
Abstract
l-Cysteine is an important amino acid in terms of its industrial applications. We previously found marked production of l-cysteine directly from glucose in recombinant Escherichia coli cells by the combination of enhancing biosynthetic activity and weakening the degradation pathway. Further improvements in l-cysteine production are expected to use the amino acid efflux system. Here, we identified a novel gene involved in l-cysteine export using a systematic and comprehensive collection of gene-disrupted E. coli K-12 mutants (the Keio collection). Among the 3,985 nonessential gene mutants, tolC-disrupted cells showed hypersensitivity to l-cysteine relative to wild-type cells. Gene expression analysis revealed that the tolC gene encoding the outer membrane channel is essential for l-cysteine tolerance in E. coli cells. However, l-cysteine tolerance is not mediated by TolC-dependent drug efflux systems such as AcrA and AcrB. It also appears that other outer membrane porins including OmpA and OmpF do not participate in TolC-dependent l-cysteine tolerance. When a low-copy-number plasmid carrying the tolC gene was introduced into E. coli cells with enhanced biosynthesis, weakened degradation, and improved export of l-cysteine, the transformants exhibited more l-cysteine tolerance and production than cells carrying the vector only. We concluded that TolC plays an important role in l-cysteine tolerance probably due to its export ability and that TolC overexpression is effective for l-cysteine production in E. coli.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
l-Cysteine is an important amino acid by virtue of its applications in the pharmaceutical, food, and cosmetic industries. Thus far, most l-cysteine has been obtained by extraction from acid hydrolysates of the keratinous proteins in human hair and animal feathers. This extraction method remains problematic for waste treatment and its yield is low. Additional applications would be feasible if the cost of l-cysteine production could be cut down, e.g., by a fermentation process. The aim of our research was to develop a process for l-cysteine fermentation directly from glucose in Escherichia coli.
The biosynthesis of l-cysteine in E. coli involves two reactions in which the hydroxyl group of l-serine is replaced by a thiol. The first step consists of the O-acetylation of l-serine by serine acetyltransferase (SAT; EC 2.3.1.30), the product of the cysE gene. The second is catalyzed by O-acetylserine sulfhydrylase, which forms l-cysteine from O-acetyl-l-serine (OAS) and sulfide (Kredich and Tomkins 1966). Because high levels of l-cysteine have been reported to be inhibitory or even toxic to cells (Datta 1967; Delaney et al. 1992; Harris and Lui 1981; Kari et al. 1971; Park and Imlay 2003; Sørensen and Pedersen 1991), intracellular l-cysteine content in E. coli is strictly controlled. Excess l-cysteine inhibits the activity of SAT, a key enzyme in the biosynthetic pathway (Kredich 1983), and therefore, l-cysteine could not be overproduced in the cell. Even if it was overproduced, it would be degraded to pyruvate, ammonia, and sulfide by at least five l-cysteine desulfhydrase (CD) enzymes (Awano et al. 2003, 2005). We have so far constructed recombinant E. coli cells that overproduce l-cysteine and l-cystine by expressing feedback inhibition-insensitive SAT (Nakamori et al. 1998; Takagi et al. 1999a, b) and disrupting the major CD genes. One of the crucial metabolic steps to achieve high-yield l-cysteine production in such a fermentation process lies in the enhancement of the export of l-cysteine out of the overproducing cells.
Four proteins have been so far identified as l-cysteine exporters in E. coli. Both YdeD and YfiK were discovered to considerably augment OAS and l-cysteine production when these proteins were overexpressed (Daßler et al. 2000; Franke et al. 2003). An ATP-binding cassette-type transporter, CydDC, exported l-cysteine from the E. coli cytoplasm to the periplasm (Pittman et al. 2002). We recently found that the multidrug transporter Bcr in the major-facilitator family type is involved in the export and overproduction of l-cysteine (Yamada et al. 2006). In contrast, two transport systems have been identified for l-cysteine uptake; one is shared with diaminopimelic acid and several l-cysteine analogues, and the other is more specific for l-cysteine (Berger and Konings 1972). However, its relationship to l-cysteine production has not yet been reported. All of these proteins are believed to be inner membrane proteins, and it has not been explained how l-cysteine is exported across inner and outer membranes. The periplasmic and outer membrane proteins involved in l-cysteine transport should also exist in E. coli cells.
Recently, the Keio collection, a complete set of viable single-gene knockout E. coli K-12 mutants, was released (Baba et al. 2006). Because high levels of l-cysteine have been reported to be toxic to cells (Harris 1981; Yamada et al. 2006), we used these mutants to screen for a novel gene required for l-cysteine tolerance (degradation or extrusion). We assumed that a lack of such genes will decrease cell viability in l-cysteine-containing medium. The screening revealed that the outer membrane channel TolC is related to l-cysteine tolerance in E. coli probably due to its export ability. This is the first report on an outer membrane l-cysteine transporter. We also showed that the TolC channel is effective in l-cysteine overproduction.
Materials and methods
Strains and plasmids
The E. coli strains and plasmids used in this study are listed in Table 1. The genomic sequenced strain MG1655 (CGSC7740; supplied by Ajinomoto), which was derived from strain K-12, was used throughout the present study. Strain W3110 was used as the host for cloning of the tolC gene. Strain BW25113 (lacIq rrnBT14 lacZWJ16 hsdR514 araBADAH33 rhaBADLD78) and two independent sets of a single-gene knockout mutant collection of all the 3,985 nonessential genes of BW25113 (the Keio collection; Baba et al. 2006), which was created by replacing the open reading frame coding regions with a kanamycin resistance cassette (Datsenko and Wanner 2000), were used for the gene screening. Strain JW 3686 was a BW25113-based tnaA::Kmr (kanamycin resistant) transductant. Strain MG1655ΔtnaA was constructed by P1 transduction from JW3686 to MG1655. Strains MG1655 harboring pCEM256I and pYdeD and MG1655ΔtnaA harboring pDES were designated as l-cysteine producer and overproducer, respectively.
The middle-copy-number pCA24N-based plasmids (pTolC, pOmpA, pOmpC, pOmpF, pOmpT, and pOmpX; Kitagawa et al. 2005) and pUC119-based plasmids (pUCacrAB, pUCacrEF, pUCemrAB, and pUCmacAB; supplied by A. Yamaguchi; Nishino and Yamaguchi 2001; Yamada et al. 2006), each of which carries the corresponding wild-type gene, were used for the genetic complementation study. The low-copy-number vector pLS219 (supplied by N. Tsukagoshi) was derived from pMW219 by replacing the bacterial kanamycin resistance gene with a bacterial spectinomycin resistance gene. Plasmid pLSTolC (supplied by N. Tsukagoshi) is pLS219-derived recombinant plasmid, which contains a 2.5-kb EcoRI-HindIII fragment carrying the E. coli W3110 tolC gene. Plasmid pCEM256I is a derivative of pBluescript II SK(+) (Toyobo, Osaka, Japan) carrying the bacterial ampicillin resistance gene and the altered cysE gene encoding the Met256Ile mutant SAT, which was constructed as reported previously (Nakamori et al. 1998). The middle-copy-number vector pSTV29 and pACYC184 were purchased from Takara Bio (Ohtsu, Japan) and Nippon Gene (Tokyo, Japan), respectively.
To obtain the ydeD gene, a polymerase chain reaction (PCR) was performed with a set of primers 5′-CGC GGA TCC AAT GGT CAT AAA TGG CAG CGT AGC GC-3′ and 5′-CGC GGA TCC GCA GGG CGT TGC GGA ACA AAC-3′ with chromosomal DNA of MG1655 as a template. PCR product was digested with BamHI and subsequently cloned into pSTV29 to give pYdeD. Plasmid pDES (supplied by Ajinomoto) is a derivative of pACYC184 containing the altered cysE gene encoding the l-cysteine feedback inhibition-insensitive mutant SAT (Thr167Ala), the wild-type ydeD gene encoding inner membrane l-cysteine transporter (Daßler et al. 2000), and the altered serA gene encoding the l-serine feedback inhibition-insensitive mutant D-3-phosphoglycerate dehydrogenase (Thr410stop). Each gene fragment is under control of the constitutive promoter of the E. coli ompA gene encoding outer membrane protein A precursor. The altered serA gene is oriented in the same direction, the altered cysE and ydeD genes are in the opposite direction of replication origin, as described previously (Winterhalter and Leinfelder 1997).
Media and cultivation
Unless otherwise stated, Lennox (L) broth [1% Bacto Tryptone (Difco Laboratories, Detroit, Mich.), 0.5% Bacto Yeast Extract (Difco), 0.5% NaCl], Luria–Bertani (LB) medium (1% Bacto Tryptone, 0.5% Bacto Yeast Extract, 1% NaCl), and SM1 minimal medium [100 mM potassium phosphate buffer (pH 7.0), 75.7 mM (NH4)2SO4, 1.7 mM NaCl, 1.0 mM MgSO4, 0.1 mM CaCl2, 7.2 µM FeSO4, 3.4 mM trisodium citrate, 0.6 µM Na2MoO4, 40.4 µM H3BO3, 2.9 µM CoCl2, 1 µM CuSO4, 8.1 µM MnCl2, 1 µM ZnSO4, 3.0% glucose (wt/vol) as the carbon source] were used for the general cultivations. If necessary, ampicillin (50 µg/ml), chloramphenicol (40 µg/ml), kanamycin (50 µg/ml), tetracycline (10 µg/ml), or/and spectinomycin (100 µg/ml) was added. For solid media, 1.5% (wt/vol) agar was added. Where indicated, SM1 was supplemented with L broth, l-methionine, and thiosulfate. The pH was adjusted to 7.0 using NH3. Cultures were incubated aerobically by vigorous shaking at 34°C or 37°C. Growth was monitored by measurement of the optical density at 660 nm (OD660). Glucose concentration was measured by Glucose C2 kit (Wako, Osaka, Japan).
Screening of the genes involved in l-cysteine tolerance
The media used for primary screening was LB medium containing kanamycin (25 µg/ml), 1.0% agar, and l-cysteine. The mutants from Keio collection restored in 384-well microplates were carefully inoculated with a 384-pin plastic replicator onto the surface of the screening plates. The plates were incubated at 37°C for 18 to 20 h. l-Cysteine sensitivity was assessed by examining the growth phenotype on the screening agar medium containing 0, 15, 20, and 25 mM of l-cysteine. The mutants being incapable of growing even in the absence of l-cysteine, and/or the clones which showed different growth between two sets, were excluded from the results.
Preparation of membrane proteins
E. coli cells grown in L medium were harvested from the culture (OD660 of 0.6) by centrifugation (10 min at 5,000×g), suspended in cold 50 mM phosphate buffer (pH 7.2), and broken by sonication. Unbroken cells were removed by the centrifugation. The supernatant was centrifuged at 100,000×g for 45 min at 4°C. The precipitate was washed and incubated in the same phosphate buffer containing 0.5% sodium dodecyl sarcosinate (sarcosyl) for 30 min at room temperature at a protein concentration of 3 mg/ml. The suspension was centrifuged at 100,000×g for 45 min at 10°C. The supernatant was recovered and used as the sarcosyl-soluble fraction (the inner membrane protein fraction). The precipitate was suspended with 0.5% sodium dodecyl sulfate (SDS) and used as the sarcosyl-insoluble fraction (the outer membrane protein fraction).
SDS-polyacrylamide gel electrophoresis
Samples were dissolved in a solubilization buffer containing 1% (wt/vol) SDS, 2.5% (vol/vol) β-mercaptoethanol, 20% (vol/vol) glycerol, and 16 mM Tris–HCl (pH 6.8) and incubated in a boiling water bath for 5 min. The samples were loaded on SDS-polyacrylamide gels.
Determination of l-cysteine
The amount of l-cysteine in culture supernatants was determined according to the method of Gaitonde (1967). Before adding a Gaitonde reagent (250 mg ninhydrin dissolved in a mixture of 4 ml of HCl and 16 ml of acetic acid), l-cystine in the samples was reduced by incubation with 5 mM dithiothreitol in 100 mM Tris–HCl buffer (pH 8.6) for 10 min. The reaction products were diluted with 99.5% (vol/vol) ethanol prior to measurement of the absorbance at 560 nm.
Results
Screening of the genes involved in l-cysteine tolerance
Mutants of the Keio collection were first screened on solid LB medium containing various l-cysteine concentrations (0–25 mM), since it is well known that E. coli cells undergo growth inhibition by l-cysteine. Among the 3,985 deletion mutants, we obtained 24 strains (0.6%) that are more sensitive to l-cysteine than the wild-type strain, but no mutants displayed l-cysteine tolerance. The genes deleted in these 24 mutants that were clearly defective in growth by l-cysteine were considered to be those required for tolerance to l-cysteine. Table 2 shows the function of each gene product. It is noteworthy that several mutants were missing the gene for redox homeostasis (dsbA, cydB, nuoL, nuoB, nuoC, and nfnB), l-cysteine desulfurase (iscS), or transporter (tolC and tolB). We further studied the tolC gene encoding an outer membrane channel.
The tolC gene was identified as a gene involved in l-cysteine tolerance
To confirm whether or not the tolC gene is involved in l-cysteine resistance in E. coli, low-copy-plasmid pLSTolC carrying tolC or the empty vector pLS219 was introduced into the ΔtolC strain (JW5503). The transformed cells grown in L plus kanamycin and spectinomycin medium were spotted onto an L agar plate containing l-cysteine. As shown in Fig. 1a, overexpression of the tolC gene complemented the l-cysteine sensitivity of the ΔtolC mutant. This result shows that the tolC gene was indeed involved in l-cysteine resistance. TolC overexpression with middle-copy-plasmid pTolC also gave a similar result, but cell growth did not completely revert to the level of the wild-type cells (BW25113 carrying pCA24N) (Fig. 1b). It is contemplated that overexpression of the outer membrane protein might influence on the expression of other genes, thereby inhibiting growth (Click et al. 1988). We concluded that TolC overexpression with low-copy-number plasmid pLSTolC is preferable for further study. The actual expression of the TolC protein was confirmed by SDS-polyacrylamide gel electrophoresis of the sarcosyl-insoluble fraction (outer membrane fraction; Fig. 1c).

Effect of the tolC gene on l-cysteine tolerance of E. coli. Growth phenotypes on l-cysteine containing medium of wild-type (BW25113) and ΔtolC mutant cells harboring low-copy-number vector pLS219 or pLSTolC (a) and middle-copy-number vector pCA24N or pTolC (b). Overnight culture of each cell and dilutions of 10−2, 10−4, and 10–6 (from left to right) were spotted and incubated onto L agar plates containing 10 mM l-cysteine at 37°C for 12 h. (c) SDS-polyacrylamide gel electrophoresis of BW25113 and ΔtolC mutant cells harboring pLS219 or pLSTolC. After cultivation of each cell in L medium at 37°C for 12 h, the outer membrane protein fraction was isolated and separated in a 12% SDS-polyacrylamide gel. Protein was stained with Coomassie brilliant blue R-250
Because TolC is a well-known component of multidrug efflux pumps in E. coli, we assumed that l-cysteine sensitivity of the ΔtolC mutant results mainly from the deficiency in l-cysteine transport. Hence, we examined whether or not the TolC protein constitutes an efflux pump such as an AcrAB complex (Fig. 2a).

Effects of the TolC-dependent drug efflux systems on l-cysteine tolerance of E. coli. (a) Schematic representation of tripartite efflux pumps in Gram-negative bacteria, using as an example the AcrAB–TolC efflux pump from E. coli. The outer membrane channel TolC interacts with an inner membrane transporter, AcrB, which provides export energy, and a protein of the adaptor family, AcrA, which brings TolC and the inner membrane transporter together. Substrates are extruded from both the cytoplasm and periplasm (Elkins and Beenken 2005). (b) Growth phenotypes on l-cysteine containing medium of E. coli strains. Overnight culture of each BW25113, ΔtolC, ΔacrA, ΔacrB, ΔacrE, ΔacrF, ΔemrA, ΔemrB, ΔmacA, and ΔmacB mutant cell and serial dilutions of 10–1 to 10–5 (from left to right) were spotted and incubated onto L agar plates containing 10 mM l-cysteine at 37°C for 12 h
l-Cysteine tolerance of E. coli is not mediated by TolC-dependent drug efflux systems
It is known that TolC forms a channel and serves as an essential element of some mechanisms responsible for tolerance to various compounds, including hydrophobic antibiotics and organic solvents (Koronakis et al. 2004). To examine whether or not l-cysteine is exported to medium by constituting a known efflux pump other than the TolC protein, cells of each deletion mutant (acrA, acrE, emrA, and macA encoding the periplasmic membrane fusion protein and acrB, acrF, emrB, and macB encoding the inner membrane pump) were spotted onto agar medium containing l-cysteine (Fig. 2b). None of the gene deletions influenced the growth phenotype of wild-type cells, indicating that l-cysteine was exported to the medium by a single TolC or unknown efflux system.
Other outer membrane porins are not involved in the TolC-dependent l-cysteine tolerance
The TolC protein is made of three monomers (Koronakis et al. 2004), which is typical of many outer membrane porins that form narrow channels to allow the passage of nutrients into the cell. We first tested the l-cysteine sensitivities of various mutants that are defective in the outer membrane porin (OmpA, OmpC, OmpF, OmpT, and OmpX). As shown in Fig. 3a, none of those mutants exhibited growth inhibition. The ompC and ompF mutants appeared to grow slowly on the plate containing l-cysteine relative to the other mutants, but the difference between the growths of all mutants was not significant after incubation for >36 h (data not shown). On the other hand, to further examine whether or not those genes could complement the l-cysteine sensitivity of the ΔtolC mutant, the plasmid carrying each of these genes was introduced into ΔtolC mutant cells (Fig. 3b). Interestingly, overexpression of these genes did not confer l-cysteine tolerance to tolC-disrupted cells. These results suggest that the function of TolC on l-cysteine is absolutely different from those of other porin proteins.

Effects of the outer membrane porins on l-cysteine tolerance of E. coli. Growth phenotypes on l-cysteine containing medium of strains BW25113, ΔtolC, ΔompA, ΔompC ΔompF, ΔompT, or ΔompX (a) and ΔtolC mutant harboring pCA24N, pTolC, pOmpA, pOmpC, pOmpF, pOmpT, or pOmpX (b). Overnight culture of each cell and serial dilutions of 10–1 to 10–5 (a), and dilutions of 10–2, 10–4, and 10–6 (b; from left to right), were spotted and incubated onto L agar plates containing 10 mM l-cysteine at 37°C for 12 h
Overexpression of TolC confers l-cysteine tolerance in E. coli cells
YdeD was previously reported to be an inner membrane l-cysteine transporter, which excretes both the precursor (OAS) of the inducer molecule (N-acetylserine; NAS) and the major end product (l-cysteine) from the E. coli cytoplasm to the periplasmic space (Daßler et al. 2000). Overexpression of YdeD conferred E. coli cells with drastic tolerance to l-cysteine. We therefore transformed MG1655 cells with pLSTolC carrying the tolC gene and/or pYdeD carrying the ydeD gene and subsequently examined the effect of their overexpression on the growth of E. coli cells in the presence of l-cysteine (Fig. 4a). In agreement with the previous results (Daßler et al. 2000), E. coli cells overexpressing YdeD exhibited greater tolerance to l-cysteine than wild-type cells. We also found that co-overexpression of TolC improves the growth of cells overexpressing only YdeD in the presence of l-cysteine. At the stationary growth phase, it appears that cells overexpressing single TolC showed slightly higher l-cysteine tolerance than those carrying the vector only. Furthermore, BW25113ΔtolC cells overexpressing both YdeD and TolC showed significantly higher l-cysteine tolerance than those overexpressing YdeD alone (Fig. 4b). Improvement of the growth by the co-expression of TolC and YedD was significant in tolC-disrupted BW25113 cells relative to MG1655 cells. This is probably because TolC is constitutively expressed in MG1655 (the wild-type strain) at the basal level (Aono et al. 1998). These results strongly suggest that the TolC protein plays an important role in l-cysteine export from the E. coli periplasm to the medium.

Effects of the TolC and YdeD proteins on l-cysteine tolerance of E. coli. Growth (OD660) of strain MG1655 with pSTV29 and pLS219 (open triangles), with pSTV29 and pLSTolC (closed triangles), with pYdeD and pLS219 (open circles), or pYdeD and pLSTolC (closed circles) (a) and strain BW25113ΔtolC with pSTV29 and pLS219 (open triangles), with pSTV29 and pLSTolC (closed triangles), with pYdeD and pLS219 (open circles), or pYdeD and pLSTolC (closed circles) (b). Cells were grown in SM1 medium plus 10 mM l-cysteine at 37°C. Values indicate means of results and SDs of results from three independent experiments
TolC overexpression contributes to l-cysteine productivity in E. coli cells
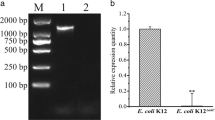
We previously found marked production of l-cysteine plus l-cystine from glucose in E. coli cells expressing the altered cysE gene encoding the mutant SAT (Nakamori et al. 1998; Takagi et al. 1999a, b). Also, overexpression of YdeD caused increases in l-cysteine levels in medium (Daßler et al. 2000). It was therefore expected that l-cysteine productivity would be further improved by introducing both pLSTolC and pYdeD into cells that express the mutant SAT. Plasmid pCEM256I, which carries an altered cysE gene encoding the Met256Ile mutant SAT, which is less sensitive to feedback inhibition, was first introduced into MG1655 cells to enhance l-cysteine biosynthesis. Next, the resultant cells were transformed by pYdeD and/or pLSTolC, and were subsequently cultivated at 37°C for l-cysteine production. Figure 5 shows the time course of growth and glucose consumption (Fig. 5a) and l-cysteine plus l-cystine production (Fig. 5b). The difference of the available l-cysteine level in each cell may have been a cause for the difference of growth between MG1655 harboring pYdeD and pLSTolC and MG1655 with pYdeD (Fig. 5a). The MG1655 cells harboring both pCEM256I and pYdeD accumulated 70 mg/l of l-cysteine plus l-cystine after 24 h of cultivation (designated as l-cysteine producer). It is noteworthy that when the tolC gene was overexpressed, the extracellular l-cysteine level (119 mg/l) was approximately 70% higher than that of the l-cysteine producer. However, the decrease in l-cysteine production commenced after 24 h of cultivation, probably due to the CD enzymes.

Time courses of growth [OD660 (solid lines) and glucose consumption (dotted lines)] (a) and l-cysteine plus l-cystine production (b) by cells of strain MG1655 with pBluescript II SK(+), pSTV29, and pLS219 (open triangles), pCEM256I, pYdeD, and pLS219 (open circles), or pCEM256I, pYdeD, and pLSTolC (closed circles). The medium without cell inoculation was used as a control (open diamonds). Cells were grown in SM1 medium containing 10% L broth, 3% glucose, and 50 mM MgSO4 at 37°C
To improve and maintain l-cysteine productivity, plasmid pDES carrying the altered cysE, ydeD, and altered serA gene was introduced into MG1655ΔtnaA cells (Awano et al. 2003). The transformed cells were designated as l-cysteine overproducers. We also investigated TolC’s effects on growth and l-cysteine productivity in the l-cysteine overproducer (Fig. 6). As shown in Fig. 6b, after 40 h of cultivation, MG1655ΔtnaA cells carrying pDES and pLSTolC produced a slightly yet significantly higher amount of l-cysteine plus l-cystine (1,127 ± 75 mg/l) than MG1655ΔtnaA cells carrying only pDES (958 ± 71 mg/l). These results indicate that TolC overexpression is effective for l-cysteine overproduction in E. coli cells.

Time courses of growth (OD660; a) and l-cysteine plus l-cystine production (b) by cells of strain MG1655ΔtnaA with pACYC184 and pLS219 (open triangles), pACYC184 and pLSTolC (closed triangles), pDES and pLS219 (open circles), or pDES and pLSTolC (closed circles). Values indicate means and SDs of results from three independent experiments. Cells were grown at 34°C in SM1 medium containing 10% L broth, 3% glucose, and 450 mg/l l-methionine. After cultivation for 6 and 12 h, 10 and 20 mM sodium thiosulfate was added, respectively
Discussion
Four of known l-cysteine transporters (YdeD, CydDC, YfiK, and Bcr) are involved in the efflux of l-cysteine from the E. coli cytoplasm. All of them are inner membrane proteins that transport l-cysteine from the cytoplasm into the periplasmic space. This is the first report to indicate that the outer membrane channel probably releases l-cysteine into the medium. Our results strongly suggest that passive or facilitated diffusion through the TolC porin increases the outer membrane permeability to l-cysteine. We also conclude that TolC is important for l-cysteine tolerance and overproduction.
It was previously reported that overexpression of the YdeD or Bcr protein in E. coli is important for effective l-cysteine production (Daßler et al. 2000; Yamada et al. 2006). Elucidation of the molecular mechanisms involved in l-cysteine export from the periplasm to the medium remains one of the most intriguing problems of l-cysteine excretion. High concentrations of l-cysteine have also been reported to be inhibitory or even toxic to E. coli cells. The complete collection of viable single-gene knockout E. coli K-12 mutants (Baba et al. 2006) was used to screen for a novel l-cysteine transporter in this study. We found that the TolC channel is apparently involved in l-cysteine tolerance (Fig. 1 and Table 2). We assumed that the mutants that cannot survive in the presence of excess l-cysteine must lack the gene(s) essential for excluding l-cysteine (degradation or extrusion) from the cells. Among 3,985 gene deletion mutants, only tolC-disrupted cells were found to be sensitive to l-cysteine (5 mM). However, five CD enzymes involved in l-cysteine degradation (tryptophanase, cystathionine β-lyase, MalY protein, O-acetyl l-serine sulfhydrylase-A, and O-acetyl l-serine sulfhydrylase-B; Awano et al. 2003, 2005) did not show sensitivity to l-cysteine under these experimental conditions. This is probably because single CD gene deletion mutants still had CD activity (Awano et al. 2003). Interestingly, a null mutant of the iscS gene (Fuentes et al. 2007), which encodes l-cysteine desulfurase that converts l-cysteine to l-alanine, was sensitive to l-cysteine (Table 2). These results also suggested that this enzyme is important for controlling intracellular l-cysteine level by a different reaction with the known CD enzymes (catalyzing conversion of l-cysteine to pyruvate, ammonia, and sulfide).
The TolC protein plays a common role in the expulsion of diverse molecules, which include protein toxins and antibacterial drugs, from the cell (Koronakis et al. 2004). We presumed that TolC-dependent l-cysteine tolerance is due to l-cysteine extrusion. In several efflux pumps, TolC acts with an inner membrane translocase and an accessory or adaptor protein, e.g., AcrAB and AcrEF from the resistance nodulation division (RND)-type drug transporter family, EmrAB from the major facilitator (MF) family, and MacAB from the ATP-binding cassette (ABC) family. For example, the AcrAB–TolC efflux pump is essential for E. coli cells to maintain organic solvent tolerance (Aono et al. 1998; Fralick 1996; White et al. 1997). AcrEF–TolC plays an important role in the normal maintenance of cell division, which might be related to its transport function (Lau and Zgurskaya 2005). EmrAB–TolC was identified as a transporter involved in multidrug resistance (Lomovskaya and Lewis 1992). MacAB–TolC confers resistance against macrolide antibiotics (Kobayashi et al. 2001). If these tripartite multidrug efflux systems are involved in l-cysteine extrusion, deficiency in any one of the genes will result in l-cysteine sensitivity. Interestingly, each mutant showed the same level of growth inhibition as wild-type cells, indicating that these proteins do not contribute to l-cysteine tolerance (Fig. 2b). It is possible that TolC might form another novel tripartite efflux pump for l-cysteine export. Because cysteine is likely to diffuse easily through general porin channels, the effect of TolC is probably due to a small contribution of a broad substrate-range efflux process (Lee et al. 2000) in the collaboration between the cytosol-to-periplasm transporters and the tripartite transporters.
According to an X-ray crystallographic examination at 2.1 Å resolutions (Koronakis et al. 2000), TolC is open to the outside medium but is closed at its periplasmic entrance with an effective diameter of ∼3.9 Å. This pore size might be sufficient to allow l-cysteine diffusion across the E. coli outer membrane. As shown in Fig. 3b, other outer membrane porins did not complement the l-cysteine sensitivity of ΔtolC mutant cells. Deletion mutants of these genes, including the ompF gene, which encodes the 11-Å-wide outer membrane porin OmpF (Koronakis et al. 2004), did not display the l-cysteine sensitivity (Fig. 3a). The porin TolC has a different function or functions than other outer membrane porins for l-cysteine diffusion and is important for the maintenance of the basal level of l-cysteine tolerance in E. coli by exporting excess l-cysteine accumulated in periplasm to outside the medium (Figs. 3 and 4). Intriguingly, these results suggest that growth inhibition by l-cysteine may occur in the periplasm, where disulfide bonds are formed, and predominantly in oxidizing environment. The presence of reductants such as l-cysteine or glutathione may perturb the redox homeostasis, leading to growth inhibition (Pittman et al. 2002; 2005).
It was previously reported that E. coli cells overexpressing the inner membrane protein YdeD, which belongs to the PecM family of transporters, excreted considerable amounts of l-cysteine, OAS, and maybe NAS (Daßler et al. 2000). Interestingly, the tolC deletion mutant was sensitive to these compounds but not to l-serine (data not shown), suggesting that YdeD transports these compounds from the cytoplasm into the periplasm and that TolC subsequently releases them into the outside medium. We predicted that overexpression of both proteins could drastically increase l-cysteine productivity. In fact, overexpression of TolC indeed slightly increased l-cysteine production (Figs. 5 and 6). Such an approach could be promising for the effective fermentative production of l-cysteine. It should be noted that overexpression of TolC did not show an enormous effect on the growth phenotype. This is probably because the TolC protein is highly expressed in E. coli cells at the basal level. On the other hand, tolC-disrupted cells that overexpress TolC could grow well in the presence of l-cysteine (Figs. 1a and 4b), indicating that TolC is important for l-cysteine tolerance.
It is known that TolC on the outer membrane forms a channel for various compounds not only organic solvents or hydrophobic antibiotics, but also hydrophilic compounds such as aminoglycoside (Rosenberg et al. 2000) or l-cysteine as reported in this study. Krishnamoorthy et al. reported that even replacement of ArcB in the E. coli AcrAB–TolC complex by MexB in the Pseudomonas aeruginosa AcrA–MexB–TolC complex was still partially functional (Krishnamoorthy et al. 2008). This implies that the diversity of substrates could result from the flexible interaction of constituting components. Although it is unknown whether YdeD interacts directly with TolC, it is possible that the artificial fusion of YdeD with TolC or the periplasmic adaptor protein, e.g., AcrA, may form a novel efflux for l-cysteine extrusion directly from the cytoplasm, resulting in higher l-cysteine productivity. The effect of TolC overexpression on l-cysteine tolerance was smaller than expected before, compared with the hypersensitivity of the null mutant. This ambiguous observation implied that TolC should have another essential function besides that of a channel. As shown in Fig. 7, we found that the tolC null mutant was more sensitive to dithiothreitol (DTT) than the DTT-hypersensitive dsbA null mutant. It seems likely that the bifunctional TolC protects E. coli cells from reductants or is involved in redox homeostasis in the cells

Effects of the outer membrane channels on DTT sensitivity of E. coli. Growth phenotypes on DTT containing medium of strains BW25113, ΔdsbA, and ΔtolC harboring pCA24N, pTolC, or pDsbA. After cultivation for overnight, dilutions of 10–2, 10–4 10–5, and 10–6 of each cell (from left to right) were spotted and incubated onto LB agar plates containing 5 or 10 mM DTT at 37°C for 12 h
References
Aono R, Tsukagoshi N, Yamamoto M (1998) Involvement of outer membrane protein TolC, a possible member of the mar-sox regulon, in maintenance and improvement of organic solvent tolerance of Escherichia coli K-12. J Bacteriol 180:938–994
Awano N, Wada M, Kohdoh A, Oikawa T, Takagi H, Nakamori S (2003) Effect of cysteine desulfhydrase gene disruption on l-cysteine overproduction in Escherichia coli. Appl Microbiol Biotechnol 62:239–243
Awano N, Wada M, Mori H, Nakamori S, Takagi H (2005) Identification and functional analysis of cysteine desulfhydrases in Escherichia coli. Appl Environ Microbiol 71:4149–4152
Baba T, Ara T, Hasegawa M, Takai Y, Okumura Y, Baba M, Datsenko KA, Tomita M, Wanner BL, Mori H (2006) The construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Methods Mol Biol 416:171–181
Berger EA, Konings LA (1972) A binding protein involved in the transport of cystine and diaminopimelic acid in Escherichia coli. J Biol Chem 247:7684–7694
Click EM, McDonald GA, Schnaitman CA (1988) Translational control of exported proteins that results from OmpC porin overexpression. J Bacteriol 170:2500–2011
Daßler T, Maier T, Winterhalter C, Böck A (2000) Identification of a major facilitator protein from Escherichia coli involved in efflux of metabolites of the cysteine pathway. Mol Microbiol 36:1101–1112
Datsenko KA, Wanner BL (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci USA 97:6640–6645
Datta P (1967) Regulation of homoserine biosynthesis by l-cysteine, a terminal metabolite of a linked pathway. Proc Natl Acad Sci 58:635–641
Delaney JM, Ang D, Georgopoulos C (1992) Isolation and characterization of the Escherichia coli htrD gene, whose product is required for growth at high temperatures. J Bacteriol 174:1240–1247
Elkins CA, Beenken KE (2005) Modeling the tripartite drug efflux pump achetype: structural and functional studies of the macromolecular constituents reveal more than their names imply. J Chemother 17:581–592
Fralick JA (1996) Evidence that TolC is required for functioning of the Mar/AcrAB efflux pump of Escherichia coli. J Bacteriol 178:5803–5805
Franke I, Resch A, Daßler T, Maier T, Böck A (2003) YfiK from Escherichia coli promotes export of O-acetylserine and cysteine. J Bacteriol 185:1161–1166
Fuentes DE, Fuentes EL, Castro ME, Pérez JM, Araya MA, Chasteen TG, Pichuantes SE, Vásquez CC (2007) Cysteine metabolism-related genes and bacterial resistance to potassium tellurite. J Bacteriol 189:8953–8960
Gaitonde MK (1967) A spectrophotometric method for the direct determination of cysteine in the presence of other naturally occurring amino acids. Biochem J 104:627–633
Harris CL (1981) Cysteine and growth inhibition of Escherichia coli: threonine deaminase as the target enzyme. J Bacteriol 145:1031–1035
Harris CL, Lui L (1981) Cysteine and growth inhibition of Escherichia coli: depression of the ilvGEDA operon. Biochem Biophys Res Commun 101:1145–1151
Kari C, Nagy Z, Kovacs P, Hernadi F (1971) Mechanism of the growth inhibitory effect of cysteine on Escherichia coli. J Gen Microbiol 68:349–356
Kitagawa M, Ara T, Arifuzzaman M, Ioka-Nakamichi T, Inamoto E, Toyonaga H, Mori H (2005) Complete set of ORF clones of Escherichia coli ASKA library (A complete set of E. coli K-12 ORF archive): unique resources for biological research. DNA Res 12:291–299
Kobayashi N, Nishino K, Yamaguchi A (2001) Novel macrolide-specific ABC-type efflux transporter in Escherichia coli. J Bacteriol 183:5639–5644
Koronakis V, Sharff A, Koronakis E, Luisi B, Hughes C (2000) Crystal structure of the bacterial membrane protein TolC central to multidrug efflux and protein export. Nature 405:914–919
Koronakis V, Eswaran J, Hughes C (2004) Structure and function of TolC: the bacterial exit duct for proteins and drugs. Annu Rev Biochem 73:467–489
Kredich NM (1983) Regulation of cysteine biosynthesis in Escherichia coli and Salmonella typhimurium. In: Herrmann KM, Sommerville (eds) Amino acids: biosynthesis and genetic regulation. Addison-Wesley, United Kingdom, pp 115–132
Kredich NM, Tomkins GM (1966) The enzymic synthesis of l-cysteine in Escherichia coli and Salmonella typhimurium. J Biol Chem 241:4955–4965
Krishnamoorthy G, Tikhonova EB, Zgurskaya HI (2008) Fitting periplasmic membrane fusion proteins to inner membrane transporters: mutations that enable Escherichia coli AcrA to function with Pseudomonas aeruginosa MexB. J Bacteriol 190:691–698
Lau SY, Zgurskaya HI (2005) Cell division defects in Escherichia coli deficient in the multidrug efflux transporter AcrEF-TolC. J Bacteriol 187:7815–7825
Lee A, Mao W, Warren MS, Mistry A, Hoshino K, Okumura R, Ishida H, Lomovskaya O (2000) Interplay between efflux pumps may provide either additive or multiplicative effects on drug resistance. J Bacteriol 182:3142–3150
Lomovskaya O, Lewis K (1992) Emr, an Escherichia coli locus for multidrug resistance. Proc Natl Acad Sci USA 89:8938–8942
Nakamori S, Kobayashi S, Kobayashi C, Takagi H (1998) Overproduction of l-cysteine and l-cystine by Escherichia coli strains with a genetically altered serine acetyltransferase. Appl Environ Microbiol 64:1607–1611
Nishino K, Yamaguchi A (2001) Analysis of a complete library of putative drug transporter genes in Escherichia coli. J Bacteriol 183:5803–5812
Park S, Imlay JA (2003) High levels of intracellular cysteine promote oxidative DNA damage by driving the Fenton reaction. J Bacteriol 185:1942–1950
Pittman MS, Corker H, Wu G, Binet MB, Moir AJG, Poole RK (2002) Cysteine is exported from the Escherichia coli cytoplasm by CydDC, an ATP-binding cassette-type transporter required for cytochrome assembly. J Biol Chem 277:49841–49849
Pittman MS, Robinson HC, Poole RK (2005) A Bacterial glutathione transporter (Escherichia coli CydDC) exports reductant to the periplasm. J Biol Chem 280:32254–32261
Rosenberg EY, Ma D, Nikaido H (2000) AcrD of Escherichia coli is an aminoglycoside efflux pump. J Bacteriol 182:1754–1756
Sørensen MA, Pedersen S (1991) Cysteine, even in low concentrations, induces transient amino acid starvation in Escherichia coli. J Bacteriol 173:5244–5246
Takagi H, Awano N, Kobayashi S, Kobayashi C, Noji M, Saito K, Nakamori S (1999a) Overproduction of l-cysteine and l-cystine by expression of genes for feedback inhibition-insensitive serine acetyltransferase from Arabidopsis thaliana in Escherichia coli. FEMS Microbiol Lett 179:453–459
Takagi H, Kobayashi C, Kobayashi S, Nakamori S (1999b) PCR random mutagenesis into Escherichia coli serine acetyltransferase: isolation of the mutant enzymes that cause overproduction of l-cysteine and l-cystine due to the desensitization to feedback inhibition. FEBS Lett 452:323–327
White DG, Goldman JD, Demple B, Levy SB (1997) Role of the acrAB locus in organic solvent tolerance mediated by expression of marA, soxS, or robA in Escherichia coli. J Bacteriol 179:6122–6126
Winterhalter C, Leinfelder W (1997) Microorganisms and process for the fermentative production of l-cysteine, l-cystine, N-acetyl-serine or thiazolidin-derivates. Consortium für elektrochemische Industrie GmbH, assignee. European patent 0885962A1
Yamada S, Awano N, Inubushi K, Maeda E, Nakamori S, Nishino K, Yamaguchi A, Takagi H (2006) Effect of drug transporter genes on cysteine export and overproduction in Escherichia coli. Appl Environ Microbiol 72:4735–4742
Acknowledgments
We greatly appreciate R. Takeuchi, N. Yoshida, and S. Morigasaki (Nara Institute of Science and Technology, Japan) for their helpful assistance and discussion on this work, respectively. We thank N. Tsukagoshi (Tokyo Institute of Technology, Yokohama, Japan), A. Yamaguchi (Osaka University, Osaka, Japan), and Ajinomoto, Co., Inc. (Tokyo, Japan) for providing the strains and plasmids. This work was supported in part by KAKENHI (Grant-in-Aid for Scientific Research) on Priority Areas “Applied Genomics” from the Ministry of Education, Culture, Sports, Science and Technology of Japan and by a grant from Ajinomoto Co., Inc. to H.T.
Author information
Authors and Affiliations
Corresponding author
Additional information
Natthawut Wiriyathanawudhiwong and Iwao Ohtsu contributed equally to this work.
Rights and permissions
About this article
Cite this article
Wiriyathanawudhiwong, N., Ohtsu, I., Li, ZD. et al. The outer membrane TolC is involved in cysteine tolerance and overproduction in Escherichia coli . Appl Microbiol Biotechnol 81, 903–913 (2009). https://doi.org/10.1007/s00253-008-1686-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-008-1686-9