
Abstract
T cells play an important role in the onset and progression of systemic lupus erythematosus (SLE), and the biases in T cell receptor beta variable (TRBV) region families and complementarity determining region three (CDR3) composition in SLE patients and mouse models have been widely reported. However, the relationship between the composition and variation in the TCR β-chain CDR3 repertoire and SLE has not been established. Here, we compared and analyzed the thymic and splenic TCR β-chain CDR3 mRNA sequences by Roche 454 high-throughput sequencing from MRL/lpr mice at different ages. Our results indicate that diversity in the TCR CDR3 repertoire from the thymus and spleen from MRL/lpr mouse was significantly decreased with increased age (disease progression) and showed a bias in usage of common TRBV and TRBJ families. The N1 region insertions in the highly expressed CDR3s significantly increased with disease progression. This study provides a new perspective for studying SLE with progression of disease in clonal level of TCR, which may provide a basis for studying the mechanism of the MRL/lpr autoreactive T cells response and tailor an individualized treatment targeting these T cells.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Systemic lupus erythematosus (SLE) manifests as a disruption of immune tolerance resulting from imbalanced humoral immunity and cellular immunity. Proliferation and activation of T cell-dependent autoreactive B cells are extensive, which produce large amounts of multiple autoantibodies that target the individual’s own tissue antigens. The numbers and function of helper and regulatory T cells in the peripheral blood of SLE patients are abnormal and are closely related to the stage and state of disease (Mok and Lau 2003).
The MRL/lpr mouse line is an inbred mouse model commonly used in SLE studies and shows a spontaneous mutation of the apoptosis-related gene Fas that affects lymphocyte apoptosis. The lpr mutation causes proliferation and activation of T cells and plays a role in the development of autoimmunity in MRL/lpr mouse (Watanabe-Fukunaga et al. 1992). Disease onset in MRL/lpr mouse is at approximately 8 weeks of age, and the disease course progresses rapidly with no gender-related differences. The average lifespan of a female mouse is 17 weeks with a mortality of 50 % at 5 months of age, (Andrews et al. 1978), and 12- to 24-week-old mice have severe lupus nephritis (Zhou et al. 2004). All MRL/lpr mice share the same MHC genetic background, and it has been shown that T cells play a role in pathogenesis by providing T cell help to autoimmune B cells (Peng and Craft 1996). It has been recently confirmed that Th1, Th2, Th17, Treg, Th22, and T follicular helper (Tfh) cells (Craft 2012; Nakashima et al. 2006; Scheinecker et al. 2010; Shin et al. 2011) play a role in the onset of SLE. Holbrook et al. (Holbrook et al. 1996) discovered biased usage of several SLE-related TRBV gene families using CDR3 spectratyping. Using RT-PCR and sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), Kolowos et al. (Kolowos et al. 1997) found a restrictive usage of certain TCR β-chain CDR3s in CD4+ T cells in peripheral blood of SLE patients. Luo et al. used CDR3 spectratyping to analyze PBMC populations from SLE patients and found biased usage of certain TRBV gene families and common CDR3 amino acid motifs in SLE patients (Luo et al. 2006). Zhou et al. studied two SLE mouse models and observed a restricted usage of TRBV2, TRBV6, TRBV8.1, TRBV10, TRBV16, and TRBV18 in the CD4+ T cells of MRL/lpr mice and restricted usage of TRBV6 and TRBV7 in the CD4+ T cells of (NZB×NZW) F1 mice. There are conserved amino acid sequences (I, D, E, and G) in the CDR3 region of these TRBV families, indicating that these clonotypes may be the T cells that recognize restricted autoantigen epitopes and may be involved in the SLE-specific autoimmune response (Zhou et al. 2004). These data obtained from CDR3 spectratyping and clone sequencing are considerably limited, and features such as the biased usage of certain TRBVs in T cell repertoire, the highly expressed of certain CDR3 sequences, and base insertion in the CDR3 region need to be further characterized and analyzed with respect to the role they may play in the development of SLE. With the successful application of high-throughput sequencing in analyzing TCR CDR3 repertoire, (Klarenbeek et al. 2012; Meier et al. 2013; Wu et al. 2012) it is now possible to study the relationship between the complete TCR CDR3 repertoire and SLE. This study used Roche 454 high-throughput sequencing to analyze and compare the T cell (mRNA) TCR β-chain CDR3 repertoire from the thymuses and spleens of 1-, 3-, and 5-month-old MRL/lpr mice.
Materials and methods
Experimental animals and reagents
Female MRL/lpr (H-2K) mice of two different ages (4-weeks old with a body weight of 12–14 g and 10- to 11-weeks old with a body weight of 18–20 g) were purchased from Shanghai Laboratory Animal Center, Chinese Academy of Sciences. The mice were bred in the SPF experimental animal breeding center of Zunyi Medical University, and animals of different ages (1-month-, 3-month-, and 5-month-old mice) were randomly selected for thymus and spleen collection. TRIzol reagent was purchased from Invitrogen, agarose from Promega, the antinuclear antibody spectrum (IgG) kit from EUROIMMUN, the reverse transcription cDNA kit from Fermentas, agarose gel extraction and PCR product purification kits from Qiagen, and conventional PCR premixed solution (Premix Ex Taq) from Takara.
Primer design and synthesis
Six pairs of upstream and downstream primers for 22 mouse TRVB family genes were designed and synthesized according to previous reports (Matsutani et al. 2007), and the 5′ end of each primer pair was designed to contain a 10-base unique tag to distinguish the gene during high-throughput sequencing. Upstream and downstream primers for GAPDH were used as a control (Tables 1 and 2). All primers were synthesized by Shanghai Invitrogen Biotechnology Co., Ltd.
Preparation of TCR β-chain CDR3 repertoire from the thymus and spleen of a MRL/lpr mouse
Preparation of spleen and thymus tissue cell suspensions
After collecting peripheral blood from MRL/lpr mice by cardiac puncture, the mice were sacrificed and immersed in 75 % ethanol for 5 min. Thymus and spleen tissues were collected under sterile conditions and placed in a 3-mL phosphate buffer solution containing bovine serum albumin and ethylenediaminetetraacetate (PBE) buffer tube. PBE homogenate was collected using the gentle MACS dissociator, filtered twice through a 200-mesh sterile cell strainer, and then centrifuged at 1500 rpm for 8 min. The supernatant was discarded, and 5 mL of red blood cell lysis buffer was added. The cell suspension was filtered again and centrifuged at 1500 rpm for 8 min, and the supernatant was discarded to obtain thymus and spleen cells.
Total RNA extraction and cDNA synthesis
Thymus and spleen cell suspensions of MRL/lpr mice at different ages were centrifuged at 300×g for 5 min at 4 °C, and TRIzol was used to extract total RNA from each sample. For each sample, cDNA synthesis was carried out in four reaction tubes. In each tube, 15 μL total RNA was added to an RNeasy EP tube as template, and 2 μL oligo(dT) primer and 7 μL of DEPC H2O were added to the reaction tube and mixed well. Reactions were incubated at 65 °C for 5 min and then placed on ice. In each reaction, 2 μL of RiboLock™ Ribonuclease Inhibitor, 8 μL of 5× reaction buffer, 4 μL of 10 mM dNTP Mix, and 2 μL of RevertAid™ M-MuLV were added into each EP tube and mixed. Reactions were carried at 42 °C for 60 min, followed by 70 °C for 5 min. Samples were removed when the temperature dropped to 4 °C and were placed on ice. Samples were stored at −20 °C until use.
PCR amplification of the complete CDR3-encoding sequence of TRBV families and preparation of TCR β-chain CDR3 repertoire
For each sample, a pair of unique base-tagged primers was used to perform PCR amplification of the complete CDR3-encoding sequence for each of the 22 TRBV families. The total reaction volume (25 μL) contained 1 μL cDNA template, 2 μL of each upstream primer and the TRBV downstream primer with the unique base tag corresponding to one of the 22 TRBV families, 12.5 μL of Premix Ex Taq, and 7.5 μL of sterilized ultrapure water. The reaction conditions were as follows: 94 °C for 3 min, 94 °C for 1 min, 55 °C for 1 min, 72 °C for 2 min (35 cycles), and 72 °C for 10 min. Samples were held at 4 °C after the reaction and then stored at −20 °C.
Purification and detection of the CDR3 repertoire
CDR3 repertoire were recovered using a Gel Extraction Kit, and purification was performed using a PCR Purification Kit.
Detection of autoantibodies in the peripheral blood of MRL/lpr mice and kidney paraffin sections
Western blotting was used to detect antinuclear antibodies (of the IgG subclass) in each serum sample following the kit instructions. Kidney samples were fixed in 10 % formaldehyde for 24 h and then embedded in paraffin blocks. Pathological analysis was conducted after hematoxylin and eosin (HE) staining.
Roche 454 sequencing of CDR3 repertoire
The LabChip GX electrophoresis system was used to measure the concentration and total amount of each sample. The qualities of each sample sequence were consistent. The original sequence obtained from sequencing included a barcode sequence, primer sequence, base segments of adapter sequence, and the targeted CDR3. Sequences were screened using the Roche 454 sequencing standard; those with poor quality bases (R <20) or with an N content over 2 % were removed, and original sequences were distributed to each source sample using the unique tag and primer sequence of each sample.
IMGT/HighV-QUEST analysis and statistics
Barcode and primer sequences were removed from the original sequence of each sample, and each sample sequence was then submitted to IMGT/HighV QUEST analyzing system in FASTA format. We removed the sequences with ‘Warnings,’ ‘Unknown functionality,’ ‘No results,’ from the highest percent of identity of the V-REGION (more than 85 % compared with that of the closest gremlin V-REGION) functionality sequence and screened out total productive sequences to identify unique productive sequences and unique CDR3 amino acid sequences. Excel and other software programs were used to plot the data and perform statistical analysis (Alamyar et al. 2012; Li et al. 2013).
Results
Autoantibodies and glomerulus pathology in MRL/lpr mice
Three- and 5-month-old MRL/lpr mice showed obvious joint swelling, skin erythema, axillary lymph node enlargement, and spleen enlargement, which were not observed in 1-month-old mice. Three-month-old MRL/lpr mice were positive for anti-dsDNA antibodies (++), strongly positive for anti-ANA antibodies (+++), and positive for anti-AHA antibodies (++); 5-month-old MRL/lpr mice were strongly positive for anti-dsDNA antibodies (+++), strongly positive for anti-ANA antibodies (+++), and positive for anti-AHA antibodies (++); and 1-month-old MRL/lpr mice were negative (Supplementary Fig. 1 and Supplementary Table 1). HE staining of 3- and 5-month-old MRL/lpr mouse kidney sections showed proliferation of glomerulus mesangial cells with basement membrane thickening, glomerular sclerosis atrophy, major renal interstitial inflammatory cell infiltration, and inflammatory cell infiltration around blood vessels with fibrosis and onion-like lesions in some samples, which were not observed in the glomeruli of 1-month-old MRL/lpr mice (Supplementary Fig. 2).
TCR β-chain CDR3 repertoire preparation
TCR β-chain CDR3 repertoire from the thymuses and spleens from MRL/lpr mice at different ages showed clear bands corresponding to their sizes by agarose gel electrophoresis (the data for the thymus derived from a 1-month-old MRL/lpr is shown in Fig. 1. The data for the other samples are not shown). The cDNA concentration in each sample was greater than 20 ng/μL after recovery and purification, and the total amount in each sample was a minimum of 200 ng (Supplementary Fig. 3 and Supplementary Table 2).

The electrophoresis diagram of the PCR products (the CDR3 repertoire) from 22 TRBV gene families in 1-month-old mouse T cells from the thymus. In TRBV, a total of 22 families upstream and downstream primer amplification products by PCR, stripes were predicted in corresponding locations in large and small regions of CDR3 repertoire
Diversity in TCR β-chain CDR3 repertoire
Total original sequences (187,624 sequences) were obtained using Roche 454 high-throughput sequencing, and the number of total productive sequences obtained from the six samples was similar. Unique productive CDR3 sequences in thymus and spleen samples from 1-month-old MRL/lpr mice accounted for 93.979 and 74.472 %, respectively, of the total productive CDR3 sequences; 73.290 and 74.287 %, respectively, in 3-month-old mice; and 47.396 and 62.399 %, respectively, in 5-month-old mice, (Table 3). The diversity (unique CDR3 frequency) in the thymic TCR β-chain CDR3 repertoire from 3- and 5-month-old mice was significantly lower than that from 1-month-old mice, while the splenic repertoire from 5-month-old mice was less diverse than that from 1- and 3-month-old mice.
Biased usage of TRBV gene families in the TCR β-chain CDR3 repertoire
While examining the total productive sequences, a high bias toward TRBV13-3 gene family usage was observed in both the thymus and spleen of MRL/lpr mice at different ages; the bias toward TRBV usage was similarly high in both 3- and 5-month-old mice; both age groups showed a highly expressed TRBV26, TRBV17, and TRBV13-3 usage in the thymus and TRBV4, TRBV13-1, TRBV13-3, and TRBV26 usage in the spleen. The percentage of thymus TRBV13-3 biased usage in 1-, 3-, and 5-month-old MRL/lpr mice was 17.19, 9.39, and 14.73 %, respectively. The percentage of spleen TRBV4 biased usage in 1-, 3-, and 5-month-old MRL/lpr mice was 23.05, 18.92, and 6.95 %, respectively (Table 4).
The TRBV and TRBJ pairing in the thymus and spleen of MRL/lpr mice at different ages
The frequency of 22 TRBV gene pairing with the 12 TRBJ gene was identified (Fig. 6). Restricted usage of TRBJ2-5 in the thymus and TRBJ2-7 in the spleen, respectively, at 1-, 3-, and 5-month-old ages of MRL/lpr mice. But there were different usages of TRBV gene recombined with each TRBJ gene in the thymus and spleen of MRL/lpr mice at different ages. The overview TRBV pairing with TRBJ gene showed the reduced complexity of the TCR repertoires from 5-month- compared to 3-month- or 1-month-old MRL/lpr mice.
Clonal proliferation of TCR β-chain CDR3
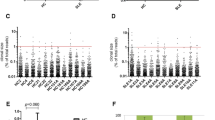
Clonal proliferation and the HEC numbers as determined by analysis of TCR β-chain CDR3 repertoire from MRL/lpr mice at different ages showed significant differences (Fig. 2). The number of highly expanded clones (HEC) in the thymuses of 3- and 5-month-old mice was higher than that in 1-month-old mice, while that in the spleens of 5-month-old mice was higher than the number in 1- and 3-month-old mice. Thymus samples from 1-month-old mice showed no significant clonal proliferation. HEC accounted for 1.1 % in 3-month-old mice and 2.8 % in 5-month-old mice; in spleen samples, HEC was 3.0 % in 1-month-old mice, 2.7 % in 3-month-old mice, and 6.3 % in 5-month-old mice.

T cell clone proliferation levels from the thymuses and spleens of MRL/lpr mice at different ages. a Scatter diagram to show the cloning status of the six samples from MRL/lpr mice. Each dot in the diagram represents one clone. The percentage in the T cell receptor repertoire could demonstrate the sizes of the T cell clones. b Clones from the six samples distributed with bias to the right. The average and standard deviation are shown in the diagram. c Thymus T cell and d spleen T cell clone frequency distribution
Sequence analysis of highly expressed clones (top 5) in TCR β-chain CDR3 repertoire
The incidence of highly expressed CDR3 sequences (top 5) from the thymus and spleen of MRL/lpr mice at different ages was unique. The thymus and spleen highly expressed CDR3 ratio was greater than 1 % in 5-month-old mice, whereas the ratios were below 1 % for both 1- and 3-month-old mice. Highly expressed CDR3s show partially conserved sequence motifs such as E-Q-F and Y-E-Q-Y (Table 5). The highly expressed CDR3 (CASSPGLGYNYAEQFF) in the spleen of a 5-month-old mouse came from the TRBV14 gene family, but TRBV14 gene were very low frequency expression at all other samples of MRL/lpr mice (Table 5, Fig. 6).
The average number of inserted bases in the N1 region of highly expressed CDR3 sequences from 3- and 5-month-old MRL/lpr mice spleens was significantly higher than that from 1-month-old mice (2.21, 2.49, and 0.48 for 5-, 3-, and 1-month-old mice, respectively); whereas in the thymus, 5-month-old mice (2.8) had a greater average number than 3- (1.52) and 1-month-old (1.68) MRL/lpr mice.
There were no significant differences between the average number of inserted bases in the N2 region of highly expressed CDR3 sequences from spleen or thymus samples from the different age groups; however, the average number of inserted bases in the N2 region was generally smaller than that in the N1 region (Fig. 3), and the average number of inserted bases in the N2 region of unique productive CDR3 sequences from the thymus and spleen of MRL/lpr mice at different ages was greater than that in the N1 region (Supplementary Fig. 4). The extent of base insertion in the N1 and N2 regions of unique productive thymic and splenic CDR3 sequences in MRL/lpr mice at different ages showed an uptrend with increased age. In addition, the average number of inserted bases in the N2 region was greater than that in the N1 region (average number of inserted bases in the N1 region of the thymus 2.13, 2.16, and 2.25; average number of inserted bases in the N1 region of the spleen 2.07, 2.28, and 2.30; average number of inserted bases in the N2 region of the thymus 2.72, 2.56, and 3.06; average number of inserted bases in the N2 region of the spleen 2.68, 2.74, and 2.90).

The T cells highly expressed proliferating clones (top five) TCR β-chain CDR3 region, N-region, and P-region insertion and splicing analysis of MRL/lpr mice at different ages
Statistical analysis of overlapping CDR3 of MRL/lpr mice at different ages
The CDR3-encoding sequences from the thymus and spleen of 1-month-old MRL/lpr mice have a low overlap ratio, and the overlap ratio have no differences between total productive and unique productive sequences that were observed. However, thymic and splenic CDR3 regions of 3- and 5-month-old MRL/lpr mice have high overlap ratios; furthermore, the total productive overlap ratio was significantly higher than the unique productive overlap ratio (Tables 6, 7 and Figs. 4, 5, and 6). There were 11 identical CDR3 sequences among thymuses and 8 identical CDR3 sequences among spleens of 1-, 3-, and 5-month-old mice (Tables 8 and 9). And there were no differences of the overlap CDR3 frequency in the 1-, 3-, and 5-month-old mice. The overlap CDR3 (GARDRANSDYT) in the thymus of 1-, 3-, and 5-month-old mice came from the same TRBV (TRBV20) family.

The unique productive and the total productive sequences of CDR3 region amino acid sequence overlapping ratios between the thymus and spleen of MRL/lpr mice at different ages

The unique productive sequences of CDR3 region amino acid sequence overlapping ratios of MRL/lpr mouse at different ages. a CDR3 region overlapping comparison among MRL/lpr-1-thymus (1-T)/MRL/lpr-3-thymus (3-T)/MRL/lpr-5-thymus (5-T) samples and MRL/lpr-1-spleen (1-S)/MRL/lpr-5-spleen (3-S)/MRL/lpr-5-spleen (5-S) samples. b CDR3 region overlapping comparisons among thymus and spleen samples from MRL/lpr mice at different ages

The TRBV and TRBJ pairing in the thymus and spleen of MRL/lpr mice at different ages. TRBJ genes family on the left and TRBV genes family at the bottom of the panels, the reads from the total productive CDR3 sequences for pairing were indicated by color code. Prominent TRBV14 gene family in the spleen of a 5-month old mouse, but TRBV14 gene were at very low frequency expression at all other samples from MRL/lpr mice. The overview TRBV pairing with TRBJ gene showed the reduced complexity of the TCR repertoires from 5-month compared to 3-month or 1-month MRL/lpr mice
Discussion
The MRL/lpr mouse is an inbred mouse model of SLE that shows robust T cell proliferation and activation at the onset of autoimmunity (Watanabe-Fukunaga et al. 1992). To date, the composition and variation of TCR β-chain CDR3 repertoire from the thymus and spleen of MRL/lpr mice have not been reported. MRL/lpr mice typically have a disease onset around 2 months of age, and approximately 50 % of mice die at around 5 months of age. In 3- and 5-month-old MRL/lpr mice in this study, joint swelling, skin erythema, axillary lymph node enlargement, and spleen enlargement were observed, and anti-dsDNA, anti-ANA, and anti-AHA antibodies were detected through autoantibody tests. In addition, proliferation of glomerular mesangial cells, basement membrane thickening, glomerular sclerosis atrophy, and major renal interstitial inflammatory cell infiltration were detected by pathological examination. Inflammatory cell infiltration was also observed around blood vessels with fibrosis and “onion skin” configuration in some mice. Those changes were less severe in 3-month-old MRL/lpr mice than in 5-month-old MRL/lpr mice, and no autoantibodies or renal pathological changes were observed in 1-month-old MRL/lpr mice. This indicates measurable autoimmune responses in 3-month-old MRL/lpr mice and obvious autoimmune responses and organ damage in 5-month-old MRL/lpr mice. We utilized Roche 454 high-throughput sequencing to compare and analyze the composition and variation in T cell (mRNA) TCR β-chain CDR3 repertoire from the thymus and spleen of MRL/lpr mice at 1, 3, and 5 months of age.
Roche 454 high-throughput sequencing of six samples resulted in 187,624 original total sequences, and the amounts of total data obtained from each sample were similar. The diversities of TCR β-chain CDR3 repertoire of thymuses and spleens from MRL/lpr mice significantly decreased with increased age, which may be due to increased self-response T cell ratio through clonal proliferation. By analyzing HECs of total productive CDR3 sequences from thymuses and spleens of the three different ages of MRL/lpr mice, we found that the HEC ratio in the thymus and spleen significantly increased with age. A previous study conducted by Simpson et al. suggested that epitope expansion occurs in early stages of SLE and increases with disease progression (Arbuckle et al. 2003). The progressive increase in the HEC ratio in the thymus and spleen may reflect an increase in autoreactive T cell clones induced by epitope expansion and chronic inflammation.
Among the total productive CDR3 sequences, an increased bias toward TRBV13-3 usage was observed in the thymuses (13.19, 9.39, and 14.73 %) and spleens (12.80, 13.49, and 11.06 %) of MRL/lpr mice at 1-, 3-, and 5-month-old MRL/lpr mice. High TRBV biased usage was similarly observed in both 3-month-old and 5-month-old mice. Prominent TRBV4 expression was observed in the spleens (23.05, 18.92, and 6.95 %) of 1-, 3-, and 5-month-old MRL/lpr mice, the decreased TRBV4 may be coming from the Treg clonal, consistent with TRBV13-3 and TRBV4 gene family usage in the spleen and other peripheral lymphoid organs of MRL/lpr mice, as reported by Casrough (Zhou et al. 2004). In a study using a transgenic mouse model of mixed connective tissue disease, Greidinger et al. also observed biased usage of TRBV13 and TRBV4 in spleen samples (Greidinger et al. 2008). These results suggest that TRBV families with bias usage are closely related to the MRL/lpr autoimmune response, which provides a basis to study specific T cell responses in these mice.
The recombined and restricted usage of specific TRBV and TRBJ genes maybe related to the diseases. And when we further analyzed the TRBV and TRBJ pairing in the thymus and spleen of MRL/lpr mice at different ages, we found the bias usage of TRBJ2-5 in the thymus and TRBJ2-7 in the spleen, respectively, in 1-, 3-, and 5-month-old ages of MRL/lpr mice, but the overview TRBV gene recombined with each TRBJ gene showed the reduced complexity of the TCR repertoires from 5-month compared to 3-month or 1-month MRL/lpr mice (Krell et al. 2013).
The observed occurrence of highly expressed (top 5) CDR3 sequences in the thymuses and spleens of MRL/lpr mice at different ages was inconsistent. Highly expressed CDR3 sequences in the thymus and spleen from a 5-month-old mouse account for over 1 % of the total sequences but accounts for less than 1 % in 3- and 1-month-old mice. No identical amino acid sequences were detected while analyzing highly expressed CDR3 between samples; however, we found that highly expressed CDR3 sequences contain partially conserved sequence motifs (E-Q-F and Y-E-Q-Y). And the highly expressed CDR3 (CASSPGLGYNYAEQFF) in the spleen of a 5-month-old mouse came from the TRBV14 gene family, but TRBV gene was at a very low frequency expression at all other samples in MRL/lpr mice. Moreover, Greidinger et al. also found the amino acid motifs E-Q-F and Y-E-Q-Y in spleens of patients with small nuclear ribonucleoprotein antigen-induced mixed connective tissue disease and an HLA-DR4 transgenic mouse model (Greidinger et al. 2008). These conserved amino acid sequences may be related to the anti-snRNP response. The average number of inserted bases in the N1 region of highly expressed CDR3 sequences from 5- and 3-month-old mouse spleens was significantly higher than that from 1-month-old mice (2.21, 2.49, and 0.48 for 5-, 3-, and 1-month-old mice, respectively). When analyzing N1 nucleotide insertion in the corresponding highly expressed CDR3 sequences in the thymus, the average number in 5-month-old mice (2.8) was greater than that in 3- (1.52) and 1-month-old (1.68) mice. There were no significant differences in the average number of inserted bases in the N2 region of highly expressed CDR3 sequences in 1-, 3-, and 5-month-old MRL/lpr mouse thymuses or spleens. However, when analyzing unique productive CDR3 sequences from thymuses and spleens of MRL/lpr mice at different ages, we found that the average number of inserted bases in the N2 region was greater than that in the N1 region, and both showed an upward trend with a positive correlation with age and with no significant differences, indicating that nucleotide insertion and excision in the N region of highly expressed CDR3 sequences in MRL/lpr mice may be closely related to the MRL/lpr autoreactive T cell response.
By statistical analysis of overlapped CDR3 of MRL/lpr mice at different ages, we found 11 identical CDR3 sequences among thymuses and eight identical CDR3 among spleens of 1-, 3-, and 5-month-old mice. And the overlap CDR3 (GARDRANSDYT) in the thymuses of 1-, 3-, and 5-month-old mice came from the same TRBV (TRBV20) family. Due to some of the overlapped sequences were coming from different TRBV families and most of these sequences (12/19) have N1 region or N2 region, they probably arise as a result of the antigen selected. These common clonal T cells might mediate autoimmune responses in these mice. Additionally, CDR3 regions between the thymus and spleen of a 1-month-old MRL/lpr mouse had a low overlap ratio, and no differences between total productive and unique productive sequences were observed. However, CDR3 regions between the thymus and spleen of 3- and 5-month-old MRL/lpr mice had high overlap ratios, and the total productive overlap ratio was significantly greater than the unique productive overlap ratio. Although other regions of these overlapping CDR3 sequences were found to differ based on further analysis, all sequences showed a highly biased usage of TRBV (including TRBV13 and TRBV2), and TRBJ (TRBJ2-7 and TRBJ1-1) families, suggesting that the overlapped sequences resulted from different T cell groups after high clonal proliferation and are positively correlated with the progression of autoimmune response in MRL/lpr mice.
Through the analysis and comparison of TCR β-chain CDR3 repertoire from the thymuses and spleens (mRNA) of MRL/lpr mice at different ages (1, 3, and 5 months), we found that diversity of TCR CDR3 repertoire of thymuses and spleens from MRL/lpr mice significantly decreased with age (disease progression). Features such as a common TRBV biased usage in the TCR CDR3 repertoire, significant changes in the N1 region of highly expressed CDR3 sequences with disease progression, composition of common overlapping CDR3 regions, and the frequency of the TRBV pairing with the TRBJ in the different-aged mice may be closely related to the MRL/lpr anti-self T cell response during onset and progression of disease. We did not sort the T cells into subsets (naïve T cells and effective T cells), these conclusions should to be proven by further experiments. Even so, this study provides a basis for studying autoreactive T cell response in SLE and a new perspective for exploring SLE with progression of disease in clonal level of TCR, which may provide a basis for studying the mechanism of the MRL/lpr autoreactive T cells response and tailor an individualized treatment targeting these T cells.
References
Alamyar E, Duroux P, Lefranc MP, Giudicelli V (2012) IMGT ((R)) tools for the nucleotide analysis of immunoglobulin (IG) and T cell receptor (TR) V-(D)-J repertoires, polymorphisms, and IG mutations: IMGT/V-QUEST and IMGT/HighV-QUEST for NGS. Methods Mol Biol 882:569–604
Andrews BS, Eisenberg RA, Theofilopoulos AN, Izui S, Wilson CB, McConahey PJ, Murphy ED, Roths JB, Dixon FJ (1978) Spontaneous murine lupus-like syndromes. Clinical and immunopathological manifestations in several strains. J Exp Med 148:1198–1215
Arbuckle MR, McClain MT, Rubertone MV, Scofield RH, Dennis GJ, James JA, Harley JB (2003) Development of autoantibodies before the clinical onset of systemic lupus erythematosus. N Engl J Med 349:1526–1533
Craft JE (2012) Follicular helper T cells in immunity and systemic autoimmunity. Nat Rev Rheumatol 8:337–347
Greidinger EL, Zang YJ, Jaimes K, Martinez L, Nassiri M, Hoffman RW (2008) CD4+ T cells target epitopes residing within the RNA-binding domain of the U1-70-kDa small nuclear ribonucleoprotein autoantigen and have restricted TCR diversity in an HLA-DR4-transgenic murine model of mixed connective tissue disease. J Immunol 180:8444–8454
Holbrook MR, Tighe PJ, Powell RJ (1996) Restrictions of T cell receptor beta chain repertoire in the peripheral blood of patients with systemic lupus erythematosus. Ann Rheum Dis 55:627–631
Klarenbeek PL, de Hair MJ, Doorenspleet ME, van Schaik BD, Esveldt RE, van de Sande MG, Cantaert T, Gerlag DM, Baeten D, van Kampen AH, Baas F, Tak PP, de Vries N (2012) Inflamed target tissue provides a specific niche for highly expanded T-cell clones in early human autoimmune disease. Ann Rheum Dis 71:1088–1093
Kolowos W, Herrmann M, Ponner BB, Voll R, Kern P, Frank C, Kalden JR (1997) Detection of restricted junctional diversity of peripheral T cells in SLE patients by spectratyping. Lupus 6:701–707
Krell PF, Reuther S, Fischer U, Keller T, Weber S, Gombert M, Schuster FR, Asang C, Stepensky P, Strahm B, Meisel R, Stoye J, Borkhardt A (2013) Next-generation-sequencing-spectratyping reveals public T-cell receptor repertoires in pediatric very severe aplastic anemia and identifies a beta chain CDR3 sequence associated with hepatitis-induced pathogenesis. Haematologica 98:1388–1396
Li S, Lefranc MP, Miles JJ, Alamyar E, Giudicelli V, Duroux P, Freeman JD, Corbin VD, Scheerlinck JP, Frohman MA, Cameron PU, Plebanski M, Loveland B, Burrows SR, Papenfuss AT, Gowans EJ (2013) IMGT/HighV QUEST paradigm for T cell receptor IMGT clonotype diversity and next generation repertoire immunoprofiling. Nat Commun 4:2333
Luo W, Ma L, Yao XS, Zou HY, Wen Q, Ruan GP, Wang XN (2006) Complementarity-determining region 3 analysis of T cell receptor beta chain variable region in peripheral blood mononuclear cells of patients with systemic lupus erythematosus. Nan Fang Yi Ke Da Xue Xue Bao 26:1128–1131
Matsutani T, Ohmori T, Ogata M, Soga H, Kasahara S, Yoshioka T, Suzuki R, Itoh T (2007) Comparison of CDR3 length among thymocyte subpopulations: impacts of MHC and BV segment on the CDR3 shortening. Mol Immunol 44:2378–2387
Meier J, Roberts C, Avent K, Hazlett A, Berrie J, Payne K, Hamm D, Desmarais C, Sanders C, Hogan KT, Archer KJ, Manjili MH, Toor AA (2013) Fractal organization of the human T cell repertoire in health and after stem cell transplantation. Biol Blood Marrow Transplant 19:366–377
Mok CC, Lau CS (2003) Pathogenesis of systemic lupus erythematosus. J Clin Pathol 56:481–490
Nakashima H, Akahoshi M, Masutani K (2006) Th1/Th2 balance of SLE patients with lupus nephritis. Rinsho Byori 54:706–713
Peng SL, Craft J (1996) T cells in murine lupus: propagation and regulation of disease. Mol Biol Rep 23:247–251
Scheinecker C, Bonelli M, Smolen JS (2010) Pathogenetic aspects of systemic lupus erythematosus with an emphasis on regulatory T cells. J Autoimmun 35:269–275
Shin MS, Lee N, Kang I (2011) Effector T-cell subsets in systemic lupus erythematosus: update focusing on Th17 cells. Curr Opin Rheumatol 23:444–448
Watanabe-Fukunaga R, Brannan CI, Copeland NG, Jenkins NA, Nagata S (1992) Lymphoproliferation disorder in mice explained by defects in Fas antigen that mediates apoptosis. Nature 356:314–317
Wu D, Sherwood A, Fromm JR, Winter SS, Dunsmore KP, Loh ML, Greisman HA, Sabath DE, Wood BL, Robins H (2012) High-throughput sequencing detects minimal residual disease in acute T lymphoblastic leukemia. Sci Transl Med 4:134ra63
Zhou G, Fujio K, Sadakata A, Okamoto A, Yu R, Yamamoto K (2004) Identification of systemically expanded activated T cell clones in MRL/lpr and NZB/W F1 lupus model mice. Clin Exp Immunol 136:448–455
Acknowledgments
We thank Beijing Genomics Institution (BGI) for help with TRBV CDR3 sequencing (with 454 GS FLX). The work was supported by grants from the National Prophase Project on Basic Research of China (973 pre-Program, 2008CB517310), the Program for New Century Excellent Talents in the University of China (NCET-10–0095), and the National Natural Science Foundation of China (31160195).
Author information
Authors and Affiliations
Corresponding author
Additional information
Ma Long—Co-first author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 1733 kb)
Rights and permissions
About this article

Cite this article
Li, Z., Long, M., ChunMei, L. et al. Composition and variation analysis of TCR β-chain CDR3 repertoire in the thymus and spleen of MRL/lpr mouse at different ages. Immunogenetics 67, 25–37 (2015). https://doi.org/10.1007/s00251-014-0809-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00251-014-0809-y