
Abstract
Environmental microbiology investigation was carried out in Jiaozhou Bay to determine the source and distribution of tetracycline-resistant bacteria and their resistance mechanisms. At least 25 species or the equivalent molecular phylogenetic taxa in 16 genera of resistant bacteria could be identified based on 16S ribosomal deoxyribonucleic acid sequence analysis. Enterobacteriaceae, Pseudomonadaceae, and Vibrionaceae constituted the majority of the typical resistant isolates. Indigenous estuarine and marine Halomonadaceae, Pseudoalteromonadaceae, Rhodobacteraceae, and Shewanellaceae bacteria also harbored tetracycline resistance. All the six resistance determinants screened, tet(A)–(E) and tet(G), could be detected, and the predominant genes were tet(A), tet(B), and tet(G). Both anthropogenic activity-related and indigenous estuarine or coastal bacteria might contribute to the tet gene reservoir, and resistant bacteria and their molecular determinants may serve as bioindicators of coastal environmental quality. Our work probably is the first identification of tet(E) in Proteus, tet(G) in Acinetobacter, tet(C) and tet(D) in Halomonas, tet(D) and tet(G) in Shewanella, and tet(B), tet(C), tet(E), and tet(G) in Roseobacter.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Extensive use and misuse of antibiotics in medication, veterinary, agriculture, and aquaculture have caused antibiotic-resistant bacteria to be widespread in the environment [29, 30]. Elevated number of resistant bacteria, especially clinical strains, could be found near civic or hospital sewage discharging locations [8, 22]. High incidences of resistant bacteria in response to antibiotic usage have also been reported in coastal mariculture areas [25]. Aquatic ecosystems have been proposed as reservoirs of antibiotic resistance [5], and resistant bacteria in coastal environments represent a serious biotic contamination and a means for the spread and evolution of resistance genes and their vectors [46]. Although all these aspects raised the concern about the potential threat of wide-spread antibiotic resistance to public health and mariculture industry, resistance mechanisms in natural marine environments were seldom studied [2, 33].
Tetracycline class antibiotics have been widely used for microbial disease treatment and prevention [9]. Oxytetracycline used to be a common medicine in China and is still being intensively and intensely used as a therapeutic drug in veterinary and mariculture. Among the 39 resistance determinants discovered so far [1, 36], genes encoding active efflux pumps, such as tet(A)–(E) and tet(G), occurred frequently among marine Gram-negative bacteria [27]. The tet genes isolated from marine environments might have the same origins as those from clinical strains [22, 34]. However, most of the studies focused on antibiotic-resistant pathogens. Appraisals of the source, abundance, diversity, distribution, dynamics, and mechanisms of bacterial resistance in natural marine environments are largely untackled globally.
Jiaozhou Bay, located on the western coast of the Yellow Sea, is a semi-enclosed waterbody with a total area of 367 km2 and an average water depth of 7 m on the eastern part of the Shandong Peninsula of China. Environmental quality of Jiaozhou Bay has deteriorated dramatically in the past three decades because of rapid development in agriculture, industry, urbanization, and mariculture in the surrounding areas. More than 30% of the bay area has been lost over the past 80 years because of coastal land usage, and more than 60% of the remaining water body has been polluted or contaminated to some extent. Red tides and species loss are serious environmental and ecological issues in Jiaozhou Bay [38]. Surrounding small rivers used to form the major sources of discharged sewage, and improperly processed hospital effluent and civic wastewater could be a source of antibiotic-resistant bacteria. Excessive usage of antibiotics, such as oxytetracycline, in mariculture might also stimulate the propagation of resistant bacteria in Jiaozhou Bay.
To gain an understanding of antibiotic resistance contamination in a typical coastal environment that has been dramatically influenced by human activity and to define the source, diversity, distribution, and dynamics of the corresponding resistance determinants, cultivation and molecular techniques were employed to determine the phylogenies of typical tetracycline-resistant bacteria and their resistance genes in Jiaozhou Bay.
Materials and Methods
Sample Collection and Processing
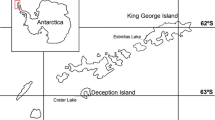
Surface seawater from ten stations (Fig. 1) of Jiaozhou Bay was collected aseptically on the 12th and 13th of September and the 11th and 12th of October of 2004. Water samples were kept on ice before being transferred to the laboratory, where samples for total microbial counts were fixed with formaldehyde (2% final concentration) and stored at 4°C in the dark, and samples for bacterial cultivations were processed immediately. A multifunction water quality monitoring system (model U-2001, Horiba, Japan) was used for in situ environmental factor measurements, except for seawater depth and diaphaneity. The environmental conditions seemed to be in normal ranges during the sampling season, except dramatic drops of seawater pH have been recorded in all the sampling stations in October (Table 1).

Maps show the Yellow Sea (left) and the sampling stations of Jiaozhou Bay (right). The locations of sewage processing plants are indicated with the asterisk symbols
Microbial Number Counts
For total microbial and cultivable bacteria counts, 4′,6-diamidino-2-phenylindole staining epifluorescence microscopy and 2,216 marine agar plating techniques were used as described previously [16, 17]. For cultivable tetracycline-resistant bacteria counts, a membrane filter technique was used to concentrate aquatic bacteria [37]. Water samples from each station were diluted into a tenfold series in sterilized artificial seawater, and 3-mL portions were filtered onto sterilized 0.22-μm-pore-size polycarbonate membrane filters in triplicates. The filters were placed onto tryptic soy agar (Difco formula) plates supplemented with 3% (w/v) NaCl and 10 μg/mL oxytetracycline (OTC10). OTC10-resistant colonies on the positively growing highest or the second highest dilution (if there were too few colonies on the highest) plates were further screened with 30 μg/mL oxytetracycline (OTC30) based on the method described for antibiotic susceptibility tests [27]. Only OTC30-resistant isolates were counted as tetracycline-resistant bacteria, which were further assayed for higher resistance to 100 μg/mL oxytetracycline (OTC100).
Phylogenetic Analysis of Typical Tetracycline-resistant Isolates
OTC100-resistant bacteria were randomly selected from each station, and some colonies were also picked based on their unique colony morphology and color features. As all of these isolates were originally screened from the positively growing highest or the second highest dilution OTC10 plates, they represented the predominant tetracycline-resistant bacteria in Jiaozhou Bay. Eventually, 89 isolates (about eight to ten isolates from each station) were selected for subsequent amplified ribosomal deoxyribonucleic acid (rDNA) restriction analysis (ARDRA) genotyping, 16S rDNA sequencing, and tet gene screening.
A simple boiling method was used for rapid bacterial genomic DNA extraction [18]. The primers 27F and 785R were used for bacterial 16S rDNA amplification as described previously [16, 17]. Polymerase chain reaction (PCR) amplicons were digested in separate reactions with MspI and TaqI (Fermentas, USA) according to the manufacturer’s instructions. Restriction fragments were resolved by electrophoresis on 4% (w/v) agarose gels and digitally photographed with an ImageMaster VDS imaging system (Pharmacia Biotech, USA). The band patterns were compared to identify unique sequence types. Bacteria with identical ARDRA pattern using both enzymes were assumed to be of the same genotype and phylogenetic affiliation in this study.
One isolate from each genotype was selected for 16S rDNA sequencing and phylogenetic determination. Partial 16S rDNA was PCR amplified to serve as the template, and primer 27F was used as the sequencing primer. The DNA fragments sequenced were usually about 600–700 bp long, covering at least the V1 to V3 hypervariable regions of the bacterial 16S rDNA.
Bioinformatic determination of sequence affiliations followed standard methods described previously [13]. Phylogenetic trees were constructed by using the DNADIST and NEIGHBOR programs within the PHYLIP package (version 3.6) [19].
Multiplex PCR Detection of Efflux tet Genes
The above selected 89 OTC100-resistant isolates were screened for the tet(A)–(E) and tet(G) genes via a multiplex PCR method developed by Jun et al. [27]. The original primers (SI_Table 1), experimental procedure, and PCR amplification condition were used, only with a minor modification for total DNA (including genomic and plasmid DNA) extraction [18]. Representative PCR products from each of the six tet gene types identified were sequenced to confirm their molecular identities as the typical efflux types of tetracycline resistance genes.
Nucleotide Sequence Accession Numbers
The 16S rDNA sequences determined have been deposited in the National Center for Biotechnology Information GenBank database under accession nos. DQ316101 to DQ316136 and the six representative tet gene sequences determined under accession nos. DQ316137 to DQ316142.
Results
Microbial Abundance and Distribution
During the sampling period, the aquatic microbial density ranged from 2.7 × 106 to 3.8 × 107 cells/mL, the cultivable bacterial abundance ranged from 1.9 × 102 to 5.2 × 105 cells/mL, and the OTC30-resistant bacteria ranged from 1 to 4.2 × 102 cells/mL in the surface seawater samples of Jiaozhou Bay (Table 2). In general, the OTC30-resistant bacteria accounted for less than 0.6% of the total cultivable bacteria. However, their spatial distribution was not homogeneous. The highest OTC30-resistant bacteria abundance occurred at station A5 of the northeastern part of Jiaozhou Bay, around the estuary of river Licun (Fig. 1).
Decrease of total microbial densities occurred in October compared to those in September (Table 2). However, the temporal changes of culturable and OTC30-resistant bacteria abundance did not have clear trends, and the percentages of resistant bacteria to culturable bacteria were not consistent for any sampling stations between the two different sampling periods. Currently, it is not evident about the relationship of the changing environment conditions, such as the dramatic drop of seawater temperature and pH (Table 1), with the temporal change of the microbial abundance in Jiaozhou Bay.
Phylogenetic Affiliations of the Typical Resistant Isolates
A total of 704 OTC30-resistant bacteria (402 from September and 302 from October) were isolated from surface seawater samples of Jiaozhou Bay, and 270 isolates (117 from September and 153 from October) showed resistance to at least 100 μg/mL oxytetracycline. A total of 89 typical resistant isolates (52 from September and 37 from October) were selected for further molecular analyses.
Based on ARDRA analyses, 32 unique bacterial genotypes could be identified for the 89 OTC100-resistant isolates. They were further analyzed by 16S rDNA sequence determination. All the representative 32 isolates had 97% or greater 16S rDNA sequence similarity to their best-match sequences retrieved from the GenBank database (SI_Table 2). The phylogenetic tree constructed further verified their phylogenetic affiliations (Fig. 2). The OTC100-resistant bacteria were quite diverse; at least 25 species or the equivalent molecular phylogenetic taxa in 16 genera could be identified. For the two strains that were closely related to Pseudomonas stutzeri, two genomovars could be distinguished from each other [39]. Most of the isolates belonged to the γ-Proteobacteria subdivision, and some might be potential human or marine animal pathogens, such as Acinetobacter, Citrobacter, Escherichia, Klebsiella, Proteus, Photobacterium, and Vibrio. The remaining isolates belonged to either the α-Proteobacteria subdivision or the phylum Bacteroidetes.

Phylogenetic tree of the 32 typical OTC100-resistant strains isolated from Jiaozhou Bay constructed based on partial 16S rDNA sequences in a 620-bp frame length using the neighbor-joining method with the Kimura two-parameter model for nucleotide change. The tree branch distances represent nucleotide substitution rate, and the scale bar represents the expected number of changes per homologous nucleotide position. Bootstrap values greater than 70% of 100 resamplings are shown near nodes as double asterisk, and those greater than 90% are shown as asterisk
Bacteria affiliated to Enterobacteriaceae contribute to 37.5% (12 strains) of the 32 sequenced OTC100-resistant isolates, including species closely related to Citrobacter freundii, Escherichia coli, Klebsiella oxytoca, Klebsiella pneumoniae, Kluyvera intermedia, Proteus mirabilis and Raoultella ornithinolytica. Most of these species occurred at stations A5, B2, C4, D6 and Y1 of September. The Acinetobacter strains occurred exclusively at station A5 of September. The other OTC100-resistant isolates may be indigenous estuarine or coastal marine bacteria, such as Halomonas, Photobacterium, Pseudoalteromonas, Pseudomonas, Psychrobacter, Roseobacter, Shewanella, and Vibrio.
Bacteria affiliated to Enterobacteriaceae, Pseudomonadaceae, and Vibrionaceae constituted the majority of the selected typical OTC100-resistant isolates. The combination of ARDRA and 16S rDNA sequence analyses indicated that 31.5% (28 out of 89) of the resistant isolates were affiliated to Enterobacteriaceae, including 15 isolates of Klebsiella, four isolates of Citrobacter, four isolates of Escherichia, two isolates of Kluyvera, two isolates of Raoultella, and one isolate of Proteus (SI_Table 2). Most of these bacteria (82.1%) were isolated from stations A5, Y1, and C4, which are located on the eastern coast of Jiaozhou Bay (Fig. 1). Pseudomonadaceae bacteria contributed to 21.3% of the 89 analyzed resistant isolates, including 16 Pseudomonas isolates and three Acinetobacter isolates. Vibrionaceae bacteria contributed to 19.1% of the 89 analyzed resistant isolates, including 16 Vibrio isolates and one Photobacterium isolate.
Diversity of Efflux tet Genes
All the six tet genes screened, tet(A)–(E) and tet(G), were detected. Of the 49 tet-positive isolates (tet gene carrying strains), 16 carried only tet(A), ten carried only tet(B), two carried only tet(C), two carried only tet(D), one carried only tet(E), 13 carried only tet(G), one carried both tet(B) and tet(C), one carried both tet(C) and tet(D), and three carried both tet(E) and tet(G) (SI_Fig. 1, SI_Table 2). The molecular mechanism encoding for tetracycline resistance of the remaining 40 strains could not be identified by the current multiplex PCR method. The predominant tetracycline resistance genes in Jiaozhou Bay during our sampling period were tet(A), tet(B), and tet(G). Most of the tet genes were found in bacteria isolated from stations A3, A5, B2, C1, C4, D6, and Y1, whereas quite a little or even none was detected in C3, D1, or D5, which are located near the mouth of Jiaozhou Bay (Table 3, Fig. 1). The resistance gene composition was highly dynamic; the same type of the screened tet genes was seldom shared by the same station of both sampling months (Table 3). The bacterial hosts of the detected tet genes were quite diverse, including both anthropogenic activity-related strains and indigenous estuarine or marine strains (Table 3, SI_Table 2).
The combination of ARDRA, 16S rDNA sequence, and tet gene analyses indicated that ten Klebsiella strains out of the 89 analyzed bacteria carried a tet gene. It is interesting to note that all the tet-positive K. oxytoca-related strains carried tet(B), and all the tet-positive K. pneumoniae-related strains carried tet(A), except for strain OO-A5-6, which carried tet(D) and was only remotely related to the other tet-positive K. pneumoniae strains found in Jiaozhou Bay (see Fig. 2). Of the other Enterobacteriaceae bacteria, the C. freundii-like isolates carried either tet(A) or tet(B), and the P. mirabilis-like isolate carried tet(E).
The Acinetobacter isolates mainly carried tet(B) or tet(G). Most of the Pseudomonas isolates carried tet(A) or tet(G) and occasionally tet(C). The Vibrio strains carried mainly tet(B) and occasionally tet(A) or tet(G). Five isolates carried two different types of tet genes simultaneously. They belonged to the genus Roseobacter or Halomonas; both are typical coastal marine bacteria.
The identities of the representative tet genes detected via the multiplex PCR method were further verified by DNA sequencing. One representative strain from each group of the bacterial isolates related to a single type of the six tet genes screened was randomly selected for tet gene sequencing (Table 4). All the representative sequences of the screened tet gene types had 99% similarity to their best-match known genes retrieved from the GenBank database, and they also had 99% or higher similarity to the best-match known efflux-type tetracycline resistance protein sequences after they were translated into conceptual amino acid sequences. The multiplex PCR method adopted [27] provides a rapid and reliable approach for typical tet gene detection and differentiation in the marine environment studied.
Discussion
Environmental bacteria may play an important role as reservoirs of antibiotic resistance, and resistance genes are exchanged by bacteria from different ecosystems [35]. A basic understanding of the source, abundance, diversity, distribution, and dynamics of antibiotic-resistant bacteria and their resistance determinants is necessary for effective prevention and control of antibiotic resistance and its dissemination. Modern molecular techniques have been proven to be suitable for studying resistant bacteria and their resistance mechanisms [2, 27]. The employment of molecular methods in the current project helps us uncover diverse tetracycline-resistant bacteria and their resistance genes. However, higher resistant bacteria abundance and diversity should be expected in Jiaozhou Bay if samples were collected from the middle or bottom of the seawater, as the surface seawater might have much reduced bacterial populations because of the effect of UV sunlight irradiation during our sampling period.
Diverse Antibiotic-resistant Bacteria from Coast Marine Environments
Two sources of OTC100-resistant bacteria could be identified in Jiaozhou Bay, the indigenous estuarine or coastal marine bacteria and the terrestrial bacteria related to anthropogenic activities. Enterobacteriaceae bacteria were detected mainly along the densely populated eastern coast of Jiaozhou Bay. Civic and hospital wastewater discharges via rivers or sewage processing plants on the seashore might be a major source of Enterobacteriaceae in the seawater. Most of the Enterobacteriaceae strains were isolated in September, suggesting a seasonal succession likely introduced by the change of environmental factors (Table 1) or the variation of the terrestrial bacterial source. Some bacteria might also enter the viable but nonculturable state with the drop of seawater temperature [10], contributing to the drop of culturable Enterobacteriaceae abundance.
The distribution of OTC100-resistant bacteria was quite heterogeneous. Stations on the eastern coast of Jiaozhou Bay, such as A5, C4, and Y1, were highly contaminated with Enterobacteriaceae. Station A5 is around the river mouth of Licun and the Licun River sewage processing plant (Fig. 1), which discharges about 50,000 tons of processed sewage into the bay daily. Station Y1 is located at the river mouths of Loushan and Moshui near the Hongdao aquacultural area. Station C4 is located near the river mouth of Haipo and the Haipo River sewage processing plant, which discharges about 80,000 tons of processed sewage into the bay daily. Our current study indicates that the spatial distribution of antibiotic resistant microbes might be closely related to geographical characteristics and anthropogenic activities and that sewage contamination might be a serious environmental problem of Jiaozhou Bay, at least in our sampling period.
The Bulletin of Marine Environmental Quality of Qingdao City of the year 2004 (http://www.coi.gov.cn/dfhygb/2004/qingdao/index.html) shows that the eastern coast of Jiaozhou Bay was heavily polluted by inorganic nitrogen, phosphate, heavy metal, oil, and other organic pollutants, indicating that anthropogenic activity has an important impact on the coastal environment. Although the seawater of Jiaozhou Bay is highly dynamic and water mixing happens frequently because of currents, tides, and other hydrological factors, confined distribution of resistant bacteria, especially the terrestrial strains, still could be identified along the eastern coast, indicating a strong terrestrial or anthropogenic impact.
Diverse Efflux tet Genes from Coastal Marine Environments
Molecular characterization of tet genes helped decode partially the tetracycline-resistance mechanisms in Jiaozhou Bay. Other or even novel mechanisms might also exist. All the tet(A)–tet(E) and tet(G) genes could be detected, and tet(A), tet(B), and tet(G) were the most predominant determinants in Jiaozhou Bay, in contrary to tet(E) that was found to be dominant in marine sediments of Norway and Denmark a decade ago [2]. The tet determinants could vary with sampling origin, suggesting ecosystem-specific reservoirs for certain antibiotic resistance [24].
Basically, two sources of tet genes could be identified, one from the terrestrial bacteria coming from anthropogenic activities of the surrounding environments and one from the indigenous estuarine or coastal marine bacteria (Table 3). Transfer of resistance determinants between different environmental compartments has been demonstrated [34], indicating that a common gene pool might be shared between environmental microorganisms and clinical strains. Terrestrial bacteria entering into seawater with antibiotic-resistant plasmids were proposed to be partially responsible for the prevalence of resistance genes in marine environments [4, 7]. The resistance gene flow between the indigenous environmental bacterial assemblage and the human activity-related bacterial assemblage in Jiaozhou Bay will be determined in the future.
Members of Enterobacteriaceae are usually found in the gastrointestinal tract of humans and animals, and certain strains are primary or opportunistic pathogens [31]. Tetracycline-resistant Enterobacteriaceae strains seem to be prevalent globally [21, 28]. Multidrug-resistant K. pneumoniae isolated from Nigeria harbored tet(A) [40]. Klebsiella strains isolated from Europe carried tet(A), tet(B), or tet(D) [20]. The tet(A) gene was also detected in most of the K. pneumoniae strains isolated from Jiaozhou Bay, indicating a possible global distribution of the common Klebsiella tetracycline-resistance determinants. Multidrug-resistant Citrobacter strains isolated from fish farms in southern Japan harbored tet(D) [22], whereas the Citrobacter strain isolated from Jiaozhou Bay carried tet(A) or tet(B). This difference seems to suggest regional or ecosystem-specific reservoirs of tetracycline resistance determinants for this genus. Our work probably is the first identification of tet(E) in Proteus.
Acinetobacter strains are usually commensal, but in the past few decades, they have emerged as important opportunistic nosocomial pathogens [44]. Most of the clinical strains isolated from Europe carried either tet(A) or tet(B) [23, 26], whereas the aquatic strains isolated from sewers receiving waste effluent from a hospital and a pharmaceutical plant in Denmark carried only tet(B) [23]. In Jiaozhou Bay, some of the Acinetobacter isolates harbored either tet(B) or tet(G). The tet(G) gene was probably the first identification in Acinetobacter, suggesting regional or ecosystem-specific reservoirs of this resistance determinant in Acinetobacter.
Marine vibrios have long been recognized as important reservoirs and vehicles of antibiotic resistance because of their importance as potential human and/or marine animal pathogens [41], their abundance and diversity in coastal waters [42], their ability to readily develop and acquire antibiotic resistance in response to selective pressure, and their ability to spread resistance by horizontal genetic material exchange [3]. The major vibrio tetracycline-resistance gene in Jiaozhou Bay was tet(B) and occasionally tet(A) or tet(G). The tet genes carried by vibrios isolated recently from a mariculture farm in northern China were tet(A), tet(B), and tet(D) [16, 17]. The tet gene carried on R plasmids from fish-pathogenic V. anguillarum isolated from 1973 to 1977 in Japan was tet(B), from 1980 to 1983 was tet(G), and from 1989 to 1991 was tet(D) [3]. The tet gene carried on R plasmids from fish-pathogenic Edwardsiella tarda in Korea was mainly tet(A) and tet(D), and tet(B) and tet(G) were occasionally detected on nonmobile nucleic acids [27]. The prevalence of similar tet genes in Korea and China at the present time to the originally isolated tet genes from Japan decades ago might aid in the speculation about the origin and spread of certain antibiotic resistance factors among the three neighboring countries [17], although local strains might also contribute to resistance gene transfer because of anthropogenic activities in coastal marine environments.
Besides Enterobacteriaceae, Pseudomonadaceae, and Vibrionaceae, many typical indigenous estuarine and costal marine bacteria also harbored tet genes in Jiaozhou Bay. It is probably the first identification of tet(B), tet(C), tet(E), and tet(G) in Roseobacter, tet(C) and tet(D) in Halomonas, and tet(D) and tet(G) in Shewanella (SI_Table 2). The Roseobacter clade is one of the major marine groups of coastal bacterioplankton communities [6, 14, 15, 45]. Roseobacter isolates occurred abundantly and widely in Jiaozhou Bay (SI_Table 2). All the Roseobacter isolates are tet positive, and more than half of them actually harbored two different tet genes simultaneously. This unique phenomenon further emphasizes the ecological and environmental importance of this clade of marine bacteria.
Deterioration of coastal environmental quality is becoming a global issue because of pollution and eutrophication. Oxygen-starved areas in bays and coastal waters have been expanding since the 1960s, and the number of known dead zones around the world has doubled since 1990 [43]. The occurrence and persistence of diverse antibiotic-resistant bacteria and their resistance genes makes the situation even worse and more complicated. The study of tetracycline-resistant bacteria isolated from marine sediments indicated that the resistant species were different between the polluted and the unpolluted sites [2]. Our current study also indicates the possibility that antibiotic-resistant bacteria and their molecular resistance determinants may serve as bioindicators of coastal environmental quality consistently influenced by anthropogenic activities.
Epidemic diseases caused by antibiotic-resistant bacteria have been reported all over the world. More than 10,000 deaths caused by cholera were reported to the World Health Organization in 1998, and antibiotic-resistant Vibrio cholera strains have been reported frequently [11, 12]. Epidemics of pathogens in coastal marine environments may spread in large scale and at extremely rapid speed because of their potential for long-term survival outside the host and the lack of barriers to dispersal [32]. Antibiotic-resistant microbes may pose a particular threat in coastal areas. Uncovering the genetic mechanisms behind resistance gene transfer will be an important research orientation for public health and coastal environment management.
References
Agerso Y, Guardabassi L (2005) Identification of Tet 39, a novel class of tetracycline resistance determinant in Acinetobacter spp. of environmental and clinical origin. J Antimicrob Chemother 55:566–569
Andersen SR, Sandaa R-A (1994) Distribution of tetracycline resistance determinants among gram-negative bacteria isolated from polluted and unpolluted marine sediments. Appl Environ Microbiol 60:908–912
Aoki T (2000) Transferable drug resistance plasmids in fish-pathogenic bacteria. In: Arthur JR, Lavilla-Pitogo CR, Subasinghe RP (eds) Use of chemicals in aquaculture in Asia. SEAFDEC Aquaculture DepartmentTigbauan, Iloilo, Philippines, pp 31–33
Baya AM, Brayton PR, Brown VL, Grime DJ, Russek-Choen E, Colwell RR (1986) Coincident plasmids and antimicrobial resistance in marine bacteria isolated from polluted and unpolluted Atlantic Ocean samples. Appl Environ Microbiol 51:1285–1292
Biyela PT, Lin J, Bezuidenhout CC (2004) The role of aquatic ecosystems as reservoirs of antibiotic resistant bacteria and antibiotic resistance genes. Water Sci Technol 50:45–50
Buchan A, Gonzalez JM, Moran MA (2005) Overview of the marine Roseobacter lineage. Appl Environ Microbiol 71:5665–5677
Chandrasekaran S, Venkatesh B, Lalithakumari D (1998) Transfer and expression of a multiple antibiotic resistance plasmid in marine bacteria. Curr Microbiol 37:347–351
Choi S, Chu W, Brown J, Becker SJ, Harwood VJ, Jiang SC (2003) Application of enterococci antibiotic resistance patterns for contamination source identification at Huntington Beach, California. Mar Pollut Bull 46:748–755
Chopra I, Roberts M (2001) Tetracycline antibiotics: mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol Mol Biol Rev 65:232–260
Colwell RR, Grimes DJ (2000) Nonculturable microorganisms in the environment. ASM, Washington, DC
Dalsgaard A, Forslund A, Sandvang D, Arntzen L, Keddy K (2001) Vibrio cholerae O1 outbreak isolates in Mozambique and South Africa in 1998 are multiple-drug resistant, contain the SXT element and the aadA2 gene located on class 1 integrons. J Antimicrob Chemother 48:827–838
Dalsgaard A, Forslund A, Tam NV, Vinh DX, Cam PD (1999) Cholera in Vietnam: changes in genotypes and emergence of class I integrons containing aminoglycoside resistance gene cassettes in Vibrio cholerae O1 strains isolated from 1979 to 1996. J Clin Microbiol 37:734–741
Dang HY, Lovell CR (2000) Bacterial primary colonization and early succession on surfaces in marine waters as determined by amplified rRNA gene restriction analysis and sequence analysis of 16S rRNA genes. Appl Environ Microbiol 66:467–475
Dang HY, Lovell CR (2002) Numerical dominance and phylotype diversity of marine Rhodobacter species during early colonization of submerged surfaces in coastal marine waters as determined by 16S ribosomal DNA sequence analysis and fluorescence in situ hybridization. Appl Environ Microbiol 68:496–504
Dang HY, Lovell CR (2002) Seasonal dynamics of particle-associated and free-living marine Proteobacteria in a salt marsh tidal creek as determined using fluorescence in situ hybridization. Environ Microbiol 4:287–295
Dang HY, Song LS, Chen MN, Chang YQ (2006) Concurrence of cat and tet genes in multiple antibiotic resistant bacteria isolated from a sea cucumber and sea urchin mariculture farm in China. Microb Ecol 52:634–643
Dang HY, Zhang XX, Song LS, Chang YQ, Yang GP (2006) Molecular characterizations of oxytetracycline resistant bacteria and their resistance genes in mariculture waters of China. Mar Pollut Bull 52:1494–1503
De Medici D, Croci L, Delibato E, Di Pasquale S, Filetici E, Toti L (2003) Evaluation of DNA extraction methods for use in combination with SYBR green I real-time PCR to detect Salmonella enterica serotype enteritidis in poultry. Appl Environ Microbiol 69:3456–3461
Felsenstein J (1989) PHYLIP-Phylogeny Inference Package (Version 3.2). Cladistics 5:164–166
Fluit AC, Florijn A, Verhoef J, Milatovic D (2005) Presence of tetracycline resistance determinants and susceptibility to tigecycline and minocycline. Antimicrob Agents Chemother 49:1636–1638
Fritsche TR, Strabala PA, Sader HS, Dowzicky MJ, Jones RN (2005) Activity of tigecycline tested against a global collection of Enterobacteriaceae, including tetracycline-resistant isolates. Diagn Microbiol Infect Dis 52:209–213
Furushita M, Shiba T, Maeda T, Yahata M, Kaneoka A, Takahashi Y, Torii K, Hasegawa T, Ohta M (2003) Similarity of tetracycline resistance genes isolated from fish farm bacteria to those from clinical isolates. Appl Environ Microbiol 69:5336–5342
Guardabassi L, Dijkshoorn L, Collard JM, Olsen JE, Dalsgaard A (2000) Distribution and in-vitro transfer of tetracycline resistance determinants in clinical and aquatic Acinetobacter strains. J Med Microbiol 49:929–936
Guillaume G, Verbrugge D, Chasseur-Libotte M, Moens W, Collard J (2000) PCR typing of tetracycline resistance determinants (Tet A–E) in Salmonella enterica serotype Hadar and in the microbial community of activated sludges from hospital and urban wastewater treatment facilities in Belgium. FEMS Microbiol Ecol 32:77–85
Herwig RP, Gray JP, Weston DP (1997) Antibacterial resistant bacteria in surficial sediments near salmon net-cage farms in Puget Sound, Washington. Aquaculture 149:263–283
Huys G, Cnockaert M, Vaneechoutte M, Woodford N, Nemec A, Dijkshoorn L, Swings J (2005) Distribution of tetracycline resistance genes in genotypically related and unrelated multiresistant Acinetobacter baumannii strains from different European hospitals. Res Microbiol 156:348–355
Jun LJ, Jeong JB, Huh MD, Chung JK, Choi DL, Lee CH, Jeong HD (2004) Detection of tetracycline-resistance determinants by multiplex polymerase chain reaction in Edwardsiella tarda isolated from fish farms in Korea. Aquaculture 240:89–100
Kim SH, Wei CI, Tzou YM, An H (2005) Multidrug-resistant Klebsiella pneumoniae isolated from farm environments and retail products in Oklahoma. J Food Prot 68:2022–2029
Kummere K (2004) Resistance in the environment. J Antimicrob Chemother 54:311–320
Levy SB, Marshall B (2004) Antibacterial resistance worldwide: causes, challenges and responses. Nat Med 10:S122–S129
Linton AH, Hinton MH (1988) Enterobacteriaceae associated with animals in health and disease. Soc Appl Bacteriol Symp Ser 17:71S–85S
McCallum H, Harvell D, Dobson A (2003) Rates of spread of marine pathogens. Ecol Lett 6:1062–1067
Mudryk ZJ (2005) Occurrence and distribution antibiotic resistance of heterotrophic bacteria isolated from a marine beach. Mar Pollut Bull 50:80–86
Rhodes G, Huys G, Swings J, McGann P, Hiney M, Smith P, Pickup RW (2000) Distribution of oxytetracycline resistance plasmids between aeromonads in hospital and aquaculture environments: implication of Tn 1721 in dissemination of the tetracycline resistance determinant Tet A. Appl Environ Microbiol 66:3883–3890
Roberts MC (1996) Tetracycline resistance determinants: mechanisms of action, regulation of expression, genetic mobility, and distribution. FEMS Microbiol Rev 19:1–24
Roberts MC (2005) Update on acquired tetracycline resistance genes. FEMS Microbiol Lett 245:195–203
Rompre A, Servais P, Baudart J, de-Roubin MR, Laurent P (2002) Detection and enumeration of coliforms in drinking water: current methods and emerging approaches. J Microbiol Methods 49:31–54
Shen ZL (2001) Historical changes in nutrient structure and its influences on phytoplankton composition in Jiaozhou Bay. Estuar Coast Shelf Sci 52:211–224
Sikorski J, Mohle M, Wackernagel W (2002) Identification of complex composition, strong strain diversity and directional selection in local Pseudomonas stutzeri populations from marine sediment and soils. Environ Microbiol 4:465–476
Soge OO, Queenan AM, Ojo KK, Adeniyi BA, Roberts MC (2006) CTX-M-15 extended-spectrum b-lactamase from Nigerian Klebsiella pneumoniae. J Antimicrob Chemother 57:24–30
Thompson FL, Iida T, Swings J (2004) Biodiversity of vibrios. Microbiol Mol Biol Rev 68:3403–3431
Thompson JR, Pacocha S, Pharino C, Klepac-Ceraj V, Hunt DE, Benoit J, Sarma-Rupavtarm R, Distel DL, Polz MF (2005) Genotypic diversity within a natural coastal bacterioplankton population. Science 307:1311–1313
UNEP (2004) GEO Year Book 2003. Global Environment Outlook (GEO)/United Nations Environment Programme (UNEP). United NationsNew York, NY 9280724150
Van Looveren M, Goossens H, the ARPAC Steering Group (2004) Antimicrobial resistance of Acinetobacter spp. in Europe. Clin Microbiol Infect 10:684–704
Wagner-Dobler I, Biebl H (2006) Environmental biology of the marine Roseobacter lineage. Annu Rev Microbiol 60:255–280
Young HK (1993) Antimicrobial resistance spread in aquatic environments. J Antimicrob Chemother 31:627–635
Acknowledgments
This work was financially supported by the Pilot Projects of Knowledge Innovation Project of Chinese Academy of Sciences grants KZCX2-YW-211-03, KZCX3-SW-214, KZCX3-SW-223, and KZCX3-SW-233, and the National Natural Science Foundation of China grants 40476058 and 40576069.
Author information
Authors and Affiliations
Corresponding authors
Electronic supplementary material
Tables for the information of the multiplex PCR primers adopted from Jun et al. [27] (SI_Table 1) and for the phylogenetic affiliations and the tet genes detected of the 89 selected OTC100-resistant bacteria (SI_Table 2) and a figure of the electrophoretic analyses of the tet genes screened with the multiplex PCR method (SI_Fig. 1) are available via the Internet of the journal Microbial Ecology website.
Below is the link to the electronic supplementary material.
SI_Table 1
Multiplex PCR primers for tet genes detection (from Jun et al. [27] (DOC 26.5 kb)
SI_Table 2
Bacterial 16S rDNA sequence affiliations and the efflux tet genes detected (DOC 50 kb)
SI_Fig. 1
Electrophoretic analyses of the tet genes amplified with the multiplex PCR method developed by Jun et al. [27] from the selected 89 OTC100-resistant isolates. Strains that did not harbor any of the tet genes screened were not shown. Top 10: E. coli strain TOP10 was used to serve as negative control for multiplex PCR reactions, as this strain did not harbor tetracycline resistance. N: negative control without template DNA. M. Molecular weight standard DL2000 (2000, 1000, 750, 500, 250 and 100 bp respectively) (DOC 119 kb).
Rights and permissions
About this article
Cite this article
Dang, H., Ren, J., Song, L. et al. Diverse Tetracycline Resistant Bacteria and Resistance Genes from Coastal Waters of Jiaozhou Bay. Microb Ecol 55, 237–246 (2008). https://doi.org/10.1007/s00248-007-9271-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-007-9271-9