
Abstract
The α-glucosidase inhibitor acarbose, O-{4,6-dideoxy-4[1s-(1,4,6/5)-4,5,6-trihydroxy-3-hydroxymethyl-2-cyclohexen-1-yl]-amino-α-d-glucopyranosyl}-(1→4)-O-α-d-glucopyranosyl-(1→4)-d-glucopyranose, is produced in large-scale fermentation by the use of strains derived from Actinoplanes sp. SE50. It has been used since 1990 in many countries in the therapy of diabetes type II, in order to enable patients to better control blood sugar contents while living with starch-containing diets. Thus, it is one of the latest successful products of bacterial secondary metabolism to be introduced into the pharmaceutical world market. Cultures of Actinoplanes sp. also produce various other acarbose-like components, of which component C is hard to separate during downstream processing, which is one of the most modern work-up processes developed to date. The physiology, genetics and enzymology of acarbose biosynthesis and metabolism in the producer have been studied to some extent, leading to the proposal of a new pathway and metabolic cycle, the “carbophore”. These data could give clues for further biotechnological developments, such as the suppression of side-products, enzymological or biocombinatorial production of new metabolites and the engineering of production rates via genetic regulation in future.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Historically, the first wave of designed screening of bacterial and other microbial secondary metabolites was aimed at the detection of anti-infective substances (antibiotics), such as antimicrobials and antivirals, and was started during world war two. Later, starting in the 1960s, other activities of potential use in many applications were also searched for in similar screening protocols. These included in the first instance microbial inhibitors of hydrolases, such as proteases and glycosidases, but also comprised antitumor agents, immunomodulators, cardiovascular agents, inhibitors or modulators of human intracellular enzymes, signal transduction components and agents involved in the cell cycle and functions of the cell skeleton (see e.g. Demain et al. 1989; Rehm et al. 1997). Glycosidase inhibitors were suggested for pharmaceutical or agricultural applications, e.g. obesity, adipose, hyperlipidemia (arteriosclerosis), diabetes, pre-diabetes, gastritis, gastric ulcer, duodenal ulcer and caries in man, or as food additive for various purposes in farm animals (Frommer et al. 1975, 1977a, 1977b, 1979). First, α-glucosidase inhibitors from plants were tested in animal trials for use in humans, but soon turned out not to be efficient (Puls and Keup 1973, Puls et al. 1977; Bischoff et al. 1994). In screenings among actinomycetes, besides the pseudooligosaccharides described herein, polypeptide-based amylase inhibitors were also detected and developed, e.g. tendamistat (Taylor and Barker 1983; Wiegand et al. 1995). From the Bacillaceae, another type of glycosidase inhibitor, 1-deoxynojirimycin (a monosaccharide), was developed and, in form of a synthetic derivative, 3,4,5-piperidinetriol,1-(2-hydroxyethyl)-2-(hydroxymethyl)-[2R-(2a,3b,4a,5b)]-deoxynojirimycin, marketed under the trade name Miglitol (Mueller 1989).
Detection, use and structural features of C7-aminocyclitol-containing glycosidase inhibitors
The first published group of C7-cyclitol-containing glycosidase inhibitors were the validamycins/validoxylamines obtained from Streptomyces hygroscopicus ssp. limoneus, having antibiotic activity against fungi and acting as trehalase inhibitors (Iwasa et al. 1970; Asano et al. 1990; Fig. 1B). The acarbose-related metabolites were first found at about the same time (priority date of the first patent on these inhibitors is 28 December 1970; see Frommer et al. 1975), in a screening of many strains of various actinomycete genera for their activity as inhibitors of three mammalian intestinal glycosidases: α-amylase, saccharase, and maltase (Frommer et al. 1975, 1977a, 1977b; Truscheit et al. 1981; Mueller 1989). The same and other metabolites of the same group and further activities, e.g. against trehalases, were detected by various groups, mainly in streptomycete producers: amylostatins (Murao 1975; Murao and Ohyama 1975), oligostatins (Itoh et al. 1981), adiposins (Namiki et al. 1982), trestatins (Yokose et al. 1983) and 5,6-epoxy-C7-cyclitol-containing derivatives of amylostatins (NS complex; Takeda et al. 1983). Acarbose, O{4,6-dideoxy-4[1s-(1,4,6/5)-4,5,6-trihydroxy-3-hydroxymethyl-2-cyclohexen-1-yl]-amino-α-d-glucopyranosyl}-(1→4)-O-α-d-glucopyranosyl-(1→4)-d-glucopyranose, is structurally an unusual pseudotetrasaccharide, analogous to maltotetraose, and is a component of a complex mixture of shorter or longer components, also called amylostatins (Fig. 1A). Its core structure is the pseudodisaccharide acarviosine (valienaminyl-4-amino-4,6-dideoxyglucose), which is α-1,4-bound to a maltose residue (Fig. 1A). In general, these compounds can be regarded as members of the class of aminocyclitol-containing aminoglycosides which, with few exceptions, are all produced by actinomycetes (Piepersberg 1997; Piepersberg et al. 2002).

Structure of acarbose (A), validamycins (B) and related natural products
The mode of action of the inhibitory activity of acarbose on several α-glucosidases and cyclodextrin glucanotransferases has been studied, e.g. in crystals of enzyme-inhibitor complexes (e.g. Qian et al. 1994; Strokopytov et al. 1995; Gilles et al. 1996; Machius et al. 1996). The acarviosine moiety binds with high affinity to the active centers of the proteins in this enzyme family (Brayer et al. 2000). Obviously, various modes of binding and cleavage and subsequent transglycosylation with two or more acarbose molecules can also occur on α-amylases and related members of the COG family of enzymes (Tatusov et al. 2001). Transglycosylation creates elongated derivatives with two or more acarviosyl residues, both in the presence and absence of additional maltooligosaccharides, and forms inhibitors with much stronger affinity to the enzyme molecule. In α-amylases, these efficient inhibitor molecules bind mostly with one of their aminoglycosidic bonds, which cannot be hydrolytically split, in the hydrolase center and thereby block the enzyme almost irreversibly. Thus, acarbose acts at least in some of the blocked hydrolases as a prodrug.
The α-glucosidase inhibitor acarbose (Glucobay) is now marketed worldwide (launched in 1990 in Germany, launched in the USA in 1995) and is used in the therapy of diabetes type II (non-insulin-dependent), in order to enable patients to better control blood sugar contents while living with starch-containing diets (Ahr et al. 1989; Bischoff et al. 1994). It was selected from the amylostatin complex because of its higher inhibitory activity towards intestinal sucrase, maltase, dextrinase and glucoamylase, relative to its weaker inhibition of α-amylase. These properties, together with the specific pharmacokinetics and toxicology of the substance, make it an ideal therapeutic. In short, since the membrane-bound oligo- and disaccharidases at the surface of the intestinal microvilli are mainly responsible for the absorption of glucose into the blood, acarbose is preferred relative to the higher order amylostatins. Acarbose acts as a competitive inhibitor, without interfering with the glucose uptake directly. Only very little acarbose (1–4%) is absorbed itself and it practically does not penetrate into tissue. Instead, this amount is secreted rapidly via the kidneys. Also, the main fraction remaining in the intestinal tract stays mostly unaltered, with the occurrence of some microbial metabolism and conversion of intestinal glucosidases. This latter effect is in part due to the fact that e.g. pancreatic α-amylase converts acarbose to the longer products mentioned above. In general, acarbose behaves like other hardly digestible polysaccharides and has no detectable toxicity. The overall effect of this on the diabetic patient is that the starch-containing diet is degraded more slowly and along the full length of the small intestine. Thus, in type II diabetics the post-prandial increase of blood glucose levels is significantly reduced. In man, a dose of 1–1.5 mg acarbose/kg body weight reduces the post-prandial hyperglycemia after ingestion of a carbohydrate-containing diet by 50% and can help to avoid glucosuria and other deleterious effects (Truscheit et al. 1988).
Technology of acarbose production
Acarbose-type compounds were found in a screening at Bayer AG in a program designed for the detection of inhibitors for mammalian intestinal α-amylase, sucrase and maltase activities among the products of various actinomycetes (first claimed in a patent in 1970; Frommer et al. 1975). The active ingredient of Glucobay, a weakly basic acarbose, was produced in a multi-step batch fermentation with developed strains derived from Actinoplanes sp. SE50 (CBS 961.70; ATCC 31042) and by use of feeding protocols. A multi-stage purification process was carried out at the Bayer plant at Wuppertal–Elberfeld (Fig. 2). The sterile fermentation took place in 30–100 m3 fermenters in a high-concentration medium, mainly based on starch hydrolysates (10% or higher) with or without added maltose (low glucose levels) under continuous control and applying a feed of nutrients to keep the main C-source (poly-, oligomaltodextrins) at a high concentration throughout (Frommer et al. 1977a; Rauenbusch and Schmidt 1978; Beunink et al. 1997). The basic downstream processing of the industrial production process involved seven steps:

Schematic flow-sheet for the large-scale production of acarbose (according to Bayer AG, brochure PH-TO/WP, PH 5)
-
1.
The ion content of the culture broth was lowered by mixing it with both a highly acidic cation exchanger and an anion exchanger. Under this condition, most (more than 80%) of the acarbose bound to the cation exchanger.
-
2.
The resins were separated from the broth and mycelia by passing the mixture through a sieve screw centrifuge, in which a washing step (deionized water) also took place.
-
3.
Elution by 0.1 M sodium acetate occurred in kettles containing a suction head with nozzle sieves in the bottom, connected via a pump and rotameter to a column line.
-
4.
Both inhibitor and contaminants were adsorbed and thereby separated via a column line of three ion-exchange columns (cation/anion/cation exchangers in acidic/basic/acidic forms, respectively) of which the first was used for desalination, the second for elevating the pH to higher than 3.0 and the third for binding the acarbose and its related components.
-
5.
The last column was eluted by a gradient of 0.01–0.05 M mineral acids and a fractionated collection was made of the different components, the acarbose fraction of which was cut off.
-
6.
The final creation of a slightly colored powder was achieved by volume reduction in a vacuum and the pH was brought to a value of about pH 5–7 by a mixture of cation and anion exchange resins, filtration through a sterile filter and freeze- or spray-drying (Rauenbusch and Schmidt 1978).
-
7.
Later, several improvements of the fermentation process and downstream processing were published, e.g. replacement of step 6 of the above procedure by an upscale of a quasi-HPLC procedure (e.g. columns of 11 m3 bed volume, 2.5 m diameter, 3 m height) via the development of a sulfonated bead cation exchanger using specially developed macroporous, hard and resistant polymers based on aromatic compounds with at least one vinyl group and one hydrophilic monomer (e.g. 6–20% divinylbenzene, 5-25% methoxymethacrylamide, among others; Lange and Rauenbusch 1986). This allowed a more rapid flow and smaller elution volumes, combined with a stronger separation.
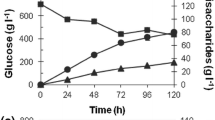
Other improvements included: (a) the refined purification and separation of the acarbose fraction from the related components and other impurities after step 6 of the above procedure, using a special weakly acidic cation exchanger based on carboxymethyl and yielding about 98% pure acarbose (Rauenbusch 1987) and (b) controlling the osmolality of the fermentation broth at an optimal value around a relatively sharp optimum of 400 mosmol/kg. This was achieved by feeding protocols or controlling the nutrient and salt supply in continuous culture (Beunink et al. 1997), both lower and higher osmolalities having severely negative effects. No exact values for the final acarbose concentrations in the harvested broth of the fermentation have been reported for the actual production strains. The deposited derivative Actinoplanes sp. SE50/110 produces up to 1 g acarbose/l under optimal culture conditions (Rauenbusch and Schmidt 1978). As a side-product, with increasing relative concentration during the elongated fermentation, component C (cf. Fig. 1) seems to be unavoidable. Due to its high structural similarity to acarbose, component C is hard to separate. The biotechnological production process requires 2–3 weeks; and the purified product still contains some contamination by component C (below 2%).
Physiology and biogenetics of acarbose formation in the producer, Actinoplanes sp. SE50
During the development of derivatives of Actinoplanes sp. SE50 for production, it soon became apparent that glucose concentration (negative control) and, relative to that, maltose and maltooligodextrin contents up to higher order hydrolysates of starch (positive control) play a key role in determining the productivity (Frommer et al. 1975; Frommer et al. 1977a, 1977b; Rauenbusch and Schmidt 1978). Further evidence for direct positive regulation (induction) by maltooligosaccharides, especially by maltotriose, came from the finding of carbohydrate-regulated operator sites, the so-called dyadic symmetry element (DSE) boxes, upstream of some of the key acarbose production genes, namely acbA, acbB, acbD and acbE, which are pairwise transcribed (acbA with acbB, acbD with acbE) from two common, divergently oriented promoter regions (see below; Stratmann 1997). The acbD and acbE genes, when heterologously expressed in strains of S. lividans 66, were largely inducible by maltotriose. Similar putative DSE-type operator sites mediating induction and glucose repression were reported for α-amylase genes in different Streptomycetes strains; and a pleiotropic regulator gene, reg1, exists in S. lividans 66, where it potentially could act heterologously on acb genes (Virolle and Gagnat 1994; Nguyen et al. 1997).
The biogenesis of the valienamine moiety and the acarbose molecule was investigated during the 1980s by feeding studies with various labeled precursors (Fig. 3) in whole-cell experiments with strains derived from Actinoplanes sp. SE50 (Degwert et al. 1987; Rinehart et al. 1992; Lee et al. 1997; Lee and Egelkrout 1998; Mahmud et al. 1999; Mahmud 2003). Biogenetic studies were conducted from the starting hypothesis that the C7N moiety of acarbose could be introduced via valienamine (cf. Fig. 3) as an intermediate and use a shikimate pathway for its biogenesis. This was tested first, but found to be not the case. Instead, a cyclization of a heptulosephosphate derived from transketolase reactions was proposed. The amino-N was shown to be most likely introduced from l-glutamate. Surprisingly, the labeled C7-cyclitol precursors which could be introduced into the valienamine moiety of validamycin (Dong et al. 2001; Mahmud et al. 2001) could not be incorporated into acarbose, except for one compound, 2-epi-5-epi-valiolone, which is the starting precursor of the biosynthetic pathway and is derived from sedo-heptulose-7-phosphate (see below). In contrast, the presumed C7-cyclitol intermediates 5-epi-valiolone, valienone and validone are efficiently incorporated into validamycin in its producer, S. hygroscopicus ssp. limoneus. The maltose unit in acarbose was shown to be directly incorporated from maltose or maltotriose, rather than via successive addition of glucose residues.

Chemical structures of C7-cyclitols used in studies of the biosynthesis of acarbose and validamycin
The enzymology of acarbose metabolism in the producer Actinoplanes sp. SE50 was extensively investigated by Pape and coworkers (Drepper and Pape 1996; Drepper et al. 1996; Goeke et al. 1996; Hemker 1997; Hemker et al. 2001; Stolpe 2001), who showed that an intracellular phosphotransferase phosphorylated the C7-hydroxyl of the cyclitol moiety (acarbose-7-kinase), and an unusual intracellular maltose-1-kinase and an acarviosyltransferase which (α-1,4)-transglycosylated the acarviosyl residue to several sugars and oligosaccharides with a glucose-like configuration around the receptor 4-OH group. The acarviosyltransferase first was found cell-attached, but was later also found in the fermentation supernatant. Some of this enzymology could also be attributed to encoding genes in the acb cluster (see below).
Genetics of acarbose metabolism in the producer Actinoplanes sp. SE50
For the isolation of gene clusters of polyketides in actinomycetes, well established and highly conserved tag sequences which have existed since the mid-1980s can now efficiently be used in heterologous PCR protocols. The findings that many antibiotics and other secondary metabolites contain 6-deoxyhexoses (6DOHs; which are made via the dTDP-glucose pathway) and that the key enzyme, dTDP-d-glucose 4,6-dehydratase, is also highly conserved allowed the screening of DNA libraries with the respective probes (Stockmann and Piepersberg 1992; Piepersberg 1994). Studies on the genetics of amylostatin (acarbose) production were the first to be started using heterologous PCR in screening and analyzing DNA-fragments around the dTDP-d-glucose 4,6-dehydratase-encoding genes in the two organisms Actinoplanes sp. SE50 [mainly in strain SE50/110 (CBS 674.73, ATCC 31044); Crueger et al. 1996, 1997; Stratmann 1997; Stratmann et al. 1999] and S. glaucescens GLA.0 (Decker 1997; Fig. 4; Table 1). This method also proved to be useful in many other cases (e.g. Decker et al. 1996). Testing other gene sequences commonly observed in secondary metabolic gene clusters, such as the secondary metabolic aminotransferase (SMAT) members which occur as aminocyclitol aminotransferases (Piepersberg 1994; Ahlert et al. 1997; Ota et al. 2000; Tamegai et al. 2002), failed to identify the acb cluster in Actinoplanes sp. SE50/110. This was later explained by the fact that the biosynthetic aminotransferase of the acarbose pathway, AcbV, is a member of another aminotransferase family (see below; Table 1).

The acb cluster of Actinoplanes sp. SE50 (Apeler et al. 2001)
Of the proteins encoded by the six genes analyzed in the partial acb gene cluster from S. glaucescens GLA.0, only the AcbA(Sgl) and AcbB(Sgl) proteins showed a strong conservation relative to the homologous proteins AcbW(Asp) and AcbV(Asp), respectively, from Actinoplanes sp. SE50. This could mean that gene clusters and production abilities for amylostatin-like compounds are both old and widespread features in the Actinomycetales. However, related gene sets do not occur in all strains; e.g. they are absent from the full-length genomes of S. coelicolor A3(2) and S. avermitilis (Piepersberg 2001; Bentley et al. 2002; Ikeda et al. 2003). Evidence for the functionality of these acb clusters to represent the real production genes for the compounds of the amylostatin complex stems from: (1) the enzymology of individual Acb proteins (see below), (2) mutant studies (Decker 1997; W. Wohlleben, personal communication) and (3) the expression of a cosmid containing the full-length acb cluster of Actinoplanes sp. SE50 in S. lividans 66 strain TK23 resulting in acarbose production in this heterologous organism (Wehmeier et al., unpublished data).
The acb gene cluster from the producer strain Actinoplanes sp. SE50 has been fully analyzed (Fig. 4; Table 1). The 25 acb genes are organized in several transcription units (at least eight units, three of which are operons). They encode a unique mixture of functions of which, besides biosynthetic enzymes, more than one-third should be involved in α-glucoside metabolism and transport, according to their similarity to known proteins. Other known or postulated functions of Acb proteins prove the existence of a characteristic biosynthetic pathway for the C7-cyclitol and 6DOH moieties and their condensation in the formation of the acarviosyl residue of acarbose (cf. Fig. 1A). Especially surprising is the fact that, besides cytoplasmically localized anabolic enzymes and export systems, as usually combined in the biosynthetic gene clusters for other secondary metabolites, catabolic (both cytoplasmic and extracellular) and import systems also seem to participate in this metabolic compartment. These puzzling observations can be interpreted in a hypothetical metabolic cycle for the acarbose-like compounds and its competitive use for the removal of the C-source (α-glucosides) from the surrounding environment of the producing cells. The known or hypothetical functions of the individual gene sets are described in more detail in the following section.
The putative biosynthetic genes acbAB and acbVUSRPIJQKMLNOC
The two genes acbA(Act) and acbB(Act) for the two first steps of 6-deoxyhexose formation (see above; Liu and Thorson 1994; Piepersberg 1994) are not located in tandem in an operon as usual, but in two adjacent and divergently oriented transcription units (cf. Fig. 4). They might be transcriptionally controlled (via so-called DSE sequences as operators) by a Reg1-like regulatory protein (Virolle and Gagnat 1994; Stratmann 1997). Their organization in the cluster and sequence similarities deviate remarkably from those observed for the equivalent genes from S. glaucescens GLA.0: acbC(Sgl) and acbD(Sgl). The genes acbVUSRPIJQKMLNOC, all oriented in the same direction, could form one or several operons. Here, for practical reasons, we treat them as two groups, acbQKMLNOC and acbVUSRPIJ, which are presumed to be mainly involved in cyclitol biosynthesis (besides intracellular metabolism, the “upper pathway”) and deoxysugar and condensation pathway (the “lower pathway”), respectively. The gene acbQ encodes a protein which has significant similarity throughout the full-length polypeptide chain with that of the amylomaltase MalQ of H. influenzae (L45989). This enzyme is a typical constituent of the intracellular metabolism of maltose and other maltooligodextrins. The genes acbK, acbC, acbM, acbO and acbL encode the enzymes acarbose-7-kinase, 2-epi-5-epi-valiolone synthase (C7-cyclitol cyclase), 2-epi-5-epi-valiolone 7-kinase, 2-epi-5-epi-valiolone-7-phosphate 2-epimerase and putatively 2-epi-valiolone-7-phosphate 1-reductase, respectively, as was shown by in vitro enzyme assays with the individual purified enzymes, heterologously expressed in strains of S. lividans 66 (Stratmann et al. 1999; Zhang et al. 2002, 2003a, 2003b). The protein product of the acbK gene has acarbose 7-phosphotransferase activity. In primary structure, the acarbose-7-kinase protein resembles other members of a family of sugar kinases, such as fructo- and ribokinases and, interestingly, it shows the highest degree of similarity to an unknown protein (57.9% identity in a 297-amino acid overlap) encoded by a gene which lies adjacent to an α-amylase gene in a Streptomyces sp. (accession number U08602). The AcbM polypeptide chain shows distant similarity to some members of the hexokinase family of sugar phosphotransferases, which form a cluster of orthologues in complete genome sequences: COG1940 (Tatusov et al. 2001). This similarity is intriguing, since both these enzymes, d-glucose 6-kinase (GlcK) and 2-epi-5-epi-valiolone 7-kinase (AcbM) phosphorylate the primary hydroxyl group on a cyclic polyol with a six-member ring system. The AcbL and AcbN proteins belong to two different families of the dinucleotide-binding oxidoreductases. The AcbL protein shows high similarities to zinc-dependent alcohol dehydrogenases and AcbN is a member of the family of short-chain dehydrogenases (cf. Table 1). AcbO shows no significant similarity to any known enzyme family. The protein product of the acbC gene is distantly similar to the AroB proteins encoded by the respective dehydroquinate synthase genes in various bacterial organisms (Stratmann et al. 1999).
The genes for extracellular α-glucoside and acarbose metabolism, acbD, acbE, acbZ
The two genes acbD and acbE form another set of divergent mono-cistronic transcription units sharing a common promoter region (cf. Fig. 2). The protein products of the acbD, acbE and acbZ genes exhibit typical signal peptide motifs at their N-termini and show strong similarity to members of the α-amylase family of proteins, which are mostly extracellular α-glucosidases or α-glucoside transglycosylases (e.g. cyclodextrin synthases). Especially, the catalytic triad of acidic residues (D–E–D) is also conserved in the equivalent positions of the AcbD, AcbE and AcbZ polypeptide chains.
The putative export and import genes, acbWXY and acbFGH
These two operons, each consisting of three genes, obviously form common transcription units. Both encode ABC-type transporter systems, however of completely different structure and functionality (see below). The proteins encoded by the acbFGH genes represent the trans-membrane and extra-cytoplasmic components of a typical binding protein-dependent ABC transporter, with the highest degree of similarity to the MsmEFG importer of S. mutans (Russell et al. 1992) and the MalEFG maltodextrin importers of enterobacteria (Boos and Lucht 1996). However, a gene for a cytoplasmic ATP-binding component, similar to MalK or MsmK and typical for this type of ABC-sugar importers (Boos and Lucht 1996), is lacking in the acb gene cluster, since unrelated genes are located upstream of the acbHFG operon of Actinoplanes sp. (our unpublished observations). Thus, one can speculate that a protein homologous to the general ATP-binding protein MsiK (Hurtubise et al. 1995), which was found to be involved in a number of ATP-dependent transport systems in streptomycetes, is the ATP-binding subunit of the AcbFGH transport system. The presence of genes for a putative sugar transporter in the vicinity of those for the metabolism of acarbose-like products in the genome of Actinoplanes sp. suggests that there is a strong metabolic link between this secondary metabolite and the import of extracellular carbohydrates (see below).
Proposal of a biosynthetic pathway for acarbose
Our proposal for the biosynthetic pathway for acarbose is shown in Fig. 5 and is described here only briefly. We have already mentioned that 2-epi-5-epi-valiolone is the first precursor for the cyclitol moiety of acarbose. Further conversion of the cyclitol precursor, 2-epi-5-epi-valiolone-7-phosphate, is catalyzed by the epimerase AcbO. The 2-epi-5-epi-valiolone-7-phosphate-epimerase AcbO catalyzes the formation of 5-epi-valiolone-7-phosphate (Zhang et al. 2003a). The AcbO reaction product, 5-epi-valiolone, has been characterized by mass-spectroscopic and NMR-spectroscopic methods. The enzyme AcbO is active without any cofactor and the deduced protein exhibits no similarities with any known epimerases so far. Therefore, AcbO is obviously the first representative of a new class of epimerases (Zhang et al. 2003a). Recent experiments gave first hints that 5-epi-valiolol-7-phosphate is the next intermediate obtained after reduction at the C-1 keto-group, catalyzed by a NADH-dependent dehydrogenase, AcbL (Wehmeier and Zhang, unpublished data). The next steps in the biosynthesis are speculative at the moment. The AcbN protein could catalyze the dehydratase reaction necessary for the synthesis of 1-epi-valienol-7-phosphate. A phosphorylation at C-1 (the respective enzyme has not yet been identified) would be a prerequisite for a subsequent nucleotidylation step. In the acb cluster, we identified the gene acbR, which encodes GlgC (COG0448; Tatusov et al. 2001), an ADP–glucose synthase-like protein, which could catalyze the nucleotidylation of the likely precursor, 1-epi-valienol-1,7-diphosphate.

Postulated pathway of acarbose biosynthesis in Actinoplanes sp. SE50 (Zhang et al. 2002)
The 7-phosphorylation of the C7-cyclitol moiety during the de novo synthesis of acarbose might be necessary to prevent an inhibitory effect of the C7-cyclitol- and acarviosyl-containing intracellular metabolites on cytoplasmic enzymes of the producer sensitive to those inhibitors, e. g. α-glucosidases or glucomaltases. For instance, after 7-phosporylation of acarbose by AcbK, the modified inhibitor does not inhibit the cytoplasmic and acarbose-sensitive amylomaltase activity of Actinoplanes sp. SE50/110 (Drepper and Pape 1996). Therefore, this phosphorylation can be regarded as a resistance-like self-protection mechanism similar to those in other aminoglycoside producers (Piepersberg and Distler 1997; He and Liu 1997; Piepersberg et al. 2002).
Recently, a new kinase activity in Actinoplanes was characterized which phosphorylates 1-epi-valienol to 1-epi-valienol-7-phosphate (and also displays weak activity with valienol; Zhang et al. 2003b). In early fermentation phases with derivatives of Actinoplanes sp. SE50 (up to 30 h growth), 1-epi-valienol and valienol were found in the supernatant; but thereafter the concentration of these intermediates in the supernatant decreased again (Mahmud et al. 1999). Therefore, the 1-epi-valienol-7-kinase activity might play a role in a salvage pathway which could reintroduce these cyclitols to the biosynthetic route (not shown in Fig. 5). This kinase activity could not be attributed to any gene of the acb gene cluster, since S. lividans strains harboring the cosmid pHTWCos6 (including all acb genes, as shown in Fig. 4; Thomas 2001) did not exhibit this 1-epi-valienol-7-kinase activity. Therefore, this enzyme is probably not encoded by the acb gene cluster (Zhang et al. 2003b).
The synthesis of the deoxysugar moiety of acarbose follows the 6DOH route mentioned above (Liu and Thorson 1994; Piepersberg and Distler 1997). It starts with glucose-1-phosphate, which is first activated by a nucleotidylation step catalyzed by a dTDP-glucose synthase (AcbA) and then further modified by a dTDP-glucose-4,6-dehydratase (AcbB) which leads to dTDP-4-keto-6-deoxy-glucose. dTDP-4-keto-6-deoxy-glucose is the substrate for the subsequent aminotransferase reaction. The aminotransferase AcbV is a member of the class III aminotransferases. When the protein was heterologously expressed in S. lividans 66, it catalyzed an l-glutamic acid-dependent amination of dTDP-4-keto-6-deoxy-d-glucose, probably resulting in dTDP-4-amino-4,6-dideoxy-d-glucose (Diaz-Guardamino Uribe 2000; Piepersberg et al. 2002). Interestingly, the protein AcbV is related to aminotransferases of the primary metabolism (being a member of the GabT-like aminotransferases) and it does not belong to the SMATs (Piepersberg 1994, 1997), which are normally found in the biosynthetic pathways of antibiotics and other secondary metabolites (Piepersberg 1997; Piepersberg and Distler 1997; Piepersberg et al. 2002).
The deduced proteins AcbI and AcbS are related to glycogen and sucrose synthases. The two glycosyltransferase-like reactions postulated in the pathway (formation of the dTDP-acarviose-7-phosphate, synthesis of acarbose-7-phosphate; cf. Figs. 5, 6) thus may be catalyzed by AcbI and AcbS. However, it is not clear whether acarbose-7-phosphate or some other compound instead of acarbose, e.g. the mono- or diglucosylated acarviosine-7-phosphate, is the end-product of the cytoplasmatic pathway.

Acarbose as a possible carbophore. The possible cycling of acarbose-derived metabolites between intra- and extracellular pools is shown. Re-uptake and intracellular phosphorylation and unloading of acarbose derivatives via a specific glucomaltase activity (which have been elongated extracellularly via transglycosylation by additional glucose residues) could lead to a cyclic pool separation and net glucose uptake
Intra- and extracellular metabolism of amylostatins and maltooligosaccharides
As described above, the acb cluster encodes proteins for the biosynthetic pathway of acarbose, extracellular starch-degrading enzymes, intracellular maltose-degrading enzymes, transport systems (import/export) and a acarbose-7-kinase. All these activities are needed for a cyclic acarbose/amylostatin metabolism (Fig. 6). In this cyclic metabolism, the three extracellular proteins encoded by the acb cluster of Actinoplanes sp. SE 50/110 (AcbE, AcbZ, AcbD) allow the degradation of starch-containing substrates and the subsequent transglucosylation of acarbose to form higher-order products (AcbE, AcbZ); and the transfer of acarviosine residues to maltose or higher maltooligosaccharides catalyzed by AcbD forms elongated species relative to acarbose. The formation of the higher-glucosylated components of the amylostatin complex yields the mixture of products observed in culture supernatants. This mixture of products includes some acarbose-related compounds, especially component C, which is not synthesized by AcbD and does not serve as a substrate for the enzyme (Hemker 1997, Hemker et al. 2001). It was also shown that the formation of component C is not catalyzed by an extracellular enzyme. Therefore, it is most likely a product of an enzyme which is released from lysed cells during the late fermentation phases. The enzymes of trehalose metabolism, which were identified in Actinoplanes sp., are probably involved in the formation of component C, which is obviously formed from acarbose (Stolpe 2001). The combination of an active export (promoted by AcbXYZ), with dephosphorylation and conversion by α-1,4-glucosyltransferases, such as α-amylases and related proteins (AcbE, AcbZ, AcbD), re-uptake by a specific importer (AcbFGH with MsiK), rephosphorylation by AcbK and deglucosylation by an amylomaltase-equivalent enzyme (AcbQ), could form a cycle. In Actinoplanes sp., a maltase activity can be detected in the cell-free extract (Pape and collaborators, unpublished data). The identity of the measured enzyme(s) and its correlation with the AcbQ gene product has not yet been demonstrated. The enzyme was sensitive to acarbose but not to acarbose-7-phosphate, an observation that fits exactly to our model.
Hypotheses on the functions of acarbose-like alpha-glycosidase inhibitors in nature: are they “carbophores”?
It is hard to imagine how the production of an extracellular inhibitor of α-glucosidases, such as extracellular α-amylases, could facilitate the life of a bacterium in an ecological niche rich in C-sources such as starch and other α-1,4-glucans, if an inhibitory action on hydrolases is the only function provided. Therefore, additional functions have to be envisaged, e.g. selective inhibition of maltose- or maltodextrin-uptake systems in competitors. First experiments have shown that the α-amylase activity of Bacillus subtilis strains on starch plates is strongly inhibited when the strains are grown together with Actinoplanes on the same plate (Wehmeier and Rockser, unpublished data). This result is supported by the finding that the Escherichia coli maltose importer (MalEFGK-system) is specifically blocked by acarbose, which strongly binds to the periplasmic binding protein, MalE (Brunkhorst et al. 1999). Also, the fact that the α-amylase AcbE is not inhibited by acarbose supports this hypothesis, since an extracellular degradation of starch-like nutrients in the environment of the producer Actinoplanes sp. SE50 would also feed other cells, particularly in the soil habitat where it was isolated.
From all these observations, one could suggest further ecological functions for this remarkable group of secondary metabolites: (1) a trap for extracellular glucose and (2) tagging of extracellular glucose pools for import.
Thus, in the cycle described above, acarbose or its core subunit plays a multifunctional role as both a sink for glucose-containing C-sources and a transport vehicle (Fig. 6). This could elegantly explain how acarbose-like compounds facilitate the life of microorganisms in a community competing for the same C-sources as major nutrients; and the scheme could be extended to similar structures, such as trestatins or validamycins. An intracellular maltooligosyltrehalose synthase activity was found in several actinomycetes and in other bacterial genera; and a gene was also detected in Actinoplanes sp. SE50 which could be responsible for its production (Kim et al. 2000; Stolpe 2001).
References
Ahlert J, Distler J, Mansouri K, Piepersberg W (1997) Identification of the gene, stsC, encoding the l-glutamine:scyllo-inosose aminotransferase from streptomycin-producing streptomycetes. Arch Microbiol 168:102–113
Ahr HJ, Boberg M, Krause HP, Maul W, Mueller FO, Ploschke HJ, Weber H, Wuensche C (1989) Pharmacokinetics of acarbose. Part I: absorption, concentration in plasma, metabolism and excretion after single administration of [14C]acarbose to rats, dogs and man. Arzneimittelforschung 39:1254–1260
Apeler H, Wehlmann H, Piepersberg W, Diaz-Guardamino P-M, Jarling M, Thomas H, Wehmeier U (2001) Neue Enzyme in der Acarbose-Synthese und deren Verwendung. German patent DE-OS 10021667
Asano N, Kameda Y, Matsui K, Horii S, Fukase H(1990) Validamycin H, a new pseudo-tetrasaccharide antibiotic. J Antibiot (Tokyo) 43:1039–1041
Bentley SD, Chater KF, Cerdeno-Tarraga AM, Challis GL, Thomson NR, James KD, Harris DE, Quail MA, Kieser H, Harper D, Bateman A, Brown S, Chandra G, Chen CW, Collins M, Cronin A, Fraser A, Goble A, Hidalgo J, Hornsby T, Howarth S, Huang CH, Kieser T, Larke L, Murphy L, Oliver K, O’Neil S, Rabbinowitsch E, Rajandream MA, Rutherford K, Rutter S, Seeger K, Saunders D, Sharp S, Squares R, Squares S, Taylor K, Warren T, Wietzorrek A, Woodward J, Barrell BG, Parkhill J, Hopwood DA (2002) Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 417:141–147
Beunink J, Schedel; M, Steiner U (1997) Osmotically controlled fermentation process for the preparation of acarbose. German patent DE 19637591 (US patent 6,130,072)
Bischoff H, Ahr HJ, Schmidt D, Stoltefuss J (1994) Acarbose—ein neues Wirkprinzip in der Diabetestherapie. Nachr Chem Tech Lab 42:1119–1128
Boos W, Lucht JM (1996) Periplasmic binding protein-dependent ABC transporters. In: Neidhardt FC (ed) Escherichia coli and Salmonella, 2nd edn. ASM Press, Washington D.C., pp 1175–1209
Brayer GD, Sidhu G, Maurus R, Rydberg EH, Braun C,| Wang Y, Nguyen NT, Overall CM, Withers SG (2000) Subsite mapping of the human pancreatic r-amylase active site through structural, kinetic, and mutagenesis techniques. Biochemistry 39:4778–4791
Brunkhorst C, Andersen C, Schneider E (1999) Acarbose, a pseudooligosaccharide, is transported but not metabolized by the maltose-maltodextrin system of Escherichia coli. J Bacteriol 18:2612–2619
Crueger A, Piepersberg W, Distler J, Stratmann A (1996) Acarbose biosynthesis genes from Actinoplanes sp., process for the isolation thereof and the use thereof. German patent DE 19507214.6 (US patent 5,753,501)
Crueger A, Dellweg H-G, Lenz J, Schroder, Pape H, Goeke K, Schaper B, Hemker M, Piepersberg W, Distler J, Stratmann A (1997) Processes for preparing acarviosyl transferase and for using it in the conversion of acarbose homologues into acarbose, for the preparation of acarbose homologues. German patents DE 19611252, 19625269 (US patent 5,989,882)
Decker H (1997) Isolation of the biosynthesis genes for pseudo-oligosaccharides from Streptomyces glaucescens GLA.O, and their use. German patent DE 19622783 (US patent 6,306,627)
Decker H, Gaisser S, Pelzer S, Schneider P, Westrich L, Wohlleben W, Bechthold A (1996) A general approach for cloning and characterizing dNDP-glucose dehydratase genes from actinomycetes. FEMS Microbiol Lett 141:195–201
Degwert U, Hulst R van, Pape H, Herrold RE, Beale JM, Keller PJ, Lee JP, Floss HG (1987) Studies on the biosynthesis of the alpha-glucosidase inhibitor acarbose: valienamine, am-C7N unit not derived from the shikimate pathway. J Antibiot (Tokyo) 40:855–861
Demain AL, Somkuti GA, Hunter-Creva JC, Rossmoore HW (1989) Novel microbial products for medicine and agriculture. Elsevier Science, Amsterdam
Díaz-Guardamino Uribe PM (2000) Untersuchungen zum Einbau des Stickstoffes in der Acarviose-Einheit der Acarbose bei Actinoplanes sp. 50/110: die Aminotransferase AcbV. PhD thesis, Bergische Universität Gesamthochschule, Wuppertal
Dong H, Mahmud T, Tornus I, Lee S, Floss HG (2001 ) Biosynthesis of the validamycins: identification of intermediates in the biosynthesis of validamycin A by Streptomyces hygroscopicus var. limoneus. J Am Chem Soc 123:2733–2742
Drepper A, Pape H (1996) Acarbose 7-phosphotransferase from Actinoplanes sp.: purification, properties, and possible physiological function. J Antibiot (Tokyo) 49:664–668
Drepper A, Peitzmann R, Pape H (1996) Maltokinase (ATP:maltose 1-phosphotransferase) from Actinoplanes sp.: demonstration of enzyme activity and characterization of the reaction product. FEBS Lett 388:177–179
Frommer W, Puls W, Schaefer D, Schmidt D (1975) Glycoside-hydrolase enzyme inhibitors. German patent DE 2064092 (US patent 3,876,766)
Frommer W, Junge B, Keup U, Mueller L, Schmidt D (1977a) Amino sugar derivatives. German patent DE 2347782 (US patent 4,062,950)
Frommer W, Puls W, Schmidt D (1977b) Process for the production of a saccharase inhibitor. German patent DE 2209834 (US patent 4,019,960)
Frommer W, Junge B, Mueller L, Schmidt D, Truscheit E (1979) Neue Enzyminhibitoren aus Mikroorganismen. Planta Med 35:195–217
Gilles C, Astier J-P, Marchis-Mouren G, Cambillau C, Payan F (1996) Crystal structure of pig pancreatic α-amylase isoenzyme II, in complex with the carbohydrate inhibitor acarbose. Eur J Biochem 238:561–569
Goeke K, Drepper A, Pape H (1996) Formation of acarbose phosphate by a cell-free extract from the acarbose producer Actinoplanes sp. J Antibiot (Tokyo) 49:661–663
He XM, Liu HW (2002) Formation of unusual sugars: mechanistic studies and biosynthetic applications. Annu Rev Biochem 71:701–754
Hemker M (1997) Pseudooligosaccharide und Staerkestoffwechsel bei Actinoplanes sp. PhD thesis, Westfaelische Wilhelms-Universitaet, Muenster
Hemker M, Stratmann A, Goeke K, Schroder W, Lenz J, Piepersberg W, Pape H (2001). Identification, cloning, expression, and characterization of the extracellular acarbose-modifying glycosyltransferase, AcbD, from Actinoplanes sp. strain SE50. J Bacteriol 183:4484–4492
Hurtubise Y, Shareck F, Kluepfel D, Morosoli R (1995) A cellulase/xylanase-negative mutant of Streptomyces lividans 1326 defective in cellobiose and xylobiose uptake is mutated in a gene encoding a protein homologous to ATP-binding proteins. Mol Microbiol 17:367–377
Ikeda H, Ishikawa J, Hanamoto A, Shinose M, Kikuchi H, Shiba T, Sakaki Y, Hattori M, Omura S (2003) Complete genome sequence and comparative analysis of the industrial microorganism Streptomyces avermitilis. Nat Biotechnol 21:526–531
Itoh J, Omoto S, Shomura T, Ogino H, Iwamatsu K, Inouye S, Hidaka H (1981) Oligostatins, new antibiotics with amylase inhibitory activity. I. Production, isolation and characterization. J Antibiot (Tokyo) 34:1424–1428
Iwasa T, Yamamoto H, Shibata M (1970) Studies on validamycins, new antibiotics. I. Streptomyces hygroscopicus var. limoneus nov. var., validamycin-producing organism. J Antibiot (Tokyo) 23:595–602
Kim YH, Kwon TK, Park S, Seo HS, Cheong JJ, Kim CH, Kim JK, Lee JS, Choi YD (2000) Trehalose synthesis by sequential reactions of recombinant maltooligosyltrehalose synthase and maltooligosyltrehalose trehalohydrolase from Brevibacterium helvolum. Appl Environ Microbiol. 66:4620–4624
Lange PM, Rauenbusch E (1986) Polymers for the purification of acarbose. German patent DE 3439008 (US patent 4,767,850)
Lee S, Egelkrout E (1998) Biosynthetic studies on the alpha-glucosidase inhibitor acarbose in Actinoplanes sp.: glutamate is the primary source of the nitrogen in acarbose. J Antibiot (Tokyo) 51:225–227
Lee S, Sauerbrei B, Niggemann J, Egelkrout E (1997) Biosynthetic studies on the alpha-glucosidase inhibitor acarbose in Actinoplanes sp.: source of the maltose unit. J Antibiot (Tokyo) 50:954–960
Liu H-W, Thorson JS (1994) Pathways and mechanisms in the biogenesis of novel deoxysugars by bacteria. Annu Rev Microbiol 48:223–256
Machius M, Vértesy L, Huber R, Wiegand G (1996) Carbohydrate and protein-based inhibitors of porcine pancreatic α-amylase: structure analysis and comparison of their binding characteristics. J Mol Biol 260:409–421
Mahmud T (2003) The C7N aminocyclitol family of natural products. Nat Prod Rep 20:137–166
Mahmud T, Tornus I, Engelkrout E, Wolf E, Uy C, Floss HG, Lee S (1999) Biosynthetic studies on the α-glucosidase inhibitor acarbose in Actinoplanes sp.: 2-epi-5-epi-valiolone is the direct precursor of the valienamine moiety. J Am Chem Soc 121:6973–6983
Mahmud T, Xu J, Choi YU (2001) Synthesis of 5-epi-[6-(2)H(2)]valiolone and stereospecifically monodeuterated 5-epi-valiolones: exploring the steric course of 5-epi-valiolone dehydratase in validamycin A biosynthesis. J Org Chem 66:5066–5073
Mueller L (1989) Chemistry, biochemistry and therapeutic potential of microbial a-glucosidase inhibitors. In: Demain AL, Somkuti GA, Hunter-Creva JC, Rossmoore HW (eds) Novel microbial products for medicine and agriculture. Elsevier Science, Amsterdam, pp 109–116
Murao S (1975) Microbial amylase inhibitor and preparation thereof with the use of Streptomyces diasticus var. amylostaticus. Japanese patent 49-29646 (US patent 4,010,258)
Murao S, Ohyama K (1975) New amylase inhibitor (S-1) from Streptomyces diasticus var. amylostaticus no. 2476. Agric Biol Chem 39:2271–2273
Namiki S, Kangouri K, Nagate T, Hara H, Sugita K, Omura S (1982) Studies on the alpha-glucoside hydrolase inhibitor, adiposin. I. Isolation and physicochemical properties. J Antibiot (Tokyo) 35:1234–1236
Nguyen J, Francou F, Virolle MJ, Guérineau M (1997) Amylase and chitinase genes in Streptomyces lividans are regulated by reg1, a pleiotropic regulatory gene. J Bacteriol 179:6383–6390
Ota Y, Tamegai H, Kudo F, Kuriki H, Koike-Takeshita A, Eguchi T, Kakinuma K (2000) Butirosin-biosynthetic gene cluster from Bacillus circulans. J Antibiot 53:1158–1167
Piepersberg W (1994) Pathway engineering in secondary metabolite-producing actinomycetes. Crit Rev Biotechnol 14:251–285
Piepersberg W (1997) Molecular biology, biochemistry, and fermentation of aminoglycoside antibiotics. In: Strohl WR, (ed) Biotechnology of industrial antibiotics, 2nd edn. Marcel–Dekker, New York, pp 81–163
Piepersberg W (2001) Endogenous antimicrobial molecules: an ecological perspective. In: Sussman M (ed) Molecular medical microbiology. Academic Press, London, pp 561–583
Piepersberg W, Distler J (1997) Aminoglycosides and sugar components in other secondary metabolites. In: Rehm H-J, Reed G, Pühler A, Stadler P (eds) Biotechnology. (Products of secondary metabolism, vol 7) VCH, Weinheim, pp 397–488
Piepersberg W, Diaz-Guardamino PM, Stratmann A, Thomas H, Wehmeier UF, Zhang CS (2002) Recent developments in the biosynthesis and regulation of aminoglycosides. In: Fierro F, Martín JF (eds) Microbial secondary metabolites: biosynthesis, genetics and regulation. Research Signpost, Kerala, pp 1–26
Puls W, Keup U (1973) Influence of an α-amylase inhibitor (BAY d7791) on blood glucose, serum insulin and NEFA in starch loading test in rats, dogs and man. Diabetologia 9:97
Puls W, Keup U, Krause HP, Thomas G, Holtmeier F (1977) Glucosidase inhibition: a new approach to the treatment of diabetes, obesity and hyperlipoproteinaemia. Naturwissenschaften 64:536–537
Qian M, Haser R, Buisson G, Duée E, Payan F (1994) The active center of a mammalian α-amylase. Structure of the complex of a pancreatic α-amylase with a carbohydrate inhibitor refined to 2.2-Å resolution. Biochemistry 33:6284–6294
Rauenbusch E (1987) Highly pure acarbose. German patent DE 3543999 (US patent 4,904,769)
Rauenbusch E, Schmidt D (1978) Verfahren zur Isolierung von (O{4,6-Dideoxy-4[[1s-(1,4,6/5)-4,5,6-trihydroxy-3-hydroxymethyl-2-cyclohexen-1-yl]-amino]-α-d-glucopyranosyl}-(1→4)-O-α-d-glucopyranosyl-(1→4)-d-glucopyranose) aus Kulturbrühen. German patent DE 2719912 (Process for isolating glucopyranose compound from culture broths; US patent 4,174,439)
Rehm H-J, Reed G, Pühler A, Stadler P (1997) Products of secondary metabolism, 2nd edn. (Biotechnology, vol 7) VCH, Weinheim
Rinehart KL Jr, Snyder WC, Staley AL, Lau RCM (1992) Biosynthetic studies on antibiotics. In: Petroski RJ, McCormick SP (eds) Secondary-metabolite biosynthesis and metabolism. Plenum Press, New York, pp 41–60
Russell R.B, Aduse-Opuko J, Sutcliffe IC, Tao L, Ferretti JJ (1992) A binding protein-dependent transport system in Streptococcus mutans responsible for multiple sugar metabolism. J Biol Chem 267:4631–4637
Stockmann M, Piepersberg W (1992) Gene probes for the detection of 6-deoxyhexose metabolism in secondary metabolite-producing streptomycetes. FEMS Microbiol Lett 90:185–190
Stolpe T (2001) Zur Bildung von Pseudooligosaccharidyl-Trehalosen durch Actinoplanes sp. PhD thesis, Westfaelische Wilhelms-Universitaet, Muenster
Stratmann A (1997) Identifizierung eines Acarbose-Biosynthesegenclusters in Actinoplanes sp. und Charakterisierung ausgewählter Enzyme des Acarbose-Stoffwechsels. PhD thesis, Bergische Universitaet, Wuppertal
Stratmann A, Mahmud T, Lee S, Distler J, Floss HG, Piepersberg W (1999) The AcbC protein from Actinoplanes species is a c7-cyclitol synthase related to 3-dehydroquinate synthases and is involved in the biosynthesis of the α-glucosidase inhibitor acarbose. J Biol Chem 274:10889–10896
Strokopytov B, Penninga D, Rozeboom HJ, Kalk KH, Dijkhuizen L, Dijkstra BW (1995) X-ray structure of cyclodextrin glycosyltransferase complexed with acarbose. Implications for the catalytic mechanism of glycosidases. Biochemistry 34:2234–2240
Takeda H, Nakagawa Y, Kiuchi A (1983) Novel amino oligosaccharide derivative. Japanese patent JP 58-172400
Tamegai H, Nango E, Kuwahara M, Yamamoto H, Ota Y, Kuriki H, Eguchi T, Kakinuma K (2002) Identification of l-glutamine: 2-deoxy-scyllo-inosose aminotransferase required for the biosynthesis of butirosin in Bacillus circulans. J Antibiot 55:707–714
Tatusov RL, Natale DA, Garkavtsev IV, Tatusova TA, Shankavaram UT, Rao BS, Kiryutin B, Galperin MY, Fedorova ND, Koonin EV (2001) The COG database: new developments in phylogenetic classification of proteins from complete genomes. Nucleic Acids Res 29:22–28
Taylor RH, Barker HM (1983) Alpha-amylase inhibitors. Lancet 1:1228–1229
Thomas H (2001) Acarbose Metabolismus in Actinoplanes sp. SE50/110. PhD, Bergische Universitaet, Wuppertal
Truscheit E, Frommer W, Junge B, Müller L, Schmidt D, Wingeder W (1981) Chemistry and biochemistry of bacterial alpha-glucosidase inhibitors. Angew Chem Int Ed Eng 20:744–761
Truscheit E, Junge B, Mueller L, Puls W, Schmidt D (1988) Microbial alpha-glucosidase inhibitors: chemistry, biochemistry and therapeutic potential. Prog Clin Biochem Med 7:17
Virolle MJ, Gagnat J (1994) Sequences involved in growth-phase-dependent expression and glucose repression of a Streptomyces α-amylase gene. Microbiology 140:1059–1067
Wiegand G, Epp O, Huber R (1995) The crystal structure of porcine pancreatic alpha-amylase in complex with the microbial inhibitor tendamistat. J Mol Biol 247:99–110
Yokose K, Ogawa K, Sano T, Watanabe K, Maruyama HB, Suhara Y (1983) New alpha-amylase inhibitor, trestatins. I. Isolation, characterization and biological activities of trestatins A, B and C. J Antibiot 36:1157–1165
Zhang CS, Stratmann A, Block O, Brueckner R, Podeschwa M, Altenbach HJ, Wehmeier UF, Piepersberg W (2002) Biosynthesis of the C(7)-cyclitol moiety of acarbose in Actinoplanes species SE50/110. 7-O-phosphorylation of the initial cyclitol precursor leads to proposal of a new biosynthetic pathway. J Biol Chem 277:22853–22862
Zhang CS, Podeschwa M, Altenbach HJ, Piepersberg W, Wehmeier UF (2003a) The acarbose-biosynthetic enzyme AcbO from Actinoplanes sp. SE 50/110 is a 2-epi-5-epi-valiolone-7-phosphate 2-epimerase. FEBS Lett 540:47–52
Zhang CS, Podeschwa M, Block O, Altenbach HJ, Piepersberg W, Wehmeier UF (2003b) Identification of a 1-epi-valienol 7-kinase activity in the producer of acarbose, Actinoplanes sp. SE50/110. FEBS Lett 540:53–57
Acknowledgements
We acknowledge excellent cooperation with the groups of A. Crueger and H. Wehlmann (Bayer AG), H. Pape (University of Muenster), H.G. Floss (University of Seattle) and H.J. Altenbach (BU Wuppertal) in this project. The work in the laboratories of the authors was supported by grants from Bayer AG (Leverkusen, Germany) and the German Ministry of Education and Research (BMBF grant BEO22-0310815).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wehmeier, U.F., Piepersberg, W. Biotechnology and molecular biology of the α-glucosidase inhibitor acarbose. Appl Microbiol Biotechnol 63, 613–625 (2004). https://doi.org/10.1007/s00253-003-1477-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-003-1477-2