
Abstract
The environmental impact of major oil spills on marine microorganisms has yet to be thoroughly investigated using molecular biology techniques. The Deepwater Horizon (DWH) drilling rig explosion of 2010 affected an approximately 176,000 km2 surface area of the Gulf of Mexico (GOM) when an estimated 210 million gallons of oil from the Macondo Prospect spilled into the environment. Pelagic Sargassum, a complex of two surface drifting species (Sargassum natans and Sargassum fluitans) of marine brown macroalgae and a critically important habitat in the GOM ecosystem, was suffused by Macondo Prospect 252 oil released during the DWH event. Using 16S rRNA PCR and Roche 454 pyrosequencing, the effect of the oil on the bacterial population associated with pelagic Sargassum and contiguous waters was examined by comparing sequence data generated from samples collected from oiled and non-oiled locations in the northern GOM. Sequence data showed similar microbial composition in Sargassum regardless of exposure to oil primarily dominated by five phyla; Proteobacteria, Bacteroidetes, Actinobacteria, Verrucomicrobia, and unclassified bacteria. The microbial composition in water samples was significantly less diverse than for Sargassum and consisted primarily of Proteobacteria, Firmicutes, and Bacteroidetes. Due to the evenly distributed abundance of microbial species on oiled and non-oiled pelagic Sargassum, study findings indicate that DWH spilled oil had minimal effect on the composition and diversity of the microbial community associated with Sargassum and contiguous waters. However, higher abundances of Sulfitobacter and one species of Psychrobacter were found in oiled water samples when compared to non-oiled water samples indicating some effect of DHW oil in the microbial composition of seawater. Though there are a number of marine studies using molecular biology approaches, this is the first molecular examination of the impact of the DWH oil spill on bacterial communities associated with pelagic Sargassum and contiguous waters from the GOM.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Marine algae, or seaweed, refer to many different species of multicellular alga, including red, green, and brown algae. These algae are widely distributed organisms that have many different applications, including food, herbal supplements, drugs, and natural products [1–4]. Due to their prevalence in the world’s oceans, marine algae have a large influence on marine ecosystems as they are capable of carbon fixation through photosynthesis and are adept at shaping the structure of habitats [5]. Phaeophyceae, or brown algae, is a large class of marine algae comprised of about 2000 species distributed worldwide [6]. Pelagic Sargassum (Family Sargassaceae), a genus of brown algae and commonly known as “gulf-weed,” is distributed throughout tropical and subtropical waters and is a well-studied neustonic organism [4].
Pelagic Sargassum is a complex of two co-occurring species (Sargassum natans and Sargassum fluitans) of surface-drifting brown algae found exclusively in the Atlantic Ocean, including the Gulf of Mexico (GOM) [7]. These are the only holopelagic species among the greater than 200 species of Sargassum found worldwide. Though the two species slightly differ morphologically, both employ small gas-filled vesicles to maintain positive buoyancy within sunlit ocean surface waters (Fig. 1). Winds and ocean currents aggregate these surface drifters into configurations that range from widely dispersed clumps to large neustonic rafts tens of meters wide and windrows that extend across the ocean surface for tens of kilometers [8, 9] (Fig. 2). Pelagic Sargassum contributes to primary productivity in ocean surface waters, whereas associated epiphytic filamentous cyanobacteria contribute to overall primary production and nutrient recycling within the pelagic Sargassum complex [10, 11]. This association was previously reported by Carpenter [10] and further investigated and reviewed by Phlips and Zeman [11].

Pelagic Sargassum: Sargassum natans (left) and Sargassum fluitans (right). Photo credit: Hazel Oxenford

Pelagic Sargassum windrow in the northern Gulf of Mexico. Photo credit: James Franks
Pelagic Sargassum is the only naturally occurring biogenic habitat in the open ocean and serves as an essential habitat for a large and diverse assemblage of marine organisms, including numerous species of fish (Fig. 3) [12–16]. Although marine life associated with this open ocean drift community is a focus of ongoing research, little is known of the annual biomass, growth, distribution, and microbial communities associated with pelagic Sargassum in the GOM. Bacterial interactions with a host such as pelagic Sargassum have yet to be fully studied, particularly using molecular and genomics approaches. As with other seaweeds, pelagic Sargassum is expected to release large amounts of organic compounds that have the potential to increase bacterial colonization as shown by other studies that investigated the microbial composition on the surfaces of other algae [17–20]. Studies of interactions between algae and their associated microbial community provide insight into host-microbe interactions, established core microbial communities, organic matter consumption, microbial influence on algal growth and morphology, detection of algal pathogens, and global carbon cycling [17–22]. However, the microbial communities associated with pelagic Sargassum have not been described using the next generation sequencing. More importantly, as pelagic Sargassum is an important component of tropical and sub-tropical waters, it is unknown how this host-microbe interaction responds to disastrous polluting events such as chemical or oil spills.

Underwater photograph of fishes associated with pelagic Sargassum in the northern Gulf of Mexico. Photo credit: Ben Raines
The Deepwater Horizon (DWH) oil spill of 2010 resulted in an input of approximately 4.9 million barrels of crude oil from Macondo Prospect 252 (MP252) into the GOM [23, 24], affecting an estimated 176,000 km2 of GOM surface area [25]. As pelagic Sargassum is an ubiquitous surface feature throughout the northern GOM, and considering the vast surface spatial extent of the DWH spill footprint, it was evident that there was overlap between pelagic Sargassum and oil during the DWH event [12]. During an offshore ichthyoplankton survey cruise in the northern GOM in May 2010, the University of Southern Mississippi Gulf Coast Research Laboratory (USM-GCRL) scientists observed vast surface oil sheens interspersed with amorphous oil globules (∼0.3–12.0 cm dia.) and innumerable clumps of oil-infused pelagic Sargassum (Fig. 4a, b). These on-site observations (location, date, and time) were confirmed to be associated with the DWH spill via assessments of satellite imagery of ocean surface hydrography and oil spill surface movement in the northern GOM provided by Roffer’s Ocean Fishing Forecasting Service, Inc. (Fig. 5) [26].

a Oiled pelagic Sargassum and surface water observed boat side at collection station 1A in the Gulf of Mexico, 23 May 2010. b Oiled Sargassum sample from station 1A. Photo credit: James Franks

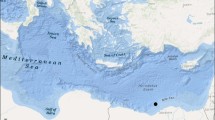
Collection locations of pelagic Sargassum and contiguous water samples analyzed in this study are shown on a map of the Gulf of Mexico (source: http://matplotlib.org/basemap) as white dots with Station ID number (see Table 1). Surface spatial extent of the Deepwater Horizon oil spill for 23–30 May 2010 (cumulative coverage) is shown as a gray overlay (provided by Roffer’s Ocean Fishing Forecasting Service, Inc.; http://www.roffs.com). The location of Station B01 was far removed from DWH surface oil and is shown on the map to provide perspective. Sea surface temperature data provided by the HYbrid Coordinate Ocean Model Consortium (https://hycom.org) are shown for 24 May 2010. The red circle indicates the location of the Deepwater Horizon well site (DWH)
To investigate the microbial composition of pelagic Sargassum and contiguous waters observed to be impacted by the DWH oil spill, samples of each, both visibly oiled and non-oiled, were collected for assessment. We hypothesize that the spilled MP252 oil had an effect on the bacterial composition and diversity of the communities associated with Sargassum and surrounding waters.
Materials and Methods
Sample Collection
Samples of non-oiled and oiled pelagic Sargassum (hereafter also referred to as Sargassum) and contiguous surface waters were collected from northern GOM locations (stations) in 2010 on 19 and 24 May and 23, 24, and 25 May, respectively (Table 1, Fig. 5). The Sargassum and water samples were collected concurrently at individual locations during daytime hours using sampling gear deployed from the USM-GCRL 30.5-m research vessel R/V Tommy Munro. Non-oiled Sargassum was collected using a surface neuston net (1 m × 2 m × 4 m, 330-micron mesh; Sea-Gear, http://www.sea-gear.net) towed by the research vessel. Oiled Sargassum was collected from surface waters using a dip net (1.5-cm-mesh stretched) and surface neuston net (described above). Clumps of Sargassum were quickly removed from the sampling gear by hand using nitrile gloves and placed into sterile, individually labeled 50-mL polypropylene centrifuge tubes. Water samples were also placed into sterile, individually labeled 50-mL polypropylene centrifuge tubes. All samples were immediately frozen (−20 °C) onboard the research vessel. A total of eight Sargassum samples and eight water samples were collected.
Global positioning system (GPS) coordinates were recorded for each collection, and hydrographic parameters, including sea surface temperature (°C), salinity (ppt), and dissolved oxygen (mg/L), were measured at each location using a calibrated YSI Model 85 multi-parameter instrument (Table 1; YSI Incorporated, www.ysi.com). Macrofauna collected in association with Sargassum were removed from the sample and returned to the sea. At the conclusion of the cruise, the frozen Sargassum and water samples were transferred on ice to USM-GCRL and maintained at −20 °C until analyzed. No specific permissions were required for this work.
DNA Extraction, 16S PCR, and Sequencing
The frozen Sargassum and water samples were express shipped to the J. Craig Venter Institute (JCVI) in Rockville, Maryland. Sargassum samples were thawed at 4 °C and then re-suspended in 50 mL TE buffer and vortexed, and the bacterial fraction was collected from the supernatant using a filtration kit provided by MO BIO Laboratories (Water Filter kit cat # 14880-50-WF), following manufacturer’s specifications. Water samples were filtered using the same method. Bacterial cells were lysed from each filter using lysozyme and proteinase digest, and DNA was extracted with phenol chloroform isoamyl alcohol and precipitated by ethanol precipitation. DNA was then quantified via NanoDrop (Thermo Scientific, http://www.nanodrop.com/) to determine concentration and overall quality using 260/280 ratio. Approximately 100 ng of DNA was used in each PCR reaction (5 min at 95 °C denaturing step followed by 30 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 2 min) using barcoded and Roche 454 adaptor ligated V1-3 16S primers 27F and 534R targeting the V1–3 variable regions of the 16S rRNA gene [27–30]. 16S rDNA gene amplicons were purified using the Qiagen PCR purification kit (Qiagen, www.qiagen.com) according to manufacturer’s specifications, followed by quantification using Sybr Gold, normalization, and pooling in preparation for Roche 454 sequencing. Emulsion PCR and Roche 454 pyrosequencing was completed with titanium chemistry using standard manufacturer’s protocols.
Quality Control and Processing of DNA Sequences
The DNA sequences were processed to ensure that only quality sequences were applied to the MOTHUR pipeline [31]. The SFF output files generated by the Roche 454 sequencer were first converted to fasta and qual files using sffinfo program included as part of the Roche 454 software package. The sequences were then demultiplexed according to barcode using the trim.seqs function in MOTHUR (version v.1.35.0) [31]. Stringent settings were kept to ensure no barcode mismatches among the demultiplexed reads. Additional sequence filtering was used by applying the screen.seqs function of mothur to remove all sequences shorter than 220 bp [31]. Additional QC steps were implemented and the sequences were aligned against the SILVA database to confirm the orientation of noise-filtered sequences along with ensuring the correct positioning of the reads with respect to which variable regions were amplified and sequenced [32]. The sequences passing QC were then checked for chimeras using chimera slayer in mothur [31]. Sequences were classified taxonomically using the RDP classifier (CITE) and hits matching mitochondria, chloroplast, archaea, unknown, and eukaryote were eliminated to avoid noise on the data. Sequence reads were then clustered at various taxonomic levels including 97 % rRNA sequence similarity (OTU), genus, and phyla.
Data Analysis
Species, genus and phyla count tables, and taxonomic classification of 16Sr RNA reads from the RDP Bayesian classifier (CITE) were used for all subsequent statistical analyses in the R statistical environment (CITE). The VEGAN and APE packages of R were used for all multivariate analyses, including Bray-Curtis dissimilarity, PCoA, PERMANOVA, ANOSIM, and sample clustering [31, 33, 34]. The mothur open-source software was also used to calculate Shannon, CHAO, and Jackknife diversity indices to calculate species richness and evenness. Kruskal-Wallis testing was used as a non-parametric approach to determine the statistical significance of the varying levels of abundance at the genus level between the samples. P values <0.05 were considered significant in these calculations.
Results
The extracted DNA from the majority of both Sargassum and water samples were amplified using standard 16S rRNA PCR conditions. However, one Sargassum sample (SARG5) and two water samples (WTR13 and WTR16) were unable to produce any amplicons after multiple rounds of PCR due to low-DNA yield. Post quality trimming and QC enabled analysis of 101,406 sequence reads with an average read length of 335 bp. Each sample averaged approximately 7800 sequence reads which were subsequently clustered at 97 % sequence identity to generate operational taxonomic units (OTUs).
Microbial Composition in Oiled and Non-Oiled Sargassum and Water Samples
Analysis of the 16S rRNA data demonstrated that the microbial population was overwhelmingly comprised of gram-negative species in pelagic Sargassum and water samples indicating that the oil had little effect on the overall gram-negative and gram-positive populations. Pelagic Sargassum collected at various distances from the DWH site was associated with a complex microbial community dominated mostly by various species of Rhodobacteraceae where the average among the Sargassum samples was 48.4 ± 17 %, followed by the second most abundant genera, Saprospiraceae 1.6 ± 0.81 %. Permutational analysis of variance and analysis of similarities indicated that there is a significant difference in microbial composition between Sargassum and water samples (P = 0.005 and P = 0.002 with R values, R = 0.903 and R = 0.9696, for PERMANOVA and ANOSIM, respectively). Sargassum and water, regardless of exposure to oil, were quite different in microbial diversity (Fig. 6a, b). The microbial composition in the water samples was less diverse compared to Sargassum, and consisted mostly of various species of Psychrobacter, Sulfitobacter, and Salegentibacter (Fig. 6b). These genera were significantly more abundant in water samples compared to non-oiled or oiled Sargassum (Kruskal-Wallis tests, P = 0.0011, 0.0044, and 0.0425, respectively). When comparing the two sample types and sample conditions, Sargassum samples, non-oiled or oiled, showed higher abundance of unclassified Rhodobacteraceae, unclassified Saprospiraceae, unclassified Sphingobacteriales, and Bacteroidetes, when compared to water samples (Kruskal-Wallis tests, P = values of 0.0042, 0.0016, 0.0275, and 0.0016, respectively). The oiled water samples appeared to be considerably different from the non-oiled water samples (Fig. 7) and were distinguishable by the abundance of two different species of Psychrobacter (Fig. 6b). Simpson diversity indices along with Shannon diversity indices were calculated to show the varying species richness and diversity between Sargassum and water (Fig. 8). Results indicate that the surface of pelagic Sargassum is contributing to the increased variation and number of species present in Sargassum samples compared to water samples.

Comparison of the microbial community in oiled and non-oiled water samples. The top 20 genera in Sargassum (a) and water (b)

Principal coordinates analysis (PCoA) of oiled and non-oiled Sargassum and oiled and non-oiled water samples. Each data point represents one sample, either Sargassum or water and oiled or non-oiled

Shannon diversity and Simpson diversity indices for Sargassum and water samples
Lack of Changes in Microbial Community Associated with Pelagic Sargassum
Overall, the microbial composition between oiled and non-oiled Sargassum samples did not change with exposure to DHW oil. Though our sample size was low, the microbial composition in the Sargassum samples did not seem to be affected by the presence of DWH oil as the most abundant taxa were present in nearly all samples, oiled or non-oiled. However, SARG3 had a higher abundance of Acidoverax, Aquabacterium, Ralstonia, Cloacibacterium, and Diaphorobacter when compared to the other oiled samples (Fig. 6a). The minimal effect of the oil on the Sargassum samples was also confirmed by principle coordinates analysis, as both the oiled Sargassum and non-oiled Sargassum samples clustered similarly with the exception of one oiled sample SARG3 (Fig. 7). These results indicate that although SARG7 was collected from an unaffected area several hundred miles away from the oil spill and SARG3 was well within the areas affected by DHW oil (Fig. 5), the microbial community did not significantly vary between the oiled and non oiled sample types.
DWH Oil Effect on Microbial Composition in Water
The effect of DWH oil on the water samples was much more apparent than on the Sargassum samples. The bacterial community in the water samples was dominated by various species of Psychrobacter averaging approximately 75.4 ± 16.4 % for each sample. More importantly, the oiled water samples were considerably different from the non-oiled samples (P = 0.0014, R 2 = 0.816) due to variation between two species of Psychrobacter (Fig. 6b). Due to the limitations of taxonomic identification using 16S sequencing, we were only able to differentiate between the two genera without identifying actual species names. The oiled water samples had significantly higher levels of what we designated as Psychrobacter 2 whereas the non-oiled water samples were dominated with what we designated as Psychrobacter 1. Principle coordinates analysis also showed that the oiled water samples clustered much differently from the non-oiled water samples where the variation was apparent along PCoA2 (Fig. 7). Three out of the four oiled water samples also showed a higher abundance of one species of Sulfitobacter when compared to non-oiled water samples (Fig. 6b). These striking differences clearly show that the DHW oil had an effect on the microbial population of the water.
Dissolved Oxygen Readings and Presence of Oil Metabolizing Bacteria in Oiled Sargassum and Water
Microbial community composition did not correlate with the amount of dissolved oxygen in the water. There was a lack of any specific trends with the number of species present in each sample and the exposure to MP252 oil in Sargassum and water samples. The oxygen concentration also did not show any specific trends based on sample type and MP252 oil exposure (Table 1). In addition to these readings, OTUs representing known oil-metabolizing species such as Acinetobacter, Thalassospira, and Pseudomonas, along with various genera belonging to the Oceanospirillales order [35, 36] were found in the oiled Sargassum and oiled water samples. Other known oil-degrading bacteria such as Pseudoalteromonas and Halomonas, along with unclassified genera belonging to the Halomonadaceae family [36], were detected in both oiled and non-oiled Sargassum and water samples showing no statistical difference in abundance between the sample types for these genera.
Discussion
Sargassum supports a dynamic GOM pelagic ecosystem as essential habitat and substrate for a multitude of motile and sessile organisms, including species of significant ecological and economic importance [6, 8]. The complexities of the pelagic Sargassum community and its multi-faceted contributions to ocean dynamics are poorly understood and require detailed investigation. The biomass of pelagic Sargassum impacted by MP252 oil is unknown; however, remotely sensed imagery of the areal surface extent of spilled oil included regions of the northern GOM where pelagic Sargassum habitat occurs [26, 37]. MP252 oil came into contact with numerous GOM Sargassum rafts, and aerial surveys conducted post- and pre-DWH spill provided evidence of both loss and subsequent recovery of Sargassum [6]. Although impacts to the microbial associates of Sargassum during the spill event are yet not well known, mesocosm experiments demonstrate potential pathways of MP252 oil injury to pelagic Sargassum and microbial associates [6].
Our results showed that there were clear differences between the two sample types through principal coordinates analysis and various statistical approaches. Though it was unusual to see one non-oiled Sargassum sample, SARG7, cluster with the other oiled Sargassum samples, we can only speculate the reasoning behind this result from what is known from previous studies in regards to other brown algae and the associated microbial population since our sample size is limited. Previous studies have shown that brown algae have associated bacteria that are responsible for growth and algal morphology, in addition to a “core” microbial population [17, 19, 21]. This core population in regards to our Sargassum study may be providing a protective effect from environmental contaminants and toxins such as MP252 oil. Currently, we can only speculate on this matter since our data shows that after contact with MP252 oil the microbial composition of oiled and non-oiled Sargassum, samples are relatively similar (Fig. 6a) in addition to the Sargassum samples clustering together regardless of exposure to MP252 oil (Fig. 7). Additional results also showed statistical significance in the abundance of several species between the two sample types as Sargassum hosts a more complex community of higher organisms. Pelagic Sargassum was observed to have a higher abundance of various species of unclassified Rhodobacteraceae, in addition to other common marine microbes belonging to the Alphaproteobacteria and Betaproteobacteria clades, when compared to the water samples. These results are somewhat anticipated as Rhodobacteraceae are metabolically versatile marine organisms and are capable of an extensive range of metabolic processes from nitrogen fixation and photosynthesis to oil degradation [35, 38]. It is also likely that the lack of changes in the microbiota associated with pelagic Sargassum was due to this high abundance of Rhodobacteraceae possibly providing a “protective” effect, as these families of bacteria are known hydrocarbon degraders. The significantly higher abundance of Saprospiraceae and Sphingobacteriales in Sargassum is likely due to the role that these two clades play in the breakdown of complex organic material in the environment [39, 40]. For instance, Saprospiraceae have been demonstrated to hydrolyze and utilize complex carbon sources such as sludge water, and Sphingobacteriales are often present in environmental samples that have come into contact with human waste matter [39, 40]. These previous findings are significant in that these two genera are absent in the water samples, yet present in pelagic Sargassum, an environment that serves as a nursery for juvenile fishes, where fish excrement is likely to be copious and has previously been demonstrated to influence the growth of Sargassum [41]. Though human and fish excrement vary in composition, the additional hydrocarbons and ammonia likely have a similar causative effect on the increased abundance of Saprospiraceae and Sphingobacteriales. Oiled and non-oiled water samples, on the other hand, were observed to have higher abundance of common marine microbes that are capable of surviving in harsh conditions with limited nutrients such as Psychrobacter, Salinimicrobium, and Salegentibacter, when compared to the Sargassum samples [42, 43].
Our sequence data also showed an unusually large proportion of Psychrobacter in all water samples regardless of exposure (or non-exposure) to oil. However, one species of Psychrobacter (designated as Psychrobacter 1) was present in larger abundance in non-oiled water samples when compared to another species of Psychrobacter (designated as Psychrobacter 2) in oiled samples (Fig. 6b). It would be interesting to see which species of Psychrobacter is more abundant in one sample type than the other; however, as our study only consisted of 16S rRNA sequencing, this would only be attained using metagenomics approaches or targeted PCR. As Psychrobacter are known oxidase positive, non-motile, and psychrotolerant organisms, it was surprising to see one species of Psychrobacter in higher abundance in oiled samples and a different Psychrobacter species in non-oiled samples, as this genus is known to be capable of surviving extreme conditions [44]. Since little is known on the effect of pollutants such as oil on the metabolic processes of various species of Psychrobacter, it would be interesting to see which species was able to tolerate the MP252 oil. Currently, we can only speculate on the striking differences between the two species of Psychrobacter without metagenomic and transcriptomic data.
Interestingly, it was also determined that the abundance of Sulfitobacter was much higher in oiled water samples, with the exception of WTR9. This is a noteworthy, yet expected, finding as this species of bacteria is capable of degrading crude oil in seawater [38]. However, Hazen et al. and Valentine et al. did not report Sulfitobacter from samples collected in the deep (ca. 1100–1200 m), dispersed GOM oil plumes, as this organism is known to be a photoheterotroph and would be less likely to survive deep waters where sunlight is virtually nonexistent [8, 45]. However, as these studies were completed using Phylochip microarray, it is likely that the molecular approaches in our study are able to detect other species of Sulfitobacter that were not detected using previous methods. The presence of other known obligate oil-degrading bacteria (OODB): Acinetobacter, Thalassospira, and Pseudomonas, along with various unclassified genera belonging to the Oceanospirillales order, on oiled Sargassum and in oiled water samples, was also quite significant as these organisms were not present in the non-oiled Sargassum and water samples. Though this is an important find in regards to Sargassum exposed to MP252 oil, it was also rather expected as these organisms have been either isolated from seawater exposed to oil or have been demonstrated to be associated with MP252 oil from the DHW oil spill [36, 46–48]. It was also noteworthy to see a few known OODB; Pseudoalteromonas, Halomonas, and unclassified Halomonadaceae present in both oiled and non-oiled Sargassum and water samples as previous studies have shown an increase in growth of Pseudoalteromonas and Halomonas when exposed to various types of hydrocarbons [47, 49]. Although 16S rRNA sequencing only provided us with a superficial interpretation of the effect of the DHW oil spill on the Sargassum associated bacterial communities, these results did provide us with some insight on which species were present in what is likely a response to the DHW spill as specific OTUs belonging to OODB were not present in the non-oiled samples.
Our findings provide substantial insight into the microbial community associated with pelagic Sargassum; a prominent marine surface community, which has not been previously studied using molecular approaches. Though our results were based on a limited number of samples, a sample set such as this one (pre- and post-oil spill water and Sargassum samples) is rarely obtained. Additionally, our results provide novel insight concerning the effect of MP252 oil on the microbial communities associated with Sargassum in the GOM. Additional genomics approaches such as metagenomics and transcriptomics to characterize functional elements in the microbial community associated with Sargassum can provide useful information in determining which key species and bacterial strains, along with which genes are responsible for maintaining a balanced ecosystem. In this regard, these techniques could reveal whether the lack of changes in community composition in the Sargassum really mean no changes at the metabolic level (community redundancy as opposed to functional redundancy). Further genomic-level approaches would also shed light on to whether the Sargassum microecosystem is naturally adapted to hydrocarbon-associated disturbances and can also help in providing a complete picture of how marine microbial communities recover after an event such as the DHW oil spill. In addition to determining overall Sargassum and associated macro-fauna health, it would be interesting to characterize the microbial community associated with Sargassum during massive algal blooms, which have been of great interest, occurring in areas of the tropical Atlantic Ocean not commonly associated with this marine alga. Exploratory approaches using ‘omics and next generation sequencing methods on pelagic Sargassum microbial communities are critical to understanding the role that these complex communities have in the health of the Sargassum and associated living marine organisms, as well as their recovery from devastating pollution events such as oil spills.
References
Blunt JW, Copp BR, Keyzers RA, Munro MH, Prinsep MR (2016) Marine natural products. Nat Prod Rep 33(3):382–431. doi:10.1039/c5np00156k
de Jesus Raposo MF, de Morais AM, and de Morais RM (2016) Emergent sources of prebiotics: seaweeds and microalgae. Mar Drugs 14(2). pii: E27. doi:10.3390/md14020027.
de Jesus Raposo MF, de Morais AM, de Morais RM (2015) Marine polysaccharides from algae with potential biomedical applications. Mar Drugs 13(5):2967–3028. doi:10.3390/md13052967
Yende SR, Harle UN, Chaugule BB (2014) Therapeutic potential and health benefits of Sargassum species. Pharmacognosy Rev 8(15):1–7
Schiel DR, Foster MS (2006) The population biology of large brown seaweeds: ecological consequences of multiphase life histories in dynamic coastal environments. Annu Rev Ecol Evol Syst 37(1):343–372
van den Hoek C, Mann DG, and Jahns HM (1996) Algae: an introduction to phycology. Cambridge University Press
South Atlantic Fishery Management Council (2002) Fishery management plan for pelagic Sargassum habitat of the South Atlantic region. p. 1-153.
Hazen TC et al (2010) Deep-sea oil plume enriches indigenous oil-degrading bacteria. Science 330(6001):204–208
Carr A (1987) New perspectives on the pelagic stage of sea turtle development. Conserv Biol 1(2):103–121
Carpenter EJ, Cox JL (1974) Production of pelagic Sargassum and a blue-green epiphyte in the western Sargasso Sea. Limnol Oceanogr 19(3):429–436
Phlips EJ, Zeman C (1990) Photosynthesis, growth and nitrogen fixation by epiphytic forms of filamentous cyanobacteria from pelagic Sargassum. Bull Mar Sci 47(3):613–621
Powers SP et al (2013) Novel pathways for injury from offshore oil spills: direct, sublethal and indirect effects of the Deepwater Horizon oil spill on pelagic Sargassum communities. PLoS One 8(9):e74802
Dooley JK (1972) Fishes associated with the pelagic Sargassum complex, with a discussion of the Sargassum community. Contrib Mar Sci 16:1–32
Wells RJD, Rooker JR (2004) Spatial and temporal patterns of habitat use by fishes associated with < i > Sargassum</i > mats in the northwestern Gulf of Mexico. Bull Mar Sci 74(1):81–99
Hoffmayer ER et al (2005) Larval and juvenile fishes associated with pelagic Sargassum in the northcentral Gulf of Mexico. Proc Gulf Caribb Fisheries Inst 56:259–270
Casazza TL, Ross SW (2008) Fishes associated with pelagic Sargassum and open water lacking Sargassum in the Gulf Stream off North Carolina. Fish Bull 106(4):348–363
Singh RP, Reddy CR (2014) Seaweed-microbial interactions: key functions of seaweed-associated bacteria. FEMS Microbiol Ecol 88(2):213–30
Martin M et al (2015) The cultivable surface microbiota of the brown alga Ascophyllum nodosum is enriched in macroalgal-polysaccharide-degrading bacteria. Front Microbiol 6:1487
Hester ER et al. (2016) Stable and sporadic symbiotic communities of coral and algal holobionts. ISME J 10: 1157–1169
Zozaya-Valdes E, Egan S, Thomas T (2015) A comprehensive analysis of the microbial communities of healthy and diseased marine macroalgae and the detection of known and potential bacterial pathogens. Front Microbiol 6:146. doi:10.3389/fmicb.2015.00146
Hollants J et al (2013) What we can learn from sushi: a review on seaweed-bacterial associations. FEMS Microbiol Ecol 83(1):1–16
Egan S et al (2013) The seaweed holobiont: understanding seaweed-bacteria interactions. FEMS Microbiol Rev 37(3):462–76
National Oceanic Atmospheric Administration (2015) About the Deepwater Horizon oil spill. Available from: http://www.gulfspillrestoration.noaa.gov/oil-spill/.
Zunkunft PF (2010) Summary report for sub-sea and sub-surface oil and disperion detection: sampling and monitoring. Available from: http://www.restorethegulf.gov/sites/default/files/documents/pdf/OSAT_Report_FINAL_17DEC.pdf.
Norse EA, Amos J (2010) Impacts, perception, and policy implications of the BP/Deepwater Horizon oil and gas disaster. Environ Law Report 40(11):11058–11073
Roffer MA (2015) Roffer’s Ocean Fishing Forecasting Service Inc. [cited 2015 21 September]; Available from: http://www.roffs.com/
Edwards U et al (1989) Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res 17(19):7843–53
Muyzer G, de Waal EC, Uitterlinden AG (1993) Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59(3):695–700
Liu Z et al (2008) Accurate taxonomy assignments from 16S rRNA sequences produced by highly parallel pyrosequencers. Nucleic Acids Res 36(18):e120
Kumar PS et al (2011) Target region selection is a critical determinant of community fingerprints generated by 16S pyrosequencing. PLoS One 6(6):e20956
Schloss PD et al (2009) Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol 75(23):7537–41
Pruesse E et al (2007) SILVA: a comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res 35(21):7188–7196
Paradis E, Claude J, Strimmer K (2004) APE: analyses of phylogenetics and evolution in R language. Bioinformatics 20(2):289–290
Jari Oksanen FGB, Roeland K, Pierre L, Peter RM, O’Hara RB, Simpson GL, Solymos P, Stevens MHH and Wagner H (2015) vegan: community ecology package. R package version 2.3-0. [cited 2016; R package version 2.3-0]. Available from: http://CRAN.R-project.org/package=vegan
Kostka JE et al (2011) Hydrocarbon-degrading bacteria and the bacterial community response in Gulf of Mexico beach sands impacted by the Deepwater Horizon oil spill. Appl Environ Microbiol 77(22):7962–74
Yakimov MM, Timmis KN, Golyshin PN (2007) Obligate oil-degrading marine bacteria. Curr Opin Biotechnol 18(3):257–66
Administration, N.O.A. (2015) National Environmental Satellite, Data, and Information Service. [cited 2015 21 September]; Available from: http://www.nesdis.noaa.gov/index.html
Brakstad OG, Lodeng AG (2005) Microbial diversity during biodegradation of crude oil in seawater from the North Sea. Microb Ecol 49(1):94–103
Buchan A et al (2014) Master recyclers: features and functions of bacteria associated with phytoplankton blooms. Nat Rev Micro 12(10):686–698
McIlroy S. and Nielsen P 2014 The Family Saprospiraceae, in The Prokaryotes, Rosenberg E et al, Editors. Springer Berlin Heidelberg. p. 863-889
Lapointe BE et al (2014) Ryther revisited: nutrient excretions by fishes enhance productivity of pelagic Sargassum in the western North Atlantic Ocean. J Exp Mar Biol Ecol 458:46–56
Lee SY et al (2012) Salinimicrobium gaetbulicola sp. nov., isolated from tidal flat sediment. Int J Syst Evol Microbiol 62(Pt 5):1027–31
Nedashkovskaya OI et al (2010) Salinimicrobium marinum sp. nov., a halophilic bacterium of the family Flavobacteriaceae, and emended descriptions of the genus Salinimicrobium and Salinimicrobium catena. Int J Syst Evol Microbiol 60(Pt 10):2303–6
Bozal N et al (2003) Characterization of several Psychrobacter strains isolated from Antarctic environments and description of Psychrobacter luti sp. nov. and Psychrobacter fozii sp. nov. Int J Syst Evol Microbiol 53(Pt 4):1093–100
Valentine DL et al (2010) Propane respiration jump-starts microbial response to a deep oil spill. Science 330(6001):208–211
Kodama Y et al (2008) Thalassospira tepidiphila sp. nov., a polycyclic aromatic hydrocarbon-degrading bacterium isolated from seawater. Int J Syst Evol Microbiol 58(Pt 3):711–5
Brakstad OG et al (2015) Microbial communities related to biodegradation of dispersed Macondo oil at low seawater temperature with Norwegian coastal seawater. Microb Biotechnol 8(6):989–98
Brakstad OG, Nordtug T, Throne-Holst M (2015) Biodegradation of dispersed Macondo oil in seawater at low temperature and different oil droplet sizes. Mar Pollut Bull 93(1-2):144–52
Chronopoulou PM et al (2015) Generalist hydrocarbon-degrading bacterial communities in the oil-polluted water column of the North Sea. Microb Biotechnol 8(3):434–47
Acknowledgments
The authors extend their sincere appreciation to the Guy Harvey Ocean Foundation for supporting critical aspects of this research. We acknowledge NOAA Fisheries, Southeast Fisheries Science Center, and SEAMAP for providing funding in support of the ichthyoplankton survey cruise. This study was partially supported by the Gulf of Mexico Research Initiative administered by the Northern Gulf Institute, Mississippi State University, and by the J. Craig Venter Institute. We express our gratitude to the captains and the crew of the R/V Tommy Munro and to Marcia Pendleton for the assistance in sample collection. We thank Tracy Berutti for performing the DNA extractions. We express our appreciation to Mitch Roffer, Greg Gawlikowski, and Matthew Upton (Roffer’s Ocean Fishing Forecasting Service, Inc.; http://www.roffs.com) for providing satellite imagery/analysis and invaluable advice to assist us in the selection of collection stations. Jason Tilly assisted in developing Fig. 3. We thank Ben Raines for the use of his underwater photograph of fishes associated with pelagic Sargassum.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no conflict of interest.
Rights and permissions
About this article

Cite this article
Torralba, M.G., Franks, J.S., Gomez, A. et al. Effect of Macondo Prospect 252 Oil on Microbiota Associated with Pelagic Sargassum in the Northern Gulf of Mexico. Microb Ecol 73, 91–100 (2017). https://doi.org/10.1007/s00248-016-0857-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-016-0857-y