
Abstract
Diverse and abundant bacterial populations play important functional roles in the multi-partite association of the coral holobiont. The specificity of coral-associated assemblages remains unclear, and little is known about the inheritance of specific bacteria from the parent colony to their offspring. This study investigated if broadcast spawning and brooding corals release specific and potentially beneficial bacteria with their offspring to secure maintenance across generations. Two coral species, Acropora tenuis and Pocillopora damicornis, were maintained in 0.2 μm filtered seawater during the release of their gametes and planulae, respectively. Water samples, excluding gametes and planulae, were subsequently collected, and bacterial diversity was assessed through a pyrosequencing approach amplifying a 470-bp region of the 16S rRNA gene including the variable regions 1–3. Compared to the high bacterial diversity harboured by corals, only a few taxa of bacteria were released by adult corals. Both A. tenuis and P. damicornis released similar bacteria, and the genera Alteromonas and Roseobacter were abundant in large proportions in the seawater of both species after reproduction. This study suggests that adult corals may release bacteria with their offspring to benefit the fitness in early coral life stages.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Corals live in close and complex relationships with their associated microbial partners including dinoflagellates (Symbiodinium spp.), bacteria, archaea, fungi and viruses. This assemblage of organisms, together known as the coral holobiont [11], inhabits a range of microhabitats within the coral, such as the carbonate skeleton, the internal tissue and the surface mucus layer [22]. Their beneficial contributions to the coral host, including nitrogen fixation [13, 14], defence [21] and coral settlement [27] have been widely reported.
The mechanism controlling coral–bacterial interactions is poorly understood, and it is currently unknown whether true symbiosis or long-term species specificity between corals and bacteria exists. Corals exhibit a range of reproductive strategies, including sexual and asexual propagation. These reproductive modes were represented in the current study by investigating two corals: Acropora tenuis and Pocillopora damicornis. A. tenuis reproduces broadcast spawning gametes for external fertilisation during annual mass spawning events which occur worldwide, usually once a year in summer, and involve the synchronous release of gametes from benthic invertebrates, including corals. In contrast, the brooding coral P. damicornis undergoes internal fertilisation and breeding and expels developed larvae at various times of the year, usually over the summer months.
The acquisition of symbionts between generations is referred to as either horizontal transmission, where the host acquires the symbionts from the surrounding environment, or as vertical transmission with microbial partners being passed on to the next generation via the egg. Various marine invertebrates, including ascidians [10], bivalves [12] and sponges [25], have been documented to transmit specific bacteria vertically to the next generation. The well-studied association between the bobtail squid Euprymna scolopes and the luminous bacterium Vibrio fischeri is regarded a model for the specific establishment, development and maintenance of horizontally transmitted symbiosis [15]. The establishment of this partnership has shown high levels of specificity and involves a step-wise elimination of potential interlopers, ensuring the exclusive symbiosis with V. fischeri, as opposed to thousands of other potential microbial partners [17]; not only does the host acquire specific species, but a specific strain of bacteria.
Previous studies revealed that the majority of broadcast spawning corals, as opposed to brooding corals, do not transfer their algal symbionts Symbiodinium with their gametes (vertical transfer) but acquire these after settlement and metamorphosis into the juvenile polyp (horizontal transfer) [20]. Similar observations have been reported for coral–bacterial partners, with several broadcast spawning corals shown to only have bacterial associated communities after settlement [1, 26]. However, the brooding coral Porites astreoides transmits bacteria vertically to planulae with two bacterial taxa (Roseobacter clade-associated bacteria and Marinobacter spp.) consistently and stably associated with juvenile P. astreoides [24].
The intergenerational maintenance of selected coral partners suggests highly evolved associations in which the presence of bacteria is advantageous and might fulfil important roles in the fitness and survival of the coral host. The present study explored whether corals initiate associations with bacteria via inheritance from parent colonies by releasing beneficial bacteria with their gametes and planulae, a previously unexplored and potentially alternative strategy to horizontal and vertical transmission.
Materials and Methods
Sample Site and Sample Collection
All corals were sampled on a reef flat (5–6-m water depth) near Coral Bay (23 °07′ S, 113°07′ E), Ningaloo Reef, Western Australia. Two coral colonies from each coral species, A. tenuis and P. damicornis colonies, were removed from the reef structure and kept in open plastic containers (80 × 50 × 50 cm) on the reef flat during the day and regularly assessed for reproductive activity on the beach at night time. The containers were kept in knee-deep water to maintain the ambient water temperature and returned to the reef at sunrise. On the night of coral spawning, A. tenuis colonies were rinsed with and transferred into 0.2 μm filtered seawater, and one water sample of 1 L was collected from the container immediately after coral spawning. The same procedure was conducted for planulating P. damicornis corals 1 week later. Water samples were filtered through Sterivex (0.22 μm) filter columns (Millipore, Fisher Scientific, UK Ltd.) and were stored at −80 °C until required for analysis. Samples were preserved for DNA extraction within an hour of collection.
DNA Extraction, PCR and Sequencing of Water Samples
DNA was extracted from Sterivex filters by the addition of 0.2 mL lysozyme (10 mg mL−1), incubation at 37 °C for 45 min, addition of 0.2 mL of proteinase K (0.2 mg mL−1) and 1 % SDS and incubation at 55 °C for 1 h. Lysates were recovered into fresh Eppendorf tubes, and DNA was extracted with a standard phenol/chloroform/isoamyl alcohol procedure and precipitated with 0.8 vol. of isopropanol [23]. A 470-bp region of the 16S ribosomal RNA gene (16S rRNA), including the variable regions 1–3, was selected for tag pyrosequencing, following the methods outlined in Ceh et al. 2012 [4].
Sequence and Statistical Analyses
The sequence Fasta and quality files were extracted from the raw sff output from the 454 sequence runs, and the sequence tag and its associated quality scores were removed. The python script split_libraries.py from the quantitative insights into microbial ecology (QIIME) pipeline [3] was used to remove poor quality (<25) and short sequences (<150 bp), remove the primer and barcode, and add a sample identifier to the header of each sequence. The resulting Fasta file was checked for chimeric sequences against a chimera-free database of 16S rRNA gene sequences (Green Genes, 29 November 2010 release) using UCHIME [8]. All sequences that were identified as potential chimeras were removed. Homopolymer sequence errors were corrected using the programme ACACIA [2], resulting in a chimera and error-free Fasta file. The number of reads per sample was quantified for each of the previous steps. The number of chimera-free and error-free reads was normalised to 1,130 reads per sample to allow comparative diversity analysis between all samples. No significant differences (P = 0.01; 1,000 permutations) were observed between the raw, cleaned and normalised datasets when PCA analysis was performed on the relative abundance of the dominant operational taxonomic units (OTUs), and correlation between datasets was assessed by Procrustes rotation [9]. Therefore, all analysis reported in this study was conducted on the randomly subsampled and normalised dataset. Sequences were clustered using UCLUST [7] to obtain groups of sequences at both the 90 and 97 % similarity levels. These groups represent operational taxonomic units defined at an approximate ‘genus’ and ‘species’ levels. The QIIME pipeline was used to identify the most abundant member of each group which was subsequently chosen as the representative sequence. Sequence taxonomy was assigned using GreenGenes [6] and BLAST (0.75 similarity, and the QIIME pipeline was used to generate OTU tables. Alpha diversity statistics in QIIME were calculated after random subsampling to ensure that sequencing effort did not affect diversity comparisons. The pyrosequencing dataset were deposited in the NCBI Sequence Read Archive database with the accession number (SRA053023.1).
Results
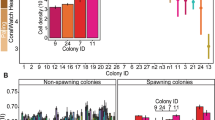
Water samples from spawning A. tenuis and planulating P. damicornis provided a total of 3,465 high-quality 16S rRNA gene sequence tags (Table 1). Grouping sequences at 97 % level of identity resulted in ten OTUs from three bacteria classes (Flavobacteria, α-Proteobacteria, γ-Proteobacteria), occurring in proportions larger than 1 % for either sample (Fig. 1). Water derived from A. tenuis displayed six OTUs with large proportions in the α-Proteobacteria (90.4 %) and smaller proportions in the Flavobacteria (4.5 %) and γ-Proteobacteria (1.8 %). Within the α-Proteobacteria, sequences affiliated with Roseobacter sp. were dominant, representing 89.7 % of all retrieved sequences. Other OTUs were affiliated with the α-Proteobacteria Rhodomicrobium (0.6 %) and Rhodospirillales (0.1 %) and the γ-Proteobacteria Shewanella (0.6 %) and Alteromonas (1.2 %). Water derived from P. damicornis was represented by ten OTUs dominated by sequences affiliated with the γ-Proteobacteria (82.7 %), α-Proteobacteria (10.1 %) and Flavobacteria (0.3 %). Within these groups, the majority of sequences were affiliated with Alteromonas bacteria (62.8 %). Other well-represented groups within the γ-Proteobacteria included Marinomonas (4.8 %), Shewanella (7.4 %) and Vibrio species (7.6 %). Roseobacter sp. was dominant within the α-Proteobacteria, representing 5 % of all retrieved sequences. The remaining OTUs within the α-Proteobacteria were affiliated with the bacteria Roseovarius (1.2 %), Rhodospirillales (1.2 %), Thalassospira (1.6 %) and Rhodomicrobium (1.2 %).

Abundance of bacterial OTUs (grouped at 97 % identity) acquired from A. tenuis and P. damicornis spawning and planulating water, respectively. Sequences abundant in proportions >1 % in either species are included
Discussion
This study highlights that corals release bacteria with their offspring, which is a potential method of intergenerational bacterial transmission that may benefit the fitness of coral larvae. The two coral species investigated in this study, a brooder and a spawner, both released similar dominant bacterial types including Flavobacteriaceae, Rhodomicrobium, Roseobacter, Rhodospirillales, Alteromonas and Shewanella, although the affiliations detected occurred in different relative abundance for the two coral species (Fig. 1). This limited number of taxa, similar bacterial profiles and two dominant genera of bacteria in both coral species support the idea that specific bacterial types are released and potentially involved in coral reproduction and, possibly, early and later coral life stages. These identified that bacterial groups display close phylogenetic affiliation to bacteria previously observed to be dominant in other corals and play important roles. For example, bacteria types Roseobacter and Alteromonas, released with gametes and planulae, were previously detected in the tissue of post-reproductive adult corals of the same species, 2 days after spawning and planulation [4]. This consistent retrieval of some bacterial sequences through coral reproduction suggests specific functions and potential benefits through coral reproduction and early coral life. A larger number of brooding and spawning coral species, a bigger sample size and an investigation through several reproduction cycles are needed, however, to further support this hypothesis.
Sequences retrieved from seawater surrounding A. tenuis after reproduction were predominantly affiliated with α-Proteobacteria, whereas high proportions of γ-Proteobacteria affiliates were retrieved from P. damicornis post-reproduction water. The two genera Roseobacter and Alteromonas occurred in the water of both coral species; however, the abundance of these bacteria appeared strikingly dissimilar for the two coral species with 89.7 % Roseobacter in A. tenuis and 62.8 % Alteromonas in P. damicornis spawning water. Both genera are abundant and diverse in seawater, and various metabolic functions have been reported for Roseobacter clade-affiliated species [19], including antibiotic properties against coral pathogens [16]. Other studies reported Alteromonas species to induce coral settlement [27]. Considering that P. damicornis planulae are ready to settle after their release, a high abundance of Alteromonas-affiliated bacteria might be highly beneficial in facilitating larval settlement. Furthermore, fully developed planulae are likely pre-equipped with Roseobacter-affiliated species via vertical transfer from mother colonies which have been shown for planulae of the coral P. astreoides [24]. Spawning coral species, which recruit their bacterial partners from the water column, are more likely to benefit from the defence properties of Roseobacter clade-affiliated species while undergoing their developmental stages into planulae until settlement.
A previous study reported that the abundance of Roseobacter increased in three coral species after a coral mass spawning event on Ningaloo Reef [4], and clone libraries retrieved from A. tenuis and P. damicornis through 1 year [5] demonstrated the presence of Roseobacter clade-affiliated species in three out of four sampling times, together indicating their proposed importance in several coral life stages. Roseobacter bacteria might provide antimicrobial activity in corals compromised by the energy-demanding process of reproduction and prevent the coral from pathogenic bacteria. These protective properties might be equally important in the vulnerable early life stages of larvae, polyps and juvenile corals. Broadcast spawning corals do not transfer bacteria vertically to the next generation, and their larvae travel long distances. If beneficial bacteria are to be successfully transferred to the next generation, they have to be highly abundant and widely dispersed in seawater. During mass spawning events, large numbers of coral species may release substantial quantities of Roseobacter-affiliated taxa with their gametes, facilitating targeted and specific horizontal uptake into the next generation. After a coral mass spawning event on the Great Barrier Reef, the bacterial abundance in the water column increased twofold and returned to pre-spawning levels after 3 days [18].
The brooder P. damicornis releases fully functional and ready-to-settle planulae 1 week after the annual coral mass spawning event, when Roseobacter abundance likely dropped to normal levels. Brooding corals might take advantage of increased Roseobacter abundance in the seawater after coral mass spawning and acquire these bacteria internally for vertical transfer to their offspring and for the time of their release. A recent study has shown the vertical transfer of Roseobacter bacteria in the brooding coral P. astreoides [24]. If Roseobacter is transferred vertically in P. damicornis corals, then compared to spawning corals, lower numbers of Roseobacter are needed to be released by mother colonies to assure the transfer of these bacteria into the next generation. Our data indeed shows that P. damicornis do release much lower proportions in Roseobacter species than A. tenuis. Planulae from brooding corals usually settle within hours of their release and in close proximity to mother colonies, and if bacteria play an essential role in the survival success of coral larvae, the dispersal distance of released bacteria might limit the settlement distance in P. damicornis planulae from mother colonies. Given the lack of data on vertical transmission in P. damicornis though, further investigation on the bacterial components of P. damicornis larvae is required.
Roseobacter-affiliated species are amongst the first colonisers in Pocillopora meandrina [1], a spawning coral species that acquires bacteria horizontally from the environment, showing that transfer of potentially beneficial bacteria to the next generation is not dependent on vertical transmission. A high abundance of bacteria released by the mother colony into the water column during spawning would, therefore, increase the chances of a horizontal and specific transfer of beneficial associates into coral larvae.
The present study is limited by low numbers of replication; nonetheless, it addresses an interesting question concerning the transmission of specific groups of bacteria that have been found across diverse corals and proposes a new hypothesis to explain how these bacteria are maintained across coral generations. Independent of the reproductive strategy, parent corals likely transfer specific groups of bacteria to the next generation which are released during spawning and are taken up by their offspring. This strategy is less specific than intergenerational vertical transfer, however, not as non-specific as random horizontal acquisition of seawater communities. The localised enrichment of specific bacteria is likely to reflect the selection of potentially important bacteria which have an implication for increasing the fitness of coral offspring. The genera Alteromonas and Roseobacter might be the most relevant taxa in coral reproduction and early coral development.
References
Apprill A, Marlow HQ, Martindale MQ, Rappe MS (2009) The onset of microbial associations in the coral Pocillopora meandrina. ISME J 3:685–699
Bragg L, Stone G, Imelfort M, Hugenholtz P, Tyson GW (2012) Fast, accurate error-correction of amplicon pyrosequences using Acacia. Nat Methods 9:425–426
Caporaso G, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK (2010) QIIME allows analysis of high-throughput community sequencing data. Nature 7:335–336
Ceh J, Raina JB, Soo MR, and Bourne DG (2012) Coral-bacterial communities before and after a coral mass spawning event on Ningaloo Reef. PLoSONE
Ceh J, van Keulen M, Bourne DG (2011) Coral-associated bacterial communities on Ningaloo Reef, Western Australia. FEMS Microbiol Ecol 75:134–144
DeSantis TZ, Hugenholtz P, Larsen N, Rojas M, Brodie EL, Keller K, Huber T, Dalevi D, Hu P, and Andersen GL (2006) Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol 5069–5072
Edgar RC (2010) Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2460–2461
Edgar RC, Haas BJ, Clemente JC, Quince C, and Knight R (2011) UCHIME improves sensitivity and speed of chimera detection. Bioinformatics
Gower JC, Dijksterhuis GB (2004) Procrustes problems. Oxford University Press, New York
Hirose E (2000) Plant rake and algal pouch of the larvae in the tropical ascidian Diplosoma similis: an adaptation for vertical transmission of photosynthetic symbionts Prochloron sp. Zool. Science 17:233–240
Knowlton N, Rohwer F (2003) Multispecies microbial mutualisms on coral reefs: the host as a habitat. Am Nat 162:52–62
Krueger DM, Gustafson RG, Cavanaugh CM (1996) Vertical transmission of chemoautotrophic symbionts in the bivalve Solemya velum (Bivalvia: Protobranchia). Biol Bull 190:195–202
Lema KA, Willis BL, and Bourne DG (2012) Corals form characteristic associations with symbiotic nitrogen-fixing bacteria. Appl. Environ. Microbiol
Lesser MP, Mazel CH, Gorbunov MY, Falkowski PG (2004) Discovery of symbiotic nitrogen-fixing cyanobacteria in corals. Science 305:997–1000
McFall-Ngai MJ, Ruby EG (1998) Squids and vibrios: when they first meet. BioScience 48:257–265
Nissimov J, Rosenberg E, Munn CB (2009) Antimicrobial properties of resident coral mucus bacteria of Oculina patagonica. FEMS Microbiol Letters 292:210–215
Nyholm SV, McFall-Ngai MJ (2004) The winnowing: establishing the squid-vibrio symbiosis. Nature Rev Microbiol 2:632–642
Patten NL, Mitchell JG, Middelboe M, Eyre BD, Seuront L, Harrison PL et al (2008) Bacterial and viral dynamics during a mass coral spawning period on the Great Barrier Reef. Aquat Microbiol Ecol 58:209–220
Piekarski T, Buchholz I, Schobert M, Wagner-Doebler I, and Tielen P (2009) Genetic tools for the investigation of Roseobacter clade bacteria. Bmc Microbiol 9
Richmond RH, Hunter CL (1990) Reproduction and recruitment of corals:comparisons among the Caribbean, the tropical Pacific and the Red Sea. Mar Ecol Prog Ser 60:185–203
Ritchie KB (2006) Regulation of microbial populations by coral surface mucus and mucus-associated bacteria. Mar Ecol Prog Ser 322:1–14
Rosenberg E, Koren O, Reshef L, Efrony R, Zilber-Rosenberg I (2007) The role of microorganisms in coral health, disease and evolution. Nat Rev Microbiol 5:355–362
Schauer M, Massana R, Pedrós-Alió C (2000) Spatial differences in bacterioplankton composition along the Catalan coast (NW Mediterranean) assessed by molecular fingerprinting. FEMS Microbiol Ecol 33:51–59
Sharp KH, Distel DL, and Paul VJ (2011) Diversity and dynamics of bacterial communities in early life stages of the Caribbean coral Porites astreoides. ISME J
Sharp KH, Eam BE, Faulkner DJ, Haygood MG (2007) Vertical transmission of diverse microbes in the tropical sponge Corticium sp. Appl Environm Microbiol 73:622–629
Sharp KH, Ritchie KB, Schupp PJ, Ritson-Williams R, Paul VJ (2010) Bacterial acquisition in juveniles of several broadcast spawning coral species. PLoSONE 5:e10898
Webster NS, Smith LD, Heyward AJ, Watts JEM, Webb RI, Blackall LL et al (2004) Metamorphosis of a scleractinian coral in response to microbial biofilms. Appl Environm Microbiol 70:1213–1221
Acknowledgments
We would like to thank the Australian Institute of Marine Science, the Western Australian Marine Science Institution and Murdoch University for their contributions to this research. Paul Dennis from the University of Queensland is thanked for his help with passing the pyrotag sequence dataset through the programme ACACIA. We also thank Frazer McGregor and Bernard O’Reilly for their help in the field.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ceh, J., van Keulen, M. & Bourne, D.G. Intergenerational Transfer of Specific Bacteria in Corals and Possible Implications for Offspring Fitness. Microb Ecol 65, 227–231 (2013). https://doi.org/10.1007/s00248-012-0105-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-012-0105-z