
Abstract
Aquatic hyphomycetes play an essential role in the decomposition of allochthonous organic matter which is a fundamental process driving the functioning of forested headwater streams. We studied the effect of anthropogenic acidification on aquatic hyphomycetes associated with decaying leaves of Fagus sylvatica in six forested headwater streams (pH range, 4.3–7.1). Non-metric multidimensional scaling revealed marked differences in aquatic hyphomycete assemblages between acidified and reference streams. We found strong relationships between aquatic hyphomycete richness and mean Al concentration (r = −0.998, p < 0.0001) and mean pH (r = 0.962, p < 0.002), meaning that fungal diversity was severely depleted in acidified streams. By contrast, mean fungal biomass was not related to acidity. Leaf breakdown rate was drastically reduced under acidic conditions raising the issue of whether the functioning of headwater ecosystems could be impaired by a loss of aquatic hyphomycete species.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Running-water ecosystems that are subjected to anthropogenic acidification are typically first- to third-order streams, i.e. headwater streams often draining forested watersheds. Due to several ecological factors, such as light limitation by the riparian vegetation, the primary production in these streams is very low. Conversely, forested headwater streams receive by far most of their energy from the riparian vegetation. Therefore, the metabolism of headwater streams is generally heterotrophic [16, 67]. Leaf litter, which represents an important fraction of the allochthonous input entering the ecosystems, is used by decomposers and detritivores. Micro-organisms and leaf-shredding macro-invertebrates play an essential role in allochthonous organic matter processing and its incorporation into the trophic webs [31, 68]. Aquatic fungi, particularly aquatic hyphomycetes which form a phylogenetically diverse group of anamorphous species (mainly ascomycetes and few basidiomycetes), contribute the most to the microbial biomass associated with leaves in streams [26, 38, 69]. These filamentous fungi commonly occur in freshwaters and produce tetraradiate, variously branched or sigmoid asexual spores (i.e. conidia) adapted to dispersal in flowing water and adhesion to decaying plant tissues [7]. A rapid conidial germination and mycelial invasion of the leaf matrix, typically within a few weeks, allow aquatic hyphomycetes to quickly colonize their ephemeral resource [32]. In addition, the mycelial development coincides with or is closely followed by a very high release of conidia which constitutes a major part of the fungal production [32]. Aquatic hyphomycetes are responsible for the initial breakdown of leaves and for enhancing the palatability of leaf detritus, thus providing a more suitable food source for shredders which further contribute to the conversion of leaf litter into fine particles and other decomposition products [3, 4, 8, 10, 15, 31, 34, 61]. Consequently, due to their function, aquatic hyphomycetes occupy an essential place in the functional role that leaf breakdown plays in forested headwater streams.
Several studies have clearly shown that chemical alteration of aquatic ecosystems can have deleterious effects on fungal communities. A decline in aquatic hyphomycete diversity has been reported in streams contaminated with either heavy metals [9, 22] or organic compounds [5, 59]. Similarly, leaf breakdown rates are often affected in rivers polluted with either mineral or organic pollutants (for review, see [29]). In particular, several studies have clearly demonstrated that leaf-litter breakdown is severely reduced in acidified ecosystems [19, 53].
During the last three decades, a large number of studies have focused on the effects of acidification on aquatic biota (fish, macroinvertebrates, zooplankton, macrophytes, algae). Most of them have revealed deleterious effects leading to the loss of biodiversity. In particular, increased concentrations of aqueous aluminium arising from freshwater acidification are considered a major environmental problem due to its high toxicity to aquatic organisms. Aqueous aluminium has been recognized as a main toxicant for aquatic animals [27], and several studies have also suggested a direct effect on microbial metabolism [12, 55, 57]. Surprisingly and despite the key role they play in headwater stream functioning, little is known about the impact of anthropogenically acidified running waters on the diversity of microorganisms in general and on aquatic hyphomycetes in particular. Results from the few studies dealing with acidic water and the diversity of aquatic hyphomycetes are contradictory. In some studies, aquatic hyphomycetes have been shown to benefit from lower pH [6, 70], whereas in other studies, the richness was lower [44, 60] or not related to pH [10]. Obviously, additional data are needed to more fully understand the effects of low-pH waters on fungi colonizing leaves in streams [62].
To answer the question of whether acidified soft waters with high aluminium concentrations affect the diversity of aquatic hyphomycete assemblages associated with decaying leaves, a litter-breakdown experiment was conducted in six streams characterized by different acidification levels. After immersion in the streams, the leaf mass remaining was determined, and the sporulation of the leaf detritus was induced in the laboratory, allowing the identification of fungal species through released conidia. The relationship between diversity and breakdown rate was investigated to link the potential effect of species loss on leaf-litter processing in headwater streams.
Materials and Methods
Study Sites
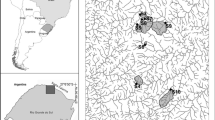
The study was conducted in the Vosges Mountains (Northeastern France) where anthropogenic acidification has adversely affected surface waters (Fig. 1) [17, 58]. Based on previous studies [19, 20, 25, 66], six headwater streams showing different levels of acidity (Table 1) but similar hydrological and morphological characteristics were selected. Soils of all catchments range from acid brown to podzolic and are underlain by granite or sandstone bedrock. Mixed forests dominated by silver fir (Abies alba), Norway spruce (Picea abies) and common beech (Fagus sylvatica) form the riparian zones. All sampling sites were located upstream of any direct anthropogenic pressures (housing, farming or industries).

Map showing the location of the six headwater streams studied in the Vosges Mountains
Physical and Chemical Analyses
At each sampling date, water samples were collected from each stream into polyethylene bottles and kept cold (4°C) until laboratory and analysed within 48 h. Stream pH was measured in the laboratory using a microprocessor pH meter (pH 3000, WTW), and acid-neutralizing capacity (ANC) was determined by Gran’s titration. Conductivity was measured with a Metrohm Herisau Conductometer E518 (Herisau, Switzerland) at 25°C. Concentrations of Ca2+, Mg2+, Na+, K+ and total aluminium (after acidification with HNO3) were determined by atomic absorption spectrophotometry (Aanalyst 100; Perkin Elmer and Varian SpectrAA-300) and concentrations of Cl−, \({\text{SO}}_4^{2 - } \) and \({\text{NO}}_3^ - \) by ion chromatography (Dionex 1500i with a AS 4 A SC column; Sunnyvale, USA).
Leaf-Litter Breakdown
We used beech (F. sylvatica) leaves because this species is by far the most common deciduous riparian tree species and provides almost all leaf-litter input. Beech leaves were collected from trees just before abscission in fall 2002. Leaf bags were created by placing 5 g (±0.05) of air-dried leaves in 10 mm plastic mesh bags (15 × 20 cm). Each exact mass of leaf material was recorded and each bag identified with a plastic label. Weighed leaves were moistened with distilled water to avoid breakage during placement into mesh bags. Leaf bags were closed with nylon line so that they took the shape of a tetrahedron thus ensuring a good circulation flow and a natural arrangement of leaves. About 24 bags were submerged at each study site in zones expected to naturally accumulate leaves. They were individually secured to the bank with plastic-coated wire, which was anchored to the stream bottom with large boulders. Four replicate bags were randomly retrieved from the six streams after 23, 35, 46, 63, 83 and 124 days of exposure, immediately placed in individual sealed plastic bags with stream water and transported to the laboratory in a cool box. For each sampling date, 7 l of stream water was collected for laboratory processing of the leaf bags.
The leaves were rinsed individually with water from the respective stream to remove attached fine particles and invertebrates. Two leaf disks were cut avoiding the central vein with a 12 mm diameter cork borer from five randomly selected leaves in each bag. Five of them were stored at −80°C in a zip-lock plastic bag until ergosterol determination, and the five others were immediately processed to allow sporulation. The remaining leaf material was oven-dried to constant mass (105°C, 48 h) and weighed to the nearest 0.1 mg, and subsamples (500 mg) were ignited in a muffle furnace (550°C, 4 h) to relate dry mass to ash-free dry mass (AFDM). Ten additional leaf bags were kept in the laboratory before the beginning of the experiment. Four replicates were used to determine leaching loss by submerging them in a bowl under a slow flow of dechlorinated water for 24 h and six others to estimate the initial oven-dried mass and AFDM of all leaf bags.
Fungal Biomass
Ergosterol was extracted from leaf disks and determined as previously described [33]. Briefly, the leaf disks were lyophilised, weighed to the nearest 0.1 mg and heated in 5 ml hot alkaline ethanol (KOH, 8 g l−1) for 30 min at 80°C. The extract was purified by solid-phase extraction on cartridges (Waters Oasis HLB, 60 mg, 3 cc). Ergosterol was separated by reversed phase high performance liquid chromatography on C18 and quantified by measuring absorbance at 282 nm. Ergosterol was converted to fungal biomass using a conversion factor of 5.5 mg ergosterol g−1 mycelial dry mass [30].
Fungal Diversity
The sporulation rate and species composition of the leaf-associated fungal assemblages were determined in the laboratory [28]. Five leaf discs from each replicate bag were placed in 100-ml Erlenmeyer flasks with 25 ml filtered water (Whatman glass fiber filter GF/F) from the stream in which the leaves were retrieved. Then, the flasks were placed on an orbital shaker (100 rpm) for 48 h at 10°C. After incubation, the suspension was poured into 50-ml centrifuge tube without the discs, which were rinsed in the flask with distilled water (3 × 2 ml) to dislodge remaining attached conidia. Rinse water was combined in the tube, and the volume was adjusted to 35 ml with distilled water and 2 ml of 37% formalin. Conidial suspensions were stored in the dark until analysis. The five leaf discs from each leaf pack were oven-dried (105°C, 48 h) and weighed to the nearest 0.1 mg, and AFDM was determined (550°C, 4 h).
The conidial suspension was transferred to a beaker with 1 ml Triton X-100 solution (0.5%) and shaken on a magnetic stirrer for 10 min, and an aliquot (4–8 ml) was filtered (membrane filter, 5-mm pore size, 25 mm diameter), stained with 0.1% Trypan blue in 60% lactic acid and examined microscopically. Conidia were identified by relying on specific literature [e.g. 39] and counted by scanning the entire filter. For each species, the sporulation rate (conidia mg−1 AFDM day−1) was determined.
Data Analysis
Principal components analysis (PCA) was carried out to ordinate streams with respect to physical and chemical variables. PCA was performed using the following variables: pH, ANC, conductivity, total Al, [NO3 + SO4] and [Ca + Mg].
Non-metric multidimensional scaling (NMDS) analysis of sporulation data was used to assess differences among sites in aquatic hyphomycete assemblages. This ordination method is a robust procedure for analysing ecological data [52]. We used the Bray-Curtis coefficient to quantify the dissimilarity among sites based on joint occurrence and abundance of taxa [14]. NMDS attempts to maximise the fit between measured dissimilarities and distance between resulting data points within a predefined number of spatial dimensions [46, 47]. The goodness of fit was estimated with a stress function, which ranges from 0 to 1, with values close to zero indicating a good fit. Axes from the NMDS analysis were correlated (Spearman rank correlation) with physical and chemical data to identify variables most strongly corresponding to among-site differences in aquatic hyphomycete assemblages [41].
Leaf breakdown rates (k) were estimated by fitting the AFDM data with the linear model m t = m 0 − kt where m t is the AFDM remaining at time t, and m 0 is the initial AFDM at the beginning of the experiment. The linear model was used because it fitted the data better than the exponential model proposed by Petersen and Cummins [56]. Daily decay rates were compared using analysis of covariance (ANCOVA) followed by a multiple-comparison (Tukey’s test, [72]).
A one-way analysis of variance (ANOVA) was used to test for differences in fungal biomass, and Pearson correlations were performed to investigate possible relationships between diversity and physical and chemical parameters.
Statistical analyses were performed using ADE4 (PCA, [63]), SPSS for windows (NMDS) and Statistica (ANOVA, ANCOVA, Spearman rank and Pearson correlations).
Results
Water Chemistry
The six headwater streams ranged from near neutral [mean pH = 7.1 and 6.6 in La Maix (LM) and Bihet (BH), respectively] to strongly acidified (mean pH = 4.6 and 4.3 in Gentil Sapin (GS) and Basse Escaliers (BE), respectively). The first factorial plane of the PCA explained 91.4% of the total variance (Fig. 2). The first axis of the PCA was defined primarily by the ANC and Ca + Mg (Fig. 2a). It explained 68.6% of the total variance and strongly separated the circumneutral streams (LM and BH) from the acidified ones [GS, BE, Wassongoutte (WA), Longfoigneux (LG); Fig. 2b). An additional 22.8% of the total variance was explained by the second axis, which discriminated among streams draining catchments underlain by sandstone vs granite bedrock. Streams draining granite catchments (BH, WA, LG) were mainly characterized by lower concentrations of strong acid anions. In addition, acidified streams draining sandstone (BE, GS) exhibited higher total Al concentrations than acidified streams draining granite (WA, LG).

PCA performed on the physico-chemical variables in the six headwater streams during the leaf breakdown experiment. a Correlation circle on the F1xF2 factorial plane. b Ordination of the six sites on the F1 × F2 factorial plane. Small squares represent the sample position at each sampling date. Circles denote the weighted average of all samples taken from a given stream. Lines link samples to their weighted average
Fungal Diversity and Mycelial Biomass
A total of 37 species of aquatic hyphomycetes was observed during the leaf-litter breakdown experiment (Table 2). The higher fungal diversity was found in the two circumneutral streams BH and LM (27 and 25 species, respectively) and the lowest in BE (seven taxa) which was also the most acidic one. Approximately 17 species were never recorded in acidic streams. Among these species, eight were shared by the two circumneutral streams BH and LM: Alatospora pulchella, Culicidospora gravida, Lemonniera aquatica, Tricladium chaetocladium, Tricladium splendens, Tumularia aquatica, Tumularia tuberculata and Ypsilina graminea. The nine other species were observed only in one of the circumneutral streams, LM or BH. On the other hand, only one species, Casaresia sphagnorum, was recorded in the four acidic streams and never in the circumneutral ones.
During beech-leaf processing, Clavariopsis aquatica was the dominant species in the two circumneutral streams (i.e. 83.4% of the conidia produced in LM and 49.5% in BH), and Flagellospora curvula was dominant in the four acidic streams (i.e. from 83.1% in WA to 98.9% in BE).
The NMDS ordination described well the overall differences in aquatic hyphomycete assemblages between the six streams (Fig. 3). Stress was very low (0.003), indicating a high degree of representation. Axis 1 of the NMDS was strongly correlated (Spearman rank correlation) with pH (r = 0.89; p < 0.05), ANC (r = 0.89; p < 0.05) and total aluminium concentration (r = −0.89; p < 0.05). Position along this axis reflected the acidification status of the streams and strongly separated the acidic streams with low fungal richness (Table 2) from the circumneutral streams with markedly higher richness. Axis 2 correlated best with [\({\text{SO}}_4^{2 - } + {\text{NO}}_3^ - \)] concentrations (r = 0.83; p < 0.05) and separated streams according to the bedrock of their catchments.

NMDS plot based on hyphomycete assemblages associated with decaying leaves in the six headwater streams. Circumneutral streams are shown in white and acidic streams in black. Squares represent streams draining granite bedrock and circles streams draining sandstone bedrock
Pearson correlation analyses between physical and chemical parameters and fungal species richness revealed significant relationships (Fig. 4) only with mean total aluminium concentration (log10 transformed; r = −0.998, p < 0.0001) and mean pH (r = 0.962; p < 0.002).

Relationship between fungi richness and a mean total aluminium concentration, b mean pH
When considering fungal biomass, no clear tendency was observed, and mean values did not differ significantly between streams (one-way ANOVA; F = 1.352, p = 0.271), demonstrating that fungal biomass associated with decaying leaves was not affected by acidification (Fig. 5).

Mean (+SD) fungi biomass in each stream. Streams are arranged following decreasing pH from the left to the right
Breakdown Rates
Breakdown rates of beech leaves were significantly different among streams (ANCOVA; F = 41.9, p < 0.001) and ranged from 0.0045 day−1 in the most acidic site (BE) to 0.0285 day−1 in the circumneutral stream BH (Fig. 6; Table 3). The breakdown rate in the circumneutral stream draining granite (BH) was significantly faster than that observed in the circumneutral stream draining sandstone (LM). However, both streams exhibited faster breakdown rates than acidic streams, while no significant differences occurred among the four acidic streams. After 124 days of exposure in these acidic streams, the mass loss did not exceed 16%, indicating that leaf-litter breakdown in these headwater streams had virtually stopped.

Mean (±SD) percent of beech leaf remaining AFDM in the six streams
Discussion
Acidification of ecosystems resulting from atmospheric pollution is one of the most revealing demonstrations that human activities can strongly impair terrestrial and aquatic ecosystems located in remote areas. Among the most striking consequences of freshwater acidification, the loss of biodiversity has been well documented for different taxonomic groups such as fish [42, 48], invertebrates [21, 37], algae [51, 54] and macrophytes [23]. By comparison, the effect of anthropogenic acidification of aquatic ecosystems on the diversity of micro-organisms in general and aquatic hyphomycetes in particular has received much less attention despite their importance at multiple ecological scales. The few studies that have attempted to link aquatic hyphomycete richness and acidity have provided very contradictory results. For instance, in a study of aquatic hyphomycetes in ten streams of New Brunswick and Nova Scotia, Bärlocher [6] concluded that “their tolerance of low pH values makes them one of the rare groups of stream organisms that may actually benefit from the effects of acid rain”. This conclusion contrasts markedly with the results of the previous studies of Iqbal and Webster [44] and Shearer and Webster [60] in Devon who reported impoverished fungal diversity in upland acidic water compared to lowland circumneutral sites, whereas Chamier [10] found low species richness in upland streams regardless of pH (4.9–6.8) in seven streams of the English Lake District.
In our studies, we observed a severe depletion of species in acidified streams (from 44 to 75%), and more than 45% of the species associated with decaying leaves in circumneutral streams were never recorded in acidified streams. The species richness was strongly correlated with aluminium concentration and pH. However, because mineral acidity is the main factor regulating Al concentrations in acid surface waters, Al is more likely to control the diversity of aquatic hyphomycetes than H+. In experiments to test for the effect of Al on aquatic hyphomycetes, Chamier and Tipping [12] found that deleterious effect of Al depend on the species considered, the effects being more marked on species common in circumneutral streams such as T. splendens. Similarly, differences in sensitivity to other metals among aquatic hyphomycete species have also been reported from experiments performed to assess the toxicity of Cd [1].
Several hypotheses can be proposed to explain these discrepancies. Chemical parameters such as Al were not analysed in earlier studies, and the type of acidity (natural, i.e. organic vs anthropogenic) was not considered. Therefore, the potential toxicity of water can markedly differ even in streams with similar pH. Furthermore, the results provided by these previous studies of species richness in relation to pH are difficult to compare as several of them have been performed in upland and lowland streams and/or in forested streams and streams without riparian trees.
In our study, the acidity of headwater streams did not originate from organic acidity (DOC < 2.3 mg l−1), all the streams were forested (mainly A. alba, P. abies and F. sylvatica) and exhibited very similar environmental characteristics, allowing us to compare fungal diversity and species composition and to investigate the effects of anthropogenic acidification. Our results show that in the Vosges mountains, aquatic hyphomycete communities are severely impoverished, as are macroinvertebrate communities, as established in previous studies [37, 66]. In these streams, a combined toxic effect of high protons and aluminium concentrations has been demonstrated for different species of invertebrates [24, 25] as commonly reported for other aquatic organisms such as fish [50], crayfish [2], macrophytes [23] and algae [13, 40].
It is interesting to note that fungal biomass associated with decaying leaves was not affected by acidification, thus corroborating the results from Dangles and Chauvet [18] in five streams from the Vosges Mountains but contrasting markedly with the findings of Griffith and Perry [35], who found significantly reduced fungal biomass associated with decomposing oak leaves in a strongly acidified West Virginia stream. Such a discrepancy illustrates the fact that leaf-mass loss is no straightforward indicator of the fungal activities. A possible explanation for the significant differences in leaf breakdown rates despite similar fungal biomasses in our study lies in the sensitivity of pectin lyases to water chemistry. The enzymatic degradation of pectin represents a key process in leaf-mass loss [45], and different studies have shown that the activities of pectin lyases are higher in neutral or alkaline waters than in acid or soft waters [11, 36, 45]. Moreover, Chamier and Tipping [12] have demonstrated that treatment with monomeric Al decreased pectinase production by four aquatic hyphomycete species.
Not surprisingly, our results showed that leaf-litter breakdown was severely depressed under acidic conditions, as previously reported in different studies [10, 19, 20, 36, 53]. In a recent study of 25 woodland first- and second-order streams along an acidification gradient in the Vosges Mountains (France), Dangles et al. [19] found that breakdown rates of beech (F. sylvatica) leaves varied more than 20-fold between the most acidified and circumneutral sites (k = 0.0002–0.0055 day−1) consistently with the associated fungal biomass and microbial respiration. Total abundance, biomass and species richness of leaf-shredding invertebrates associated with decomposing leaves were, however, not related to stream acidity (the diversity of aquatic hyphomycetes was unfortunately not investigated in this study) which suggests that fungi played a prominent role in acidified streams. That the slow breakdown rates we have measured in acidified headwater streams is linked to the decreased aquatic hyphomycete diversity is questionable. Several experimental studies have provided evidence that the loss of species can affect the efficiency with which resources are processed within an ecosystem [43, 49, 65] although the mechanisms underlying the response are not always well understood [71]. Whether diversity of aquatic hyphomycetes interferes with their functional role in leaf breakdown in the context of the present study should deserve special attention, e.g. through complementary in vitro and field experiments [e.g. 64]. Potential implications of such relationships are of major importance for higher trophic levels within the stream ecosystem.
References
Abel TH, Bärlocher F (1984) Effects of cadmium on aquatic hyphomycetes. Appl Environ Microbiol 48:245–251
Appelberg M (1985) Changes in haemolymph ion concentrations of Astacus astacus L. and Pacifastacus leniusculus (Dana) after exposure to low pH and aluminum. Hydrobiologia 121:19–25
Arsuffi TL, Suberkropp K (1985) Selective feeding by caddisfly (Trichoptera) detritivores on leaves with fungal colonized patches. Oikos 45:50–58
Arsuffi TL, Suberkropp K (1989) Selective feeding by shredders on leaf-colonizing stream fungi: comparison of macroinvertebrate taxa. Oecologia 79:30–37
Au DWT, Vrijmoed LLP, Hodgkiss IJ (1992) Fungi and cellulolytic activity associated with decomposition of Bauhinia purpurea leaf litter in a polluted and unpolluted Hong Kong waterway. Can J Bot 70:1071–1079
Bärlocher F (1987) Aquatic hyphomycete spora in 10 streams of New Brunswick and Nova Scotia. Can J Bot 65:76–79
Bärlocher F (1992) Research on aquatic hyphomycetes: historical background and overview. In: Bärlocher F (ed) The ecology of aquatic hyphomycetes. Springer, Berlin, pp 1–15
Bärlocher F (2005) Freshwater fungal communities. In: Dighton J, White JF, Oudemans P (eds) The fungal community. Its organization and role in the ecosystem. 3rd edn. CRC, Boca Raton, FL, pp 39–59
Bermingham S, Maltby L, Cooke R (1996) Effects of a coal mine effluent on aquatic hyphomycetes. I. Field Study. J Appl Ecol 33:1311–1321
Chamier AC (1987) Effect of pH on microbial degradation of leaf litter in seven streams of the English Lake District. Oecologia 71:491–500
Chamier AC (1992) Water chemistry. In: Bärlocher F (ed) The ecology of aquatic hyphomycetes. Springer, Berlin, pp 152–172
Chamier AC, Tipping E (1997) Effects of aluminium in acid streams on growth and sporulation of aquatic hyphomycetes. Environ Pollut 96:289–298
Claesson A, Törnqvist L (1988) The toxicity of aluminium to two acido-tolerant green algae. Water Res 22:977–983
Clarke KR, Warwick RM (1994) Change in marine communities: an approach to statistical analysis and interpretation. Natural Environment Research Council, Plymouth, UK, p 234
Covich AP, Palmer MA, Crowl TA (1999) The role of benthic invertebrate species in freshwater ecosystems. Bioscience 49:119
Cummins KW, Wilzbach MA, Gates DM, Perry JB, Taliaferro WB (1989) Shredders and riparian vegetation. Bioscience 39:24–30
Dambrine E, Pollier B, Poszwa A, Ranger J, Probst A, Viville D, Biron P, Granier A (1998) Evidence of current soil acidification in spruce stands in the vosges mountains, north-eastern France. Water Air Soil Pollut 104:31–41
Dangles O, Chauvet E (2003) Effects of stream acidification on fungal biomass in decaying beech leaves and leaf palatability. Water Res 37:533–538
Dangles O, Gessner MO, Guérold F, Chauvet E (2004) Impact of stream acidification on litter breakdown: implication for assessing ecosystem functioning. J Appl Ecol 41:365–378
Dangles O, Guerold F (2001) Influence of shredders in mediating breakdown rates of beech leaves in circumneutral and acidic forest streams. Arch Hydrobiol 151:649–666
Driscoll CT, Lawrence GB, Bulger AJ, Butler TJ et al (2001) Acidic deposition in the Northeastern United States: Sources and inputs, ecosystem effects, and management strategies. Bioscience 51:180
Duarte S, Pascoal C, Cássio F (2004) Effects of zinc on leaf decomposition by fungi in streams: studies in microcosms. Microb Ecol 48:366–374
Farmer AM (1990) The effects of lake acidification on aquatic macrophytes—a review. Environ Pollut 65:219–240
Felten V, Guérold F (2004) Haemolymph [Na+] and [Cl−] loss in Gammarus fossarum exposed in situ to a wide range of acidic streams. Dis Aquat Org 61:113–121
Felten V, Guérold F (2006) Short-term physiological responses to a severe acid stress in three macroinvertebrate species: a comparative study. Chemosphere 63:1427–1435
Findlay S, Tank J, Dye S, Valett HM, Mulholland PJ, McDowell WH, Johnson SL, Hamilton SK, Edmonds J, Dodds WK, Bowden WB (2002) A cross-system comparison of bacterial and fungal biomass in detritus pools of headwater streams. Microb Ecol V43:55–66
Gensemer RW, Playle RC (1999) The bioavailability and toxicity of aluminum in aquatic environments. Crit Rev Environ Sci Technol 29:315–450
Gessner MO, Bärlocher F, Chauvet E (2003) Qualitative and quantitative analyses of aquatic hyphomycetes in streams. Fungal Divers Res Ser 10:127–157
Gessner MO, Chauvet E (2002) A case for using litter breakdown to assess functional stream integrity. Ecol Appl 12:498–510
Gessner MO, Chauvet E (1993) Ergosterol-to-biomass conversion factors for aquatic hyphomycetes. Appl Environ Microbiol 59:502–507
Gessner MO, Chauvet E, Dobson M (1999) A perspective on leaf litter breakdown in streams. Oikos 85:377–384
Gessner MO, Gulis V, Kuehn KA, Chauvet E, Suberkropp K (2007) Fungal decomposers of plant litter in aquatic ecosystems. In: Kubicek CP, Druzhinina IS (eds) The Mycota IV: Microbial and Environmental Relationships. 2nd edn. Springer-Verlag, Berlin, pp. 301–324
Gessner MO, Schmitt AL (1996) Use of solid-phase extraction to determine ergosterol concentrations in plant tissue colonized by fungi. Appl Environ Microbiol 62:415–419
Golladay SW, Webster JR, Benfield EF (1983) Factors affecting food utilization by a leaf shredding aquatic insect: leaf species and conditioning time. Holarct Ecol 6:157–162
Griffith MB, Perry SA (1994) Fungal biomass and leaf-litter processing in streams of different water chemistry. Hydrobiologia 294:51–61
Griffith MB, Perry SA, Perry WB (1995) Leaf litter processing and exoenzyme production on leaves in streams of different pH. Oecologia 102:460–466
Guérold F, Boudot JP, Jacquemin G, Vein D, Merlet D, Rouiller J (2000) Macroinvertebrate community loss as a result of headwater stream acidification in the Vosges Mountains (N-E France). Biodiv Conserv 9:767–783
Gulis V, Suberkropp K (2003) Effect of inorganic nutrients on relative importance of fungi and bacteria in carbon flow from submerged decomposing leaf litter. Microb Ecol 45:11–19
Gulis V, Marvanová L, Descals E (2005) An illustrated key to the common temperate species of aquatic hyphomycetes. In: Graça MAS, Bärlocher F, Gessner MO (eds) Methods for studying litter decomposition. Kluwer Academic, Dordrecht, pp 130–153
Havens KE, Decosta J (1987) The role of aluminium contamination in determining phytoplankton and zooplankton responses to acidification. Water Air Soil Pollut V33:277–293
Hawkins CP, Hogue JN, Decker LM, Feminella JW (1997) Channel morphology, water temperature, and assemblage structure of stream insects. J North Am Benthol Soc 16:728–749
Heard RM, Sharp WE, Carline RF, Kimmel WG (1997) Episodic acidification and changes in fish diversity in Pennsylvania headwater streams. Trans Am Fish Soc 123:977–984
Hector A, Schmid B, Beierkuhnlein C, Caldeira MC, Diemer M, Dimitrakopoulos PG, Finn JA, Freitas H, Giller PS, Good J, Harris R, Högberg P, Huss-Danell K, Joshi J, Jumpponen A, Körner Ch, Leadley PW, Loreau M, Minns A, Mulder CPH, O’Donovan G, Otway SJ, Pereira JS, Prinz A, Read DJ, Scherer-Lorenzen M, Schulze E-D, Siamantziouras A-SD, Spehn EM, Terry AC, Troumbis AY, Woodward FI, Yachi S, Lawton JH (1999) Plant diversity and productivity experiments in European grasslands. Science 286:1123–1127
Iqbal SH, Webster JR (1977) Aquatic hyphomycete spora of some Dartmoor streams. Trans Brit Mycol Soc 69:233–241
Jenkins CC, Suberkropp K (1995) The influence of water chemistry on the enzymatic degradation of leaves in streams. Freshw Biol 33:245–253
Legendre P, Legendre L (1998) Numerical Ecology. Elsevier, Amsterdam
Legendre P, Marti JA (1999) Distance-based redundancy analysis: testing multispecies responses in multifactorial ecological experiments. Ecol Monogr 69:1–24
Leivestad H, Muniz IP (1976) Fish kill at low pH in a Norwegian river. Nature 259:391–392
Loreau M, Naeem S, Inchausti P (2002) Biodiversity and ecosystems functioning: synthesis and perspectives. Oxford University Press, Oxford (UK)
Masson N, Guérold F, Dangles O (2002) Use of blood parameters in fish to assess acidic stress and chloride pollution in French running waters. Chemosphere 47:467–473
Meegan SK, Perry SA (1996) Periphyton communities in headwater streams of different water chemistry in the Central Appalachian Mountains. J Freshw Ecol 11:247–256
Minchin PR (1987) An evaluation of the relative robustness of techniques for ecological ordination. Vegetatio 69:89–107
Mulholland PJ, Driscoll CT, Elwood JW, Osgood MP, Palumbo AV, Rosemond AD, Smith ME, Schofield CL (1992) Relathionships between stream acidity and bacteria, macroinvertebrates, and fish: a comparison of north temperate mountain streams, USA. Hydrobiologia 239:7–24
Muller P (1980) Effects of artificial acidification on the growth of periphyton. Can J Fish Aquat Sci 37:355–363
Myrold DD, Nason GE (1992) Effect of acid rain on soil microbial processes. In: Mitchell R (ed) Environmental microbiology. Wiley–Liss, New York, pp 59–81
Petersen RC, Cummins KW (1974) Leaf processing in a woodland stream. Freshw Biol 4:343–368
Pina RG, Cervantes C (1996) Microbial interactions with aluminium. BioMetals 9:311–316
Probst A, Party JP, Fevrier C, Dambrine E, Thomas AL, Stussi JM (1999) Evidence of springwater acidification in the Vosges Mountains (North-East of France): Influence of bedrock buffering capacity. Water Air Soil Pollut 114:395–411
Raviraja NS, Sridhar KR, Bärlocher F (1998) Breakdown of ficus and eucalyptus leaves in an organically polluted river in India: fungal diversity and ecological functions. Freshw Biol 39:537–545
Shearer CA, Webster JR (1985) Aquatic hyphomycete communities in the River Teign. I. Longitudinal distribution patterns. Trans Brit Mycol Soc 84:489–501
Suberkropp K (1998) Microorganisms and organic matter processing. In: Naiman RJ, Bilby RE (eds) River ecology and management: lessons from the Pacific Coastal Ecoregion. Springer, Berlin, pp 120–143
Suberkropp K (2001) Fungal growth, production, and sporulation during leaf decomposition in two streams. Appl Environ Microbiol 67:5063–5068
Thioulouse J, Chessel D, Dolédec S, Olivier JM (1997) ADE-4: a multivariate analysis and graphical display software. Stat Comput 7:75–83
Thompson PL, Bärlocher F (1989) Effect of pH on leaf breakdown in streams and in the laboratory. J North Am Benthol Soc 8(3):203–210
Tilman D, Reich P, Knops J, Wedin D, Mielke T, Lehman C (2001) Diversity and productivity in a long-term grassland experiment. Science 294:843–845
Tixier G, Guérold F (2005) Plecoptera response to acidification in several headwater streams in the Vosges Mountains (northeastern France). Biodiv Conserv 14:1525–1539
Vannote RL, Minshall GW, Cummins JR, Sedell JR, Cushing CE (1980) The river continuum concept. Can J Fish Aquat Sci 37:130–137
Webster JR, Benfield EF (1986) Vascular plant breakdown in freshwater ecosystems. Ann Rev Ecol Syst 17:567–594
Weyers HS, Suberkropp K (1996) Fungal and bacterial production during the breakdown of yellow poplar leaves in two streams. J North Am Benthol Soc 15:408–420
Wood-Eggenschwiller S, Bärlocher F (1983) Aquatic hyphomycetes in sixteen streams in France, Germany and Switzerland. Trans Brit Mycol Soc 81:371–379
Zak DR, Holmes WE, White DC, Peacock AD, Tilman D (2003) Plant diversity, soil microbial communities and ecosystem function: are there any links? Ecology 84:2042–2050
Zar JH (1996) Biostatistical analysis, 3rd edn. Prentice Hall International, Englewood Cliffs, NJ
Acknowledgment
The present research was financially supported by the Office National des Forêts, the Conseil Général des Vosges and by the French ministry of ecology and sustainable development. Additional support was provided by the EU Commission (RivFunction: contact EVK1-CT-2001-00088).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Baudoin, J.M., Guérold, F., Felten, V. et al. Elevated Aluminium Concentration in Acidified Headwater Streams Lowers Aquatic Hyphomycete Diversity and Impairs Leaf-Litter Breakdown. Microb Ecol 56, 260–269 (2008). https://doi.org/10.1007/s00248-007-9344-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-007-9344-9