni E, Rustichelli F, Accossato P, Nicolini C (1995) Langmuir–Blodgett-films of bipolar lipids from thermophilic archaea. Mater Sci Eng C Biomim Mater Sens Syst 31:13–21
Berzina TS, Troitsky VI, Vakula S, Riccio A, De Rosa M, Nicolini C (1997) Surface potential study of selective interaction of potassium ions with valinomycin in Langmuir-Blodgett films. Mater Sci Eng C Biomim Mater Sens Syst 51:1–6
Blöchl E, Rachel R, Burggraf S, Hafenbradl D, Jannasch HW, Stetter KO (1997) Pyrolobus fumarii gen. and sp. nov., represents a novel group of archaea, extending the upper temperature limit for life to 113 °C. Extremophiles 11:14–21
Brard M, Laine C, Rethore G, Laurent I, Neveu C, Lemiegre L, Benvegnu T (2007) Synthesis of archaeal bipolar lipid analogues: a way to versatile drug/gene delivery systems. J Org Chem 7222:8267–8279
Brochier-Armanet C, Boussau B, Gribaldo S, Forterre P (2008) Mesophilic crenarchaeota: proposal for a third archaeal phylum, the Thaumarchaeota. Nat Rev Microbiol 63:245–252
Chang EL (1992) New class of lubricants derived from archaebacterial lipids. The United States of America as represented by the Secretary of the Navy. United States Patent 5,098,588
Chang EL (1994) Unusual thermal stability of liposomes made from bipolar tetraether lipids. Biochem Biophys Res Commun 2022:673–679
Chong PLG, Zein M, Khan TK, Winter R (2003) Structure and conformation of bipolar tetraether lipid membranes derived from thermoacidophilic archaeon Sulfolobus acidocaldarius as revealed by small-angle X-ray scattering and high-pressure FT-IR spectroscopy. J Phys Chem B 10733:8694–8700
Chong PLG, Ravindra R, Khurana M, English V, Winter R (2005) Pressure perturbation and differential scanning calorimetric studies of bipolar tetraether liposomes derived from the thermoacidophilic archaeon Sulfolobus acidocaldarius. Biophys J 893:1841–1849
Comita PB, Gagosian RB, Pang H, Costello CE (1984) Structural elucidation of a unique macrocyclic membrane lipid from a new, extremely thermophilic, deep-sea hydrothermal vent archaebacterium, Methanococcus jannaschii. J Biol Chem 25924:5234–5241
De Rosa M, Gambacorta A, Trincone A, Basso A, Zillig W, Holz I (1987) Lipids of Thermococcus celer, a sulfur-reducing archaebacterium—structure and biosynthesis. Syst Appl Microbiol 91–2:1–5
De Rosa M, Gambacorta A (1988) The lipids of archaebacteria. Prog Lipid Res 273:153–175
De Rosa M, Morana A, Riccio A, Gambacorta A, Trincone A, Incani O (1994) Lipids of the archaea: a new tool for bioelectronics. Biosens Bioelectron 99–10:669–675
De Rosa M (1996) Archaeal lipids: structural features and supramolecular organization. Thin Solid Films 284–285:13–17
Elferink MGL, de Wit JG, Driessen AJM, Konings WN (1994) Stability and proton-permeability of liposomes composed of archaeal tetraether lipids. Biochim Biophys Acta Biomembr 11932:247–254
Ellen A, Albers SV, Huibers W, Pitcher A, Hobel C, Schwarz H, Folea M, Schouten S, Boekema E, Poolman B, Driessen A (2009) Proteomic analysis of secreted membrane vesicles of archaeal Sulfolobus species reveals the presence of endosome sorting complex components. Extremophiles 131:67–79
Erauso G, Reysenbach AL, Godfroy A, Meunier JR, Crump B, Partensky F, Baross JA, Marteinsson V, Barbier G, Pace NR, Prieur D (1993) Pyrococcus abyssi sp-nov, a new hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent. Arch Microbiol 1605:338–349
Ferrante G, Richards JC, Sprott GD (1990) Structures of polar lipids from the thermophilic, deep-sea archaeobacterium Methanococcus jannaschii. Biochem Cell Biol 681:274–283
Fox GE, Stackebrandt E, Hespell RB, Gibson J, Maniloff J, Dyer TA, Wolfe RS, Balch WE, Tanner RS, Magrum LJ, Zablen LB, Blakemore R, Gupta R, Bonen L, Lewis BJ, Stahl DA, Luehrsen KR, Chen KN, Woese CR (1980) The phylogeny of prokaryotes. Science 2094455:457–463
Gambacorta A, Gliozzi A, De Rosa M (1995) Archaeal lipids and their biotechnological applications. World J Microbiol Biotechnol 111:115–131
Gliozzi A, Relini A, Chong PLG (2002) Structure and permeability properties of biomimetic membranes of bolaform archaeal tetraether lipids. J Membr Sci 2061:131–147
Godfroy A, Meunier JR, Guezennec J, Lesongeur F, Raguenes G, Rimbault A, Barbier G (1996) Thermococcus fumicolans sp. nov., a new hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent in the North Fiji Basin. Int J Syst Bacteriol 464:1113–1119
Hensel R, Matussek K, Michalke K, Tacke L, Tindall BJ, Kohlhoff M, Siebers B, Dielenschneider J (1997) Sulfophobococcus zilligii gen. nov., spec. nov. a novel hyperthermophilic archaeum isolated from hot alkaline springs of Iceland. Syst Appl Microbiol 201:102–110
Hoefs MJL, Schouten S, De Leeuw JW, King LL, Wakeham SG, Damste JSS (1997) Ether lipids of planktonic archaea in the marine water column. Appl Environ Microbiol 638:3090–3095
Huber R, Dyba D, Huber H, Burggraf S, Rachel R (1998) Sulfur-inhibited Thermosphaera aggregans sp. nov., a new genus of hyperthermophilic archaea isolated after its prediction from environmentally derived 16S rRNA sequences. Int J Syst Bacteriol 48:31–38
Huber H, Burggraf S, Mayer T, Wyschkony I, Rachel R, Stetter KO (2000) Ignicoccus gen. nov., a novel genus of hyperthermophilic, chemolithoautotrophic archaea, represented by two new species, Ignicoccus islandicus sp. nov. and Ignicoccus pacificus sp. nov. Int J Syst Evol Microbiol 506:2093–2100
Itoh T, Suzuki K, Nakase T (1998) Thermocladium modestius gen. nov., sp. nov., a new genus of rod-shaped, extremely thermophilic crenarchaeote. Int J Syst Bacteriol 48:879–887
Itoh T, Suzuki K, Sanchez PC, Nakase T (1999) Caldivirga maquilingensis gen. nov., sp. nov., a new genus of rod-shaped crenarchaeote isolated from a hot spring in the Philippines. Int J Syst Bacteriol 493:1157–1163
Itoh Y, Kurosawa N, Uda I, Sugai A, Tanoue S, Itoh T, Horiuchi T, Itoh T (2001) Metallosphaera sedula TA-2, a calditoglycerocaldarchaeol deletion strain of a thermoacidophilic archaeon. Extremophiles 54:241–245
Itoh T, Suzuki K, Sanchez PC, Nakase T (2003) Caldisphaera lagunensis gen. nov., sp. nov., a novel thermoacidophilic crenarchaeote isolated from a hot spring at Mt Maquiling, Philippines. Int J Syst Evol Microbiol 534:1149–1154
Itoh T, Yoshikawa N, Takashina T (2007) Thermogymnomonas acidicola gen. nov., sp. nov., a novel thermoacidophilic, cell wall-less archaeon in the order Thermoplasmatales, isolated from a solfataric soil in Hakone, Japan. Int J Syst Evol Microbiol 5711:2557–2561
Jahn U, Summons R, Sturt H, Grosjean E, Huber H (2004) Composition of the lipids of Nanoarchaeum equitans and their origin from its host Ignicoccus sp. strain KIN4/I. Arch Microbiol 1825:404–413
Kashefi K, Lovley DR (2003) Extending the upper temperature limit for life. Science 3015635:934
Khan TK, Chong PLG (2000) Studies of archaebacterial bipolar tetraether liposomes by perylene fluorescence. Biophys J 783:1390–1399
Koga Y, Kyuragi T, Nishihara M, Sone N (1998) Did archaeal and bacterial cells arise independently from noncellular precursors? A hypothesis stating that the advent of membrane phospholipid with enantiomeric glycerophosphate backbones caused the separation of the two lines of descent. J Mol Evol 461:54–63
Koga Y, Morii H (2005) Recent advances in structural research on ether lipids from archaea including comparative and physiological aspects. Biosci Biotechnol Biochem 6911:2019–2034
Koga Y, Morii H (2006) Special methods for the analysis of ether lipid structure and metabolism in archaea. Anal Biochem 3481:1–14
Koga Y, Morii H (2007) Biosynthesis of ether-type polar lipids in archaea and evolutionary considerations. Microbiol Mol Biol Rev 711:97–120
Komatsu H, Chong PLG (1998) Low permeability of liposomal membranes composed of bipolar tetraether lipids from thermoacidophilic archaebacterium Sulfolobus acidocaldarius. Biochemistry 371:107–115
Krishnan L, Sprott GD (2008) Archaeosome adjuvants: immunological capabilities and mechanism(s) of action. Vaccine 2617:2043–2055
Kurosawa N, Itoh YH, Iwai T, Sugai A, Uda I, Kimura N, Horiuchi T, Itoh T (1998) Sulfurisphaera ohwakuensis gen. nov., sp. nov., a novel extremely thermophilic acidophile of the order Sulfolobales. Int J Syst Bacteriol 48:451–456
Lai D, Springstead J, Monbouquette H (2008) Effect of growth temperature on ether lipid biochemistry in Archaeoglobus fulgidus. Extremophiles 122:271–278
Lanzotti V, De Rosa M, Trincone A, Basso AL, Gambacorta A, Zillig W (1987) Complex lipids from Desulfurococcus mobilis, a sulfur-reducing archaebacterium. Biochim Biophys Acta 9222:95–102
Lanzotti V, Trincone A, Nicolaus B, Zillig W, De Rosa M, Gambacorta A (1989) Complex lipids of Pyrococcus and AN1, thermophilic members of archaebcteria belonging to Thermococcales. Biochim Biophys Acta 10041:44–48
Lattuati A, Guezennec J, Metzger P, Largeau C (1998) Lipids of Thermococcus hydrothermalis, an archaea isolated from a deep-sea hydrothermal vent. Lipids 333:319–326
Leininger S, Urich T, Schloter M, Schwark L, Qi J, Nicol GW, Prosser JI, Schuster SC, Schleper C (2006) Archaea predominate among ammonia-oxidizing prokaryotes in soils. Nature 4427104:806–809
Macalady JL, Vestling MM, Baumler D, Boekelheide N, Kaspar CW, Banfield JF (2004) Tetraether-linked membrane monolayers in Ferroplasma spp a key to survival in acid. Extremophiles 85:411–419
Marteinsson VT, Birrien JL, Reysenbach AL, Vernet M, Marie D, Gambacorta A, Messner P, Sleytr UB, Prieur D (1999) Thermococcus barophilus sp. nov., a new barophilic and hyperthermophilic archaeon isolated under high hydrostatic pressure from a deep-sea hydrothermal vent. Int J Syst Bacteriol 492:351–359
Mathai JC, Sprott GD, Zeidel ML (2001) Molecular mechanisms of water and solute transport across archaebacterial lipid membranes. J Biol Chem 27629:27266–27271
Matsuno Y, Sugai A, Higashibata H, Fukuda W, Ueda K, Uda I, Sato I, Itoh T, Imanaka T, Fujiwara S (2009) Effect of growth temperature and growth phase on the lipid composition of the archaeal membrane from Thermococcus kodakaraensis. Biosci Biotechnol Biochem 731:104–108
Meister A, Blume A (2007) Self-assembly of bipolar amphiphiles. Curr Opin Colloid Interface Sci 123:138–147
Morii H, Eguchi T, Nishihara M, Kakinuma K, König H, Koga Y (1998) A novel ether core lipid with H-shaped C80-isoprenoid hydrocarbon chain from the hyperthermophilic methanogen Methanothermus fervidus. Biochim Biophys Acta 13903:339–345
Morii H, Yagi H, Akutsu H, Nomura N, Sako Y, Koga Y (1999) A novel phosphoglycolipid archaetidyl(glucosyl)inositol with two sesterterpanyl chains from the aerobic hyperthermophilic archaeon Aeropyrum pernix K1. Biochim Biophys Acta Mol Cell Biol Lipids 14363:426–436
Mozafari MR, Flanagan J, Matia-Merino L, Awati A, Omri A, Suntres ZE, Singh H (2006) Recent trends in the lipid-based nanoencapsulation of antioxidants and their role in foods. J Sci Food Agric 8613:2038–2045
Muller S, Pfannmuller M, Teuscher N, Heilmann A, Rothe U (2006) New method for surface modification of nanoporous aluminum oxide membranes using tetraether lipid. J Biomed Nanotechnol 21:16–22
Patel GB, Sprott GD (1999) Archaeobacterial ether lipid liposomes (archaeosomes) as novel vaccine and drug delivery systems. Crit Rev Biotechnol 194:317–357
Peretó J, López-García P, Moreira D (2004) Ancestral lipid biosynthesis and early membrane evolution. Trends Biochem Sci 299:469–477
Raghoebarsing AA, Pol A, van de Pas-Schoonen KT, Smolders AJP, Ettwig KF, Rijpstra WIC, Schouten S, Damste JSS, Op den Camp HJM, Jetten MSM, Strous M (2006) A microbial consortium couples anaerobic methane oxidation to denitrification. Nature 4407086:918–921
Reysenbach AL, Liu Y, Banta AB, Beveridge TJ, Kirshtein JD, Schouten S, Tivey MK, Von Damm KL, Voytek MA (2006) A ubiquitous thermoacidophilic archaeon from deep-sea hydrothermal vents. Nature 4427101:444–447
Sako Y, Nomura N, Uchida A, Ishida Y, Morii H, Koga Y, Hoaki T, Maruyama T (1996) Aeropyrum pernix gen. nov., sp. nov., a novel aerobic hyperthermophilic archaeon growing at temperatures up to 100 degrees C. Int J Syst Bacteriol 464:1070–1077
Schleper C, Puehler G, Holz I, Gambacorta A, Janekovic D, Santarius U, Klenk HP, Zillig W (1995) Picrophilus gen. nov., fam. nov.; a novel aerobic, heterotrophic, thermoacidophilic genus and family comprising archaea capable of growth around pH-0. J Bacteriol 17724:7050–7059
Schleper C, Puhler G, Klenk HP, Zillig W (1996) Picrophilus oshimae and Picrophilus torridus fam nov, gen nov, sp nov, two species of hyperacidophilic, thermophilic, heterotrophic, aerobic archaea. Int J Syst Bacteriol 463:814–816
Shimada H, Nemoto N, Shida Y, Oshima T, Yamagishi A (2008) Effects of pH and temperature on the composition of polar lipids in Thermoplasma acidophilum HO-62. J Bacteriol 19015:5404–5411
Sprott GD, Agnew BJ, Patel GB (1997a) Structural features of ether lipids in the archaeobacterial thermophiles Pyrococcus furiosus, Methanopyrus kandleri, Methanothermus fervidus, and Sulfolobus acidocaldarius. Can J Microbiol 435:467–476
Sprott GD, Tolson DL, Patel GB (1997b) Archaeosomes as novel antigen delivery systems. FEMS Microbiol Lett 1541:17–22
Sprott GD, Brisson JR, Dicaire CJ, Pelletier AK, Deschatelets LA, Krishnan L, Patel GB (1999) A structural comparison of the total polar lipids from the human archaea Methanobrevibacter smithii and Methanosphaera stadtmanae and its relevance to the adjuvant activities of their liposomes. Biochim Biophys Acta Mol Cell Biol Lipids 14402–14403:275–288
Sprott GD, Sad S, Fleming LP, Dicaire CJ, Patel GB, Krishnan L (2003) Archaeosomes varying in lipid composition differ in receptor-mediated endocytosis and differentially adjuvant immune responses to entrapped antigen. Archaea 13:151–164
Stadnitskaia A, Baas M, Ivanov MK, Tjeerd CE, Weering V, Damste JSS (2002) Novel archaeal macrocyclic diether core membrane lipids in a methane-derived carbonate crust from a mud volcano in the Sorokin Trough, NE Black Sea. Archaea 1:165–173
Takai K, Sugai A, Itoh T, Horikoshi K (2000) Palaeococcus ferrophilus gen. nov., sp nov., a barophilic, hyperthermophilic archaeon from a deep-sea hydrothermal vent chimney. Int J Syst Evol Microbiol 50:489–500
Trincone A, Nicolaus B, Palmieri G, De Rosa M, Huber R, Huber G, Stetter KO, Gambacorta A (1992) distribution of complex and core lipids within new hyperthermophilic members of the archaea domain. Syst Appl Microbiol 151:11–17
Uda I, Sugai A, Itoh Y, Itoh T (2000) Characterization of caldarchaetidylglycerol analogs, dialkyl-type and trialkyl-type, from Thermoplasma acidophilum. Lipids 3510:1155–1157
Ulrih NP, Adamlje U, Nemec M, Sentjurc M (2007) Temperature- and pH-induced structural changes in the membrane of the hyperthermophilic archaeon Aeropyrum pernix K1. J Membr Biol 2191–2193:1–8
Valentine DL (2007) Adaptations to energy stress dictate the ecology and evolution of the Archaea. Nat Rev Microbiol 54:316–323
Volkl P, Huber R, Drobner E, Rachel R, Burggraf S, Trincone A, Stetter KO (1993) Pyrobaculum aerophilum sp. nov., a novel nitrate-reducing hyperthermophilic archaeum. Appl Environ Microbiol 599:2918–2926
Wachtershauser G (2003) From pre-cells to Eukarya—a tale of two lipids. Mol Microbiol 471:13–22
William M, Michael JR (2003) On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells. Philos Trans R Soc Lond B 3581429:59–85
Woese CR, Fox GE (1977) Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proc Natl Acad Sci U S A 7411:5088–5090
Woese CR (1987) Bacterial evolution. Microbiol Mol Biol Rev 512:221–271
Woese CR, Kandler O, Wheelis ML (1990) Towards a natural system of organisms: proposal for the domains archaea, bacteria, and eucarya. Proc Natl Acad Sci U S A 87:4576–4579
Wuchter C, Abbas B, Coolen MJL, Herfort L, van Bleijswijk J, Timmers P, Strous M, Teira E, Herndl GJ, Middelburg JJ, Schouten S, Damste JSS (2006) Archaeal nitrification in the ocean. Proc Natl Acad Sci U S A 10333:12317–12322
Yamauchi K, Doi K, Yoshida Y, Kinoshita M (1993) Archaebacterial lipids: highly proton-impermeable membranes from 1, 2-diphytanyl-sn-glycero-3-phosphocoline. Biochim Biophys Acta Biomembr 11462:178–182
Zillig W, Holz I, Janekovic D, Klenk HP, Imsel E, Trent J, Wunderl S, Forjaz VH, Coutinho R, Ferreira T (1990) Hyperthermus butylicus, a hyperthermophilic sulfur-reducing archaebacterium that ferments peptides. J Bacteriol 1727:3959–3965
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ulrih, N.P., Gmajner, D. & Raspor, P. Structural and physicochemical properties of polar lipids from thermophilic archaea. Appl Microbiol Biotechnol 84, 249–260 (2009). https://doi.org/10.1007/s00253-009-2102-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-009-2102-9