
Abstract
It has been suggested that oxidative stress products play an important role in the etiology of epilepsy. We investigated the effects of selenium (Se) administration on topiramate (TPM)- and pentylentetrazol (PTZ)-induced brain toxicity in rats. Forty male Wistar rats were divided into five equal groups. The first and second groups were used as the control and PTZ groups, respectively. TPM, 50 mg, and Se, 0.3 mg, were administered to rats constituting the third and fourth groups, respectively, for 7 days. The combination of 50 mg TPM and Se was given to animals in the fifth group for 7 days. At the end of 7 days all groups except the first received a single dose of PTZ. Brain cortex samples were taken at 3 h of PTZ administration. PTZ resulted in a significant increase in brain cortex and microsomal lipid peroxidation (LP) levels, number of spikes, and epileptiform discharges on the EEG, although brain cortex vitamin E, brain cortex and microsomal reduced glutathione (GSH), and microsomal calcium (Ca) levels, Ca2+-ATPase activities, and latency to first spike on the EEG were decreased by PTZ. LP, GSH, vitamin E, and Ca levels and Ca2+-ATPase activities were increased by both Se and TPM, although vitamin A and C concentrations were increased by Se only. There were no effects of TPM and Se on brain cortex and microsomal glutathione peroxidase, brain cortex nitric oxide, or β-carotene levels. In conclusion, TPM and selenium caused protective effects on PTZ-induced brain injury by inhibiting free radical production, regulating calcium-dependent processes, and supporting the antioxidant redox system.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Oxidative stress is defined as an imbalance between higher cellular levels and reactive oxygen species (ROS), e.g., superoxide radical, hydrogen peroxide, and nitric oxide (NO) (Yatin et al. 2000; Nazıroğlu 2007a, b), and cellular antioxidant defense (Ilhan et al. 2005). Generation of ROS is ubiquitous since ROS are generated during aerobic metabolism, i.e., mitochondrial oxidations and other monoamine oxidants. To scavenge ROS, various defense systems exist in the brain. Selenium (Se) is an essential dietary trace element which plays an important role in a number of biological processes (Whanger 2001). Glutathione peroxidase (GSH-Px), a Se-containing enzyme, is responsible for the reduction of hydro and organic peroxides in the presence of reduced glutathione (GSH) (Nazıroğlu 2003). GSH is a hydroxyl radical and singlet oxygen scavenger and it participates in a wide range of cellular functions (Whanger 2001). Vitamin E, α-tocopherol, is the most important antioxidant in the lipid phase of cells (Nazıroğlu 2007a). Vitamin C, as well as being a free radical scavenger, also transforms vitamin E to its active form (Arrigoni and De Tullio 2002). Vitamin A, retinol, serves as a prohormone for retinoids and is involved with signal transduction at cytoplasmic and membrane sites (Czernichow and Hersberg 2001).
If ROS are not controlled by enzymatic and nonenzymatic antioxidants, they can cause oxidative injury, i.e., peroxidation of cell membrane phospholipids, proteins (receptor and enzymes), and DNA. The brain is extremely susceptible to oxidative damage induced by these ROS, because it generates very high levels of ROS due to its very high aerobic metabolism and blood perfusion and it has relatively poor enzymatic antioxidant defense (Nazıroğlu 2007a). The brain contains polyunsaturated fatty acids (PUFAs) which can readily be peroxidized (Özmen et al. 2007). Lipid peroxidation (LP) causes injury to cell and intracellular membranes and may lead to cell destruction and, subsequently, cell death (Yatin et al. 2000; Eren et al. 2007). Brains are protected by antioxidants from peroxidative damage (Nazıroğlu et al. 2008).
Epilepsy is one of the most common neurological disorders. Oxidative stress due to generation of mitochondrial ROS is strongly implicated in seizures in the disorder. The oxidative stress product and NO are known to be a cause of epileptic seizures (Patel 2004; Ilhan et al. 2005). Oxidative stress has been shown in several rodent models of experimental epilepsy such as the amygdala kindling model (Frantseva et al. 2000) and the pentylentetrazol (PTZ) model (Uma Devi et al. 2006; Akbas et al. 2005; Armağan et al. 2008). PTZ is characterized by an increased susceptibility to seizures following injection of initially subconvulsion doses of PTZ culminating in generalized tonic-clonic seizures. PTZ is a selective blocker of chloride ionosphere complex to the γ-aminobutyric acidA (GABAA) receptor, and after repeated or single-dose administration it can lead to decreased GABAergic function (White et al. 2007). Increased activity of glutamatergic systems induced by status epilepticus causes energy impairment and enhanced formation of ROS as described for kainite-evoked seizures. Balance of certain minerals is crucial for a healthy nervous system and neuronal susceptibility to excitability. Several reports have suggested that body Se levels play a vital role in development of seizure conditions (see review by Schweizer et al. 2004a).
Topiramate (TPM) is new antiepileptic drug which inhibits voltage-gated sodium and calcium channels, blocks glutamate AMPA/kainite receptors, and enhances GABAA receptor-mediated chloride (White et al. 2007). An increasing body of evidence indicates that TPM possesses not only antiepileptic but also neuroprotective properties due to its multiple mechanisms of action (Cardile et al. 2001). However, it was reported that it may cause adverse effects such as blood toxicity (Eraković et al. 2003; Akbas et al. 2005), although there is no information on the effects of oxidative stress or role of Se in PTZ or TPM in oxidative stress-induced toxicity in brain. Reports on effects of antiepileptic drugs except for TPM related to oxidative stress and NO are present in some tissues and plasma, and they are also controversial. For example, Cardile et al. (2001) and Pavone and Cardile (2003) reported that in astrocytes TPM increased oxidative stress as malondialdehyde (MDA), although Kubera et al. (2004) recently reported that TPM plays an antioxidant role and it attenuated LP levels in the pyriform cortex of rats. Exposure of mitochondria to high cytosolic free Ca2+ was shown to increase formation of ROS (Dykens 1994). It has been reported that TPM (Weiergräber et al. 2006) in PTZ-induced epileptic cells of mice and vitamin E in hippocampal slice culture (Kovács et al. 2002) treatments modulated cytosolic Ca2+ levels by regulation of voltage-gated calcium channels (VGCCs). Modulation of VGCCs in brain by means of treatment with TPM with or without Se may cause a decrease in mitochondrial ROS production.
It has not been examined whether TPM with or without Se modifies alterations in the antioxidant enzyme system or LP and NO levels in the brain cortex and microsomes in rat. Hence, we aimed to evaluate whether there is a protective effect of TPM and Se on oxidative stress, enzymatic antioxidants and NO, plasma membrane Ca2+-ATPase (PMCA), or calcium (Ca) values in PTZ-induced brain injury in rats.
Materials and Methods
Animals
Forty male Wistar albino rats weighing 200 ± 20 g were used for experimental procedures. Rats were allowed 1 week to acclimate to the surroundings before beginning any experimentation. Animals were housed in individual plastic cages with bedding. Standard rat food and tap water were available ad libitum for the duration of the experiments unless otherwise noted. The temperature was maintained at 22 ± 2°C. A 12/12-h light/dark cycle was maintained, with lights on at 06.00, unless otherwise noted. The experimental protocol was approved by the ethical committee of the Medical Faculty of Suleyman Demirel University. Animals were maintained and used in accordance with the Animal Welfare Act and the Guide for the Care and Use of Laboratory Animals prepared by Suleyman Demirel University.
Experimental Design
TPM at 50 mg/kg body weight (BW)/day has been commonly used for treatment of epilepsy in humans. Hence we wanted to study the effect of a 50-mg dose of TPM on the investigated values. Forty animals were randomly divided into five groups as follows.
-
Group I: Control group (n = 8). Placebo was administered to rats in this group.
-
Group II: PTZ (60 mg/kg BW) was administered orally to rats (n = 8) for induction of epilepsy.
-
Group III: TPM (50 mg/kg/day) was given orally (via gastric gavage) to animals (n = 8) for 7 consecutive days.
-
Group IV: Se (0.3 mg/kg over 1 day) was intraperitoneally (i.p.) administered to rats (n = 8) for 6 consecutive days before PTZ administration.
-
Group V: TPM (50 mg/kg/day) was orallyadministered (via gastric gavage) to animals (n = 8) for 7 consecutive days before PTZ administration. Rats also received i.p. Se (0.3 mg/kg BW over 1 day; 1-day interval) three times before PTZ administration.
Epilepsy was induced in groups II, III, IV, and V by administration of PTZ (60 mg/kg). After 3 h of PTZ administration all rats were sacrificed and brain samples were taken. Due to the absorption difference between intraperitoneal and intragastric administrations, Se was administered to the animals for 6 days (total of three doses), but TPM was given for 7 days (total of seven doses).
Seizure intensity was evaluated using the following modified scale (Eraković et al. 2003; Ilhan et al. 2005).
-
0:
no response
-
1:
ear and facial twitching
-
2:
convulsive waves axially through body
-
3:
myoclonic body jerks
-
4:
generalized clonic convulsion, with a turn onto the side
-
5:
generalized convulsions, tonic extension episode, and status epilepticus
Anesthesia and Preparation of Brain Samples
Rats were anesthetized with a cocktail of ketamine hydrochloride (50 mg/kg) and xylazine (5 mg/kg) administered i.p. before sacrifice of each rat and removal of cortex brain samples used for EEG records. The brain was also taken as follows: the cortex was dissected out after the brain was split in the midsagittal plane. Following removal of the cortex the brain was dissected from the total brain as described in our previous study (Eren et al. 2007).
Cortex brain tissues were washed twice with cold saline solution, placed into glass bottles, labeled, and stored in a deep freeze (−30°C) until processing (maximum, 10 h). After weighing, half of the cortex samples were placed on ice, cut into small pieces using scissors, and homogenized (2 min at 5000 rpm) in 5 vol (1:5, w/v) of ice-cold Tris-HCl buffer (50 mM, pH 7.4) using a glass Teflon homogenizer (Caliskan Cam Teknik, Ankara, Turkey). All procedures were performed on ice. The homogenate was used for determination of LP and antioxidant levels. The remaining cortex samples were used for Ca2+-ATPase assay and isolation of microsomes by ultracentrifugation.
After the addition of butylhydroxytoluol (4 μl/ml), brain homogenate and microsome samples were used for immediate LP levels and enzyme activities. Antioxidant vitamin analyses were performed within 3 months.
Isolation of Brain Microsomes
Tissues were cleaned, minced, and then homogenized in 6 vol of freshly prepared buffer A containing 0.3 M sucrose, 10 mM Hepes-HCl, pH 7.4, and 2 mM dithiothreitol. The material was homogenized with a glass-Teflon homogenizer. The homogenate was centrifuged (MS 80; Sanyo Inc.) at 85,000g (Sorvall; Teknolab A.Ş, Ankara, Turkey) for 75 min. The supernatant was discarded and the pellet was resuspended in the original volume of buffer A containing 0.6 M KCl using four strokes of the pestle and centrifuged again at 85,000g for 75 min. The pellet was resuspended in the original volume of buffer A. After centrifugation at 85,000g for 75 min, the pellet was suspended in buffer A using four strokes of the pestle at a protein concentration of 2–7 mg/ml. The procedures took 10–12 h for eight samples. Samples were frozen and stored at −30°C until assay. The isolation procedure was carried out at +4°C (Doğru Pekiner et al. 2005).
Measurement of Microsomal Ca2+-ATPase Activity
Ca2+-ATPase activity was measured spectrophotometrically by the method of Niggli et al. (1981). Assay medium contained 120 mM KCl, 60 mM Hepes, pH 7 (at 37°C), 1 mM MgCl2, 0.5 mM K2-ATP, 0.2 mM NADH, 0.5 mM PEPA, 1 IU/L pyruvate kinase, 1 IU/L lactate dehydrogenase, and 500 mM EGTA. After preincubation of the assay medium (total volume, 1 ml) for 4 min at 37°C, 50 mg of the microsome preparation was added to the medium. After 2 min the reaction was started by the addition of 600 mM CaCl2. ATPase activity as oxidation of NADH was followed by continuously measuring the absorbance at 340 nm.
Based on the extinction coefficient for NADH, ε = 6.2 × 106 M−1, the amount of NADH oxidized is equivalent to the amount of ATP hydrolyzed. Values are expressed as International Units per milligram protein.
Determination of Total Brain Calcium Levels
Microsomal Ca levels were analyzed with a plasma optic emission atomic absorption spectrophotometer (ICP-OES; Optima 4300 DV; Perkin Elmer Life and Analytical Sciences, Inc., Waltham, MA, USA) by following the wet ashing procedure with nitric acid. Ca values were measured at 422.7 nm in the atomic absorption spectrophotometer.
Lipid Peroxidation Level Determinations
LP levels in the brain homogenate and microsomal samples were measured using the thiobarbituric acid reaction by the method of Placer et al. (1966). LP levels in the brain and microsomes are expressed as micromoles per gram protein. Although the method is not specific for LP, measurement of the thiobarbituric acid reaction is an easy and reliable method, which is used as an indicator of LP and ROS activity in biological samples.
Brain Cortex Nitric Oxide Assay
The NO content in the brain cortex was measured in an ELISA (ELx808 Absorbance Microplate Reader; BioTec Instrument Inc., USA) using commercial kits (nitrate/nitrite colorimetric assay kit; Catalog No. 780001; Cayman Chemical Inc., USA).
Brain Cortex and Microsomal Reduced Glutathione, Glutathione Peroxidase, and Protein Assay
The GSH content of the brain homogenate and microsomes was measured at 412 nm using the method of Sedlak and Lindsay (1968) as described in our previous studies (Eren et al. 2007). GSH-Px activities of red blood cells were measured spectrophotometrically at 37ºC and 412 nm according to Lawrence and Burk (1976). The protein content in the brain cortex and microsomes was measured by the method of Lowry et al. (1951) with bovine serum albumin as the standard.
Brain Cortex β-Carotene and Vitamin A, C, and E Analyses
Vitamins A (retinol) and E (-tocopherol) were determined in brain cortex samples by a modification of the method described by Desai (1984) and Suzuki and Katoh (1990). Vitamin A and E concentrations were measured in a spectrophotometer (Shimadzu 1700) at 325 and 535 nm, respectively. Calibration was performed using standard solutions of all-trans retinol and α-tocopherol in hexane. Quantification of vitamin C (ascorbic acid) in brain cortex samples was performed according to the method of Jagota and Dani (1982). Absorbance of the samples was measured spectrophotometrically at 760 nm.
Electroencephalographic Recording
Before sacrifice of the animals, EEG records were taken by a polygraph (ADI Instruments, Australia). In a stereotaxic frame, two monopolar stainless-steel electrodes were placed bilaterally over the frontal and parietal cortex surface EEG recording. Electrodes were connected to a female connector anchored to the skull with dental acrylate. A neutral electrode was connected to the tail or ear of each animal.
EEG recording and behavioral observations were carried out in the Faraday cage. Animals were connected with a male microconnector to the headbox of the recording apparatus with a polygraph system and XA-400 quad-channel differential amplifier (ML 870; ADI Instruments). EEG biopotentials were recorded using the ADI Instruments X-chart 5 program (time constant, 1 s; low-pass filter, 50 Hz; high-pass filter, 1 Hz; range, 200 μV). Baseline EEG records were performed 15 min before TPM and Se treatment with or without PTZ administration. After administration of TPM with or without Se, records were again taken before PTZ administration. The last EEG records were taken for 10 min from groups II, III, IV, and V after PTZ administration. Alpha waves are responsible for 70% of brain biopotentials. For calculation of a certain spike, the records were filtered (high cutoff frequency, 12 Hz; low cutoff frequency, 8 Hz).
A spike was defined as a high-amplitude (twice baseline), sharply contoured wave form with a duration of 20–70 ms. Latency to first spike and total number of spikes were calculated 1 min after PTZ administration. These parameters were chosen according to Kharatishvili et al. (2007).
Pentylentetrazol Test
To detect any enhanced seizure susceptibility, numbers of spikes were considered after single PTZ administration and the results were compared with those for controls. Spikes occurred within 15 min of PTZ administration. Forms of spikes in PTZ-administered groups were significantly different from normal EEG records. To determine whether the seizure threshold was reduced after TPM and Se administration, seizures were compared in the TPM + Se group versus the PTZ group.
Behavioral signs of seizure activity, which typically occurred sequentially after PTZ administration, were (1) myoclonic twitch—sudden, involuntary jerking of the whole body; (2) clonic movements of forelimbs with preservation of the righting reflex; and (3) generalized clonic seizure with loss of righting reflex, which corresponds to the observations by Kharatishvili et al. (2007).
In the EEG records, a single dose of PTZ induced electrographic epileptiform discharges (brief bursts of high-amplitude slow waves, spikes, and/or spike-and-wave episodes) in all animals in both treatment and PTZ groups. A brief (≤1-s) spike wave and a high-amplitude delta burst correspond to myoclonic twitches, while ictal epileptiform discharges of >5-s duration correspond to clonic seizures. The general evaluation of EEG records after PTZ injection was as follows: (1) brief bursts of high-amplitude slow waves, (2) isolated spike waves or bursts of several spikes lasting ≤1 s, and (3) electrographic seizures. The general evaluation of EEG records in TPM- and Se-administered groups after PTZ injection was as follows: (a) decreased brief bursts of high-amplitude slow waves, (b) a decreased count of isolated spike waves or bursts of several spikes lasting ≤1 s, and (c) decreased electrographic seizures.
Statistical Analyses
All results are expressed as means ± SD. To determine the effect of treatment, data were analyzed using ANOVA. p-values <0.05 were regarded as significant. Significant values were assessed by LSD test. Data were analyzed using the SPSS statistical program (version 9.05; SPSS Inc., Chicago, IL, USA).
Results
Results of Lipid Peroxidation and Nitric Oxide Results
Mean brain cortex and microsomal LP levels in the five groups are reported in Tables 1 and 2. The results show that LP levels in the brain cortex and microsomes in the PTZ group were significantly higher than in the control group. Hence, PTZ caused increased oxidative stress of the brain. However, administration of TPM with or without Se caused decreased LP levels in the brain cortex. The brain cortex and microsomal LP levels in the TPM, Se, and TPM + Se groups were significantly lower than in the PTZ group. Mean brain cortex NO levels in the five groups are reported in Table 1, and NO levels were not affected by PTZ, TPM, and Se administration.
Glutathione Peroxidase Activities and Reduced Glutathione Levels
Mean brain cortex and microsomal GSH-Px activities in the five groups are listed in Tables 1 and 2. The results showed that brain cortex and microsomal GSH-Px activities were insignificantly lower in the PTZ group compared to the control, although their activities were increased by TPM and Se administrations.
Mean brain cortex and microsomal GSH levels in the five groups are listed in Tables 1 and 2. GSH levels in microsomes were significantly decreased in PTZ groups versus the control, although brain cortex GSH levels were insignificantly decreased. The decreased brain cortex and microsomal GSH levels were improved by either 50-mg TPM supplementation or TPM + Se administration. On the other hand, brain cortex and microsomal GSH levels in the TPM, Se, and TPM + Se groups were significantly higher than in the PTZ group.
Results of Antioxidant Vitamin Concentrations
Mean brain cortex β-carotene concentrations in the five groups are reported in Table 1, and they were not affected by PTZ, TPM, and Se administrations.
Mean brain cortex vitamin A, C, and E concentrations in the five groups are also listed in Tables 1 and 2. The results show that the brain cortex vitamin A and C concentrations were insignificantly lower in the PTZ group compared to the control, although vitamin E concentrations were significantly lower in the PTZ group compared to the control. The decreased vitamin A, C, and E concentrations were not affected by TPM supplementation. However, the decreased vitamin A, C, and E concentrations were increased by Se with and without TPM administration. Vitamin A, vitamin C, and vitamin E levels in the TPM, Se, and TPM + Se groups were significantly higher than in the PTZ group.
Results on Microsomal Calcium Levels and Plasma Membrane Ca2+-ATPase Activities
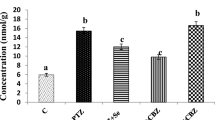
Mean Ca levels and PMCA activities in microsomes of the five groups are reported in Table 2 and Fig. 1, respectively. Ca levels were significantly higher in the PTZ group than in the control, although Ca levels in the TPM, Se, and TPM + Se groups were significantly lower than in the PTZ groups.

Effects of topiramate (TPM) and selenium (Se) on microsomal plasma membrane Ca2+-ATPase activity in pentylentetrazol (PTZ)-induced brain toxicity in rats. Mean ± SD; n = 8
PMCA activities were significantly lower in the PTZ group than in the control, although PMCA activities in the TPM, Se, and TPM + Se groups were significantly higher than in the PTZ groups.
Results of EEG Records
Mean EEG records and alpha waves (filtered records) are shown in Figs. 2 and 3, respectively. Mean spikes in the brain of the five groups are shown in Fig. 4. The total number of spikes during 1-min follow-up was higher in the PTZ group compared to controls (mean, 32 versus 1), although the total numbers of spikes in the TPM (mean, 27.6), Se (mean, 19.1), and TPM + Se (mean, 14.25) groups were significantly lower compared to the PTZ group. Also, the total number of epileptiform discharges was higher in PTZ group compared to controls (mean, 35 versus 3), although the total number of epileptiform discharges in the TPM (mean, 29), Se (mean, 19), and TPM + Se (mean, 15) groups were significantly lower compared to the PTZ group.

Examples of EEG records in the five groups (control, PTZ, PTZ + TPM, PTZ + Se, and PTZ + TPM + Se). EEG biopotentials were recorded using the ADI Instruments X-chart 5 program (time constant, 1 s; low-pass filter, 50 Hz; high-pass filter, 1 Hz; range, 200 μV). Baseline EEG records were obtained 15 min before TPM and Se treatment or with PTZ administration. After administration of TPM with or without Se, EEG records were again taken before PTZ administration. The last records were taken for 10 min from the TPM, Se, and TPM + Se groups after PTZ administration. n = 8

Examples of EEG records in the control, PTZ, PTZ + TPM, PTZ + Se, and PTZ + TPM + Se groups according to alpha waves. Alpha waves are responsible for 70% of brain biopotentials. The mean records were filtered (high cutoff frequency; 12 Hz, low cutoff frequency, 8 Hz) to obtain alpha records. n = 8

PTZ test results for number of spikes and number of epileptiform discharges in alpha waves in the control, PTZ, PTZ + TPM, PTZ + Se, and PTZ + TPM + Se groups. Parameters used to assess lowered threshold in the PTZ test. Rats with PTZ had increased numbers of spikes and increased numbers of epileptiform discharges, indicating hyperexcitability. In animals receiving TPM, Se, and Se + TPM before PTZ administration, the seizure duration, number of spikes, and number of epileptiform discharges decreased, which indicated that Se and TPM had protective effects on hyperexcitability. Mean ± SD; n = 8. *p < 0.001 versus control group. a p < 0.05, b p < 0.01, and c p < 0.001 versus PTZ group
The latency to first spike in the brain of the five groups is shown in Fig. 5. The latency to first spike during 1-s follow-up was shorter in the PTZ group than in the control (mean, 109 versus 55.01). The latency to first spike was recovered in the TPM, Se, and TPM + Se groups. The latencies to first spike in the TPM (mean, 90.6), Se (mean, 94) and TPM + Se groups (mean, 99) were significantly longer compared to the PTZ group.

PTZ test results for latency to first spike in the control, PTZ, PTZ + TPM, PTZ + Se, and PTZ + TPM + Se groups. Parameters used to assess lowered threshold in the PTZ test. Rats with PTZ had shorter latencies to first spike, indicating hyperexcitability. In animals receiving TPM, Se, and Se + TPM before PTZ administration, the latency to first spike increased. Thus, Se and TPM had protective effects on hyperexcitability. Mean ± SD; n = 8. *p < 0.001 versus control group. c p < 0.001 versus PTZ group
According to the results, rats administered PTZ had a shorter latency to first spike, an increased number of spikes, and an increased number of epileptiform discharges, indicating hyperexcitability. In animals receiving TPM, Se, and Se + TPM before PTZ administration, the seizure duration was lower. Thus, Se and TPM had protective effects on hyperexcitability.
Discussion
We found that brain cortex and microsomal LP and microsomal Ca levels were increased by PTZ administration, although brain and microsomal GSH, brain vitamin E levels, and PMCA activities were decreased. Hence, PTZ administrations in the rats were characterized by increased LP and decreased PMCA, GSH, and vitamin E antioxidant values. Administration of TPM and Se caused a decrease in microsomal Ca and brain cortex and microsomal LP levels, although GSH levels, vitamin A, E, and C concentrations, and PMCA activities increased. A limited number of in vivo or in vitro studies in tissues besides the brain cortex and microsomes of experimental animals have been reported regarding the effects of TPM on the antioxidant enzymatic system and LP and PMCA values (Cardile et al. 2001; Pavone and Cardile 2003; Kubera et al. 2004). To the best of our knowledge, the current study is the first to compare TPM and Se with particular reference to its effects on oxidative stress and the antioxidant redox system in PTZ-induced brain injury in rats.
The current study indicates that PTZ administration at a convulsive dose of 60 mg/kg produced a significant increase in brain cortex and microsomal LP levels. Our results are in accordance with previous reports of increased LP in brain, erythrocytes, and liver during epileptic seizures (Kubera et al. 2004; Akbas et al. 2005; Deniz Onay et al. 2008). On the other hand, the current study is the first report on brain microsomal LP in epileptic rats. PTZ may trigger a variety of biochemical process including the activation of membrane phospholipases, proteases, and nucleases (Deniz Onay et al. 2008). Marked alterations in membrane phospholipid metabolism result in the liberation of LP and ROS. Hence, involvement of LP as MDA in PTZ-induced epilepsy contributed to the activation of membrane phospholipases. In the current study, GSH and vitamin E in the brain cortex and microsomes were decreased by PTZ-induced epilepsy. If GSH levels decreases, superoxide radical production may increase and, finally, lead to oxidative stress and LP (Nazıroğlu 2007a, b).
Inactivation of ROS can be achieved with antioxidant vitamins (Nazıroğlu 2007b). Vitamin E, α-tocopherol, is the most important antioxidant in the lipid phase of cells. Vitamin E acts to protect cells against the effects of free radicals, which are potentially damaging by-products of the body’s metabolism (Yatin et al. 2000). Vitamin C, as well as being a free radical scavenger, also transforms vitamin E to its active form (Arrigoni and De Tullio 2002). Levels of antioxidants in the brain are considerably low. Therefore, low antioxidant levels and a high PUFA content result in limited antioxidant defense in the brain. Vitamin A, C, and E concentrations in the brain cortex were decreased in the PTZ group, although their concentrations in the brain cortex were increased in the TPM and Se treatment groups. The increased concentrations of antioxidant vitamins could be due to their depletion or inhibition as a result of the increased production of free radicals. The increase in brain cortex vitamin A, C, and E levels in animals during TPM and Se treatments has been attributed to the inhibition of free radicals and LP (Kubera et al. 2004; Ilhan et al. 2005).
Anticonvulsant drug therapy can alter intracellular GSH levels. GSH plays an important role in protecting cells against oxidative damage as a nonenzymatic antioxidant and the most abundant nonprotein thiol source in cells (Akbas et al. 2005; Deniz Onay et al. 2008). Large doses of these convulsant drugs tend to deplete hepatic stores of GSH and leave hepatocytes much more vulnerable to oxidative stress (Ono et al. 2000). Ono et al. (2000) demonstrated that some anticonvulsants caused a significant decrease in plasma total GSH levels, suggesting that oxidative stress can occur during anticonvulsant treatment. In the current study, brain cortex and microsomeal GSH concentrations decreased in epileptic rats not receiving any epileptic drug therapy, although GSH levels were increased by TPM administration. Hence, the study supports the notion that oxidative stress does not occur with anticonvulsant drugs such as TPM, but seizures themselves might directly induce oxidative stress.
The role of Se in the detoxifying enzyme, GSH-Px, has been known; this awful poison has a positive biological function in various aspects of human health (Whanger 2001). We found that there were no significant changes in GSH-Px activity in brain cortex and microsomal PTZ-induced epileptic rats. There are conflicting reports on the effects of antiepileptic drugs except TPM on GSH-Px activity in humans and animals. For example, Kürekçi et al. (1995) reported increased GSH-Px activity in human blood caused by anticonvulsant valproate supplementation, although other, more recent papers (Verroti et al. 2002; Ashafi et al. 2007) indicated that no changes in GSH-Px activity in blood of epileptic patients were caused by drug supplementation.
It has also been suggested that antiepileptic drugs except TPM are associated with detrimental effects on brain antioxidant defense systems (Maertens et al. 1995). In addition, Ca and Ca-dependent processes have been hypothesized to be involved in the induction of epilepsy. Recently, Isokawa (2005) reported that persistent swelling in the epileptic hippocampus was reduced by the endoplasmic reticulum Ca2+-ATPase inhibitor, thapsigargin, and suggested that it might involve a mechanism of Ca2+ release from internal stores. LP and Ca levels were decreased by TPM with or without Se, although PMCA activities were increased by TPM and Se administrations. These findings suggest that TPM may interfere with production of ROS due to an inhibitory effect on VGCCs. This observation is in agreement with the findings of other studies that TPM was effective against kainite-induced LP production in the pyriform cortex (Kubera et al. 2004) and other regions of the brain (Ilhan et al. 2005), ischemia-induced neuronal damage (Lee et al. 2000), and PTZ-induced kidney damage (Armağan et al. 2008).
A large number of studies have linked seizure-induced cell damage to excitotoxic mechanisms (for review see Kovács et al. 2002). Convulsions can result in augmented glutamate release, leading to Ca2+ uptake through N-methyl-D-aspartate and VGCCs. In fact during convulsions induced by different means and in different models, extracellular Ca2+ decreases while the cytosolic Ca2+ concentration increases (White et al. 2007) Mitochondria were reported to accumulate Ca2+ provided that cytosolic Ca2+ rises to >400 nm or provided that mitochondrial uptake dominates mitochondrial Ca2+ extrusion (Colegrove et al. 2000), thereby leading to depolarization of mitochondrial membranes (Kovács et al. 2002). Uptake of Ca2+ into mitochondria stimulates the tricarboxylate cycle, resulting in augmented reduction of pyridine nucleotides, which may be one of the mechanisms of the coupling of neuronal and metabolic activity (Duchen 2000). On the other hand, exposure of mitochondria to high cytosolic free Ca2+ was shown to increase formation of ROS (Weiergräber et al. 2006). It has been reported that TPM (Weiergräber et al. 2006) in PTZ-induced epileptic cells of mice and vitamin E in hippocampal slice culture (Kovács et al. 2002) treatments modulated cytosolic Ca2+ levels by regulation of VGCCs. In the current study brain cortex and microsomal LP levels were lower in the TPM and Se groups than in the PTZ group. Modulation of VGCC in brain cells by means of treatment with TPM and Se might cause a decrease in mitochondrial ROS production.
In the current study, the level of NO did not change in PTZ and treatment groups. The role of NO in the pathophysiology of seizures remains unclear and debatable. Experiments performed with nonselective NOS inhibitors yielded conflicting results, and accordingly, the authors proposed both pro- and anticonvulsant effectives of NO (Kirkby et al. 1996). Similar to the present NO results, Ilhan et al. (2005) reported that NO levels in brains of PTZ-induced epileptic mice were not increased by antioxidant Nigella sativa oil supplementation. Consequently, more research is needed to reach a final conclusion on the possible contribution of NO to the pathophysiology of seizures in epilepsy.
In the current study, the numbers of spikes and epileptiform discharges on the EEG increased in the PTZ group, although the latency to first spike on the EEG was decreased by PTZ. The changes were reversed by Se administration. A link between seizures and Se has also been suggested by observations of patients on total parenteral nutrition who were at risk to develop seizures until Se was incorporated into their formulations (Brown et al. 1986), and our results are supported by the observation. Similarly, Schweizer et al. (2004b) reported that mice deficient in selenoprotein P exhibit a disturbed Se distribution and reduced activities of other selenoenzymes and display defects in growth and motor coordination, although they have normalized selenoenzyme activities, and rescued the phenotype of mutant mice by supplementing their nursing mothers with Se (sodium selenite). In a model of Fe2+-induced epileptic discharges, application of Se normalized the EEG records and reduced tissue damage as assessed by a histological method (Willmore and Rubin 1981). Öztaş et al. (2001) have shown that in PTZ-induced seizures the breakdown of the blood-brain barrier is attenuated by dietary Se administration.
In conclusion, our results in brain for the PTZ group are consistent with a generalized antioxidant abnormality in different tissues of epileptic animals and humans. However, TPM with Se supplementation has a protective effect on oxidative stress and the antioxidant redox system in the brain cortex and microsomes. TPM and selenium have beneficial effects on enzymatic antioxidant systems including regulation of EEG records, GSH, antioxidant vitamins, PMCA activities, and LP levels in the brain. Hence, the use of Se with or without TPM could be a potential approach to arresting or inhibiting the genesis of seizures caused by excitotoxic agents.
Abbreviations
- GSH:
-
Glutathione
- GSH-Px:
-
Glutathione peroxidase
- LP:
-
Lipid peroxidation
- MDA:
-
Malondialdehyde
- NO:
-
Nitric oxide
- PMCA:
-
Plasma membrane Ca2+-ATPase
- PTZ:
-
Pentylentetrazol
- ROS:
-
Reactive oxygen species
- SOD:
-
Superoxide dismutase
- TPM:
-
Topiramate
- VGCC:
-
Voltage gated calcium channels
References
Akbas SH, Yegin A, Ozben T (2005) Effect of pentylenetetrazol-induced epileptic seizure on the antioxidant enzyme activities, glutathione and lipid peroxidation levels in rat erythrocytes and liver tissues. Clin Biochem 38:1009–1014
Armağan A, Kutluhan S, Yılmaz M, Yılmaz N, Bülbül M, Vural H, Soyupek S, Nazıroğlu M (2008) Topiramate and vitamin E modulates antioxidant enzymes, nitric oxide and lipid peroxidation in pentylentetrazol-induced nephrotoxicity in rats. Basic Clin Pharm Toxicol 103:166–170
Arrigoni O, De Tullio MC (2002) Ascorbic acid: much more than just an antioxidant. Biochim Biophys Acta 1569:1–9
Ashrafi MR, Shams S, Nouri M, Mohseni M, Shabanian R, Yekaninejad MS, Chegini N, Khodadad A, Safaralizadeh R (2007) A probable causative factor for an old problem: selenium and glutathione peroxidase appear to play important roles in epilepsy pathogenesis. Epilepsia 48:1750–1755
Brown MR, Cohen HJ, Lyons JM, Curtis TW, Thunberg B, Cochran WJ, Klish WJ (1986) Proximal muscle weakness and selenium deficiency associated with long term parenteral nutrition. Am J Clin Nutr 43:549–554
Cardile V, Pavone A, Renis M, Maci T, Perciavalle V (2001) Effects of gabapentin and topiramate in primary rat astrocyte cultures. Neuroreport 12:1705–1708
Colegrove SL, Albrecht MA, Friel DD (2000) Dissection of mitochondrial Ca2+ uptake and release fluxes in situ after depolarization-evoked [Ca2+]i elevations in sympathetic neurons. J Gen Physiol 115:351–370
Czernichow S, Hersberg S (2001) Interventional studies concerning the role of antioxidant vitamins in cardiovascular disease: a review. J Nutr Health Aging 5:188–195
Deniz Onay B, Tasdemir E, Tümer C, Bilgin HM, Atmaca M (2008) Dose dependent effects of ghrelin on pentylenetetrazole-induced oxidative stress in a rat seizure model. Peptides 29:448–455
Desai ID (1984) Vitamin E analysis methods for animal tissues. Methods Enzymol 105:138–147
Doğru Pekiner B, Daş Evcimen N, Nebioğlu S (2005) Diabetes-induced decrease in rat brain microsomal Ca2+-ATPase activity. Cell Biochem Funct 23:239–243
Duchen MR (2000) Mitochondria and calcium: from cell signalling to cell death. J Physiol 529:57–68
Dykens JA (1994) Isolated cerebral and cerebellar mitochondria produce free radicals when exposed to elevated Ca2+ and Na+: implications for neurodegeneration. J Neurochem 63:584–591
Eraković V, Zupan G, Varljen J, Simonić A (2003) Pentylenetetrazol-induced seizures and kindling: changes in free fatty acids, superoxide dismutase, and glutathione peroxidase activity. Neurochem Int 42:173–178
Eren I, Naziroglu M, Demirdaş A (2007) Protective effects of lamotrigine, aripirazole and escitalopram on depression- induced oxidative stress in rat brain. Neurochem Res 32:1188–1195
Frantseva MV, Perez Velazquez JL, Tsoraklidis G, Mendonca AJ, Adamchik Y, Mills LR, Carlen PL, Burnham MW (2000) Oxidative stress is involved in seizure-induced neurodegeneration in the kindling model of epilepsy. Neuroscience 97:431–435
Ilhan A, Gurel A, Armutcu F, Kamisli S, Iraz M (2005) Antiepileptogenic and antioxidant effects of Nigella sativa oil against pentylenetetrazol-induced kindling in mice. Neuropharmacology 49:456–464
Isokawa M (2005) N-Methyl-D-aspartic acid-induced and Ca-dependent neuronal swelling and its retardation by brain-derived neurotrophic factor in the epileptic hippocampus. Neuroscience 131:801–812
Jagota SK, Dani HM (1982) A new colorimetric technique for the estimation of vitamin C using Folin phenol reagent. Anal Biochem 127:178–182
Kharatishvili I, Immonen R, Gröhn O, Pitkänen A (2007) Quantitative diffusion MRI of hippocampus as a surrogate marker for post-traumatic epileptogenesis. Brain 130(Pt 12):3155–3168
Kirkby RD, Carroll DM, Grossman AB, Subramaniam S (1996) Factors determining proconvulsant and anticonvulsant effects of inhibitors of nitric oxide synthase in rodents. Epilepsy Res 24:91–100
Kovács R, Schuchmann S, Gabriel S, Kann O, Kardos J, Heinemann U (2002) Free radical-mediated cell damage after experimental status epilepticus in hippocampal slice cultures. J Neurophysiol 88:2909–2918
Kubera M, Budziszewska B, Jaworska-Feil L, Basta-Kaim A, Leśkiewicz M, Tetich M, Maes M, Kenis G, Marciniak A, Czuczwar SJ, Jagła G, Nowak W, Lasoń W (2004) Effect of topiramate on the kainate-induced status epilepticus, lipid peroxidation and immunoreactivity of rats. Pol J Pharmacol 56:553–561
Kürekçi AE, Alpay F, Tanindi S, Gökçay E, Ozcan O, Akin R, Işimer A, Sayal A (1995) Plasma trace element, plasma glutathione peroxidase, and superoxide dismutase levels in epileptic children receiving antiepileptic drug therapy. Epilepsia 366:600–604
Lawrence RA, Burk RF (1976) Glutathione peroxidase activity in selenium-deficient rat liver. Biochem Biophys Res Commun 71:952–958
Lee SR, Kim SP, Kim JE (2000) Protective effect of topiramate against hippocampal neuronal damage after global ischemia in the gerbils. Neurosci Lett 281:183–186
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the Folin-phenol reagent. J Biol Chem 193:265–275
Maertens P, Dyken P, Graf W, Pippenger C, Chronister R, Shah A (1995) Free radical, anticonvulsants, and the neuronal ceroid-lipofuscinoses. Am J Med Genet 57:225–228
Nazıroğlu M (2003) Enhanced testicular antioxidant capacity in streptozotocin-induced diabetic rats: protective role of vitamins C and E and selenium. Biol Trace Elem Res 94:61–72
Nazıroğlu M (2007a) Molecular mechanisms of vitamin E on intracellular signaling pathways in brain. In: Goth L (ed) Reactive oxygen species and diseases. Research Signpost Press, Kerala, India, pp 239–256
Nazıroğlu M (2007b) New molecular mechanisms on the activation of TRPM2 channels by oxidative stress and ADP-ribose. Neurochem Res 32:1990–2001
Nazıroğlu M, Uğuz AC, Gokçimen A, Bülbül M, Karatopuk DU, Türker Y, Cerçi C (2008) Tenoxicam modulates antioxidant redox system and lipid peroxidation in rat brain. Neurochem Res 33:1832–1837
Niggli V, Adunyah ES, Penniston JT, Carafoli E (1981) Purified (Ca2+-Mg2+)-ATPase of the erythrocyte membrane. Reconstitution and effect of calmodulin and phospholipids. J Biol Chem 256:395–401
Ono H, Sakamato A, Sakura N (2000) Plasma total glutathione concentrations in epileptic patients taking anticonvulsants. Clin Chim Acta 298:135–143
Özmen I, Nazıroğlu M, Alicı HA, Sahin F, Cengiz M, Eren İ (2007) Spinal morphine administration reduces the fatty acid contents in spinal cord and brain in rabbits due to oxidative stress. Neurochem Res 32:19–25
Öztaş B, Kiliç S, Dural E, Ispir T (2001) Influence of antioxidants on the blood-brain barrier permeability during epileptic seizures. J Neurosci Res 66:674–678
Patel M (2004) Mitochondrial dysfunction and oxidative stress: cause and consequence of epileptic seizures. Free Radic Biol Med 37:1951–1962
Pavone A, Cardile V (2003) An in vitro study of new antiepileptic drugs and astrocytes. Epilepsia 44(Suppl 10):34–39
Placer ZA, Cushman L, Johnson BC (1966) Estimation of products of lipid peroxidation (malonyl dialdehyde) in biological fluids. Anal Biochem 16:359–364
Schweizer U, Bräuer AU, Köhrle J, Nitsch R, Savaskan NE (2004a) Selenium and brain function: a poorly recognized liaison. Brain Res Brain Res Rev 45:164–178
Schweizer U, Michaelis M, Köhrle J, Schomburg L (2004b) Efficient selenium transfer from mother to offspring in selenoprotein-P-deficient mice enables dose-dependent rescue of phenotypes associated with selenium deficiency. Biochem J 378(Pt 1):21–26
Sedlak J, Lindsay RHC (1968) Estimation of total, protein bound and non-protein sulfhydryl groups in tissue with Ellmann’s reagent. Anal Biochem 25:192–205
Suzuki J, Katoh N (1990) A simple and cheap method for measuring vitamin A in cattle using only a spectrophotometer. Jpn J Vet Sci 52:1282–1284
Uma Devi P, Pillai KK, Vohora D (2006) Modulation of pentylenetetrazole-induced seizures and oxidative stress parameters by sodium valproate in the absence and presence of N-acetylcysteine. Fund Clin Pharmacol 20:247–253
Verrotti A, Basciani F, Trotta D, Pomilio MP, Morgese G, Chiarelli F (2002) Serum copper, zinc, selenium, glutathione peroxidase and superoxide dismutase levels in epileptic children before and after 1 year of sodium valproate and carbamazepine therapy. Epilepsy Res 48:71–75
Weiergräber M, Henry M, Krieger A, Kamp M, Radhakrishnan K, Hescheler J, Schneider T (2006) Altered seizure susceptibility in mice lacking the Ca(v)2.3 E-type Ca2+ channel. Epilepsia 47:839–850
Whanger PD (2001) Selenium and the brain: a review. Nutr Neurosci 4:81–97
White HS, Smith MD, Wilcox KS (2007) Mechanisms of action of antiepileptic drugs. Int Rev Neurobiol 81:85–110
Willmore LJ, Rubin JJ (1981) Antiperoxidant pretreatment and iron-induced epileptiform discharges in the rat: EEG and histopathologic studies. Neurology 31:63–69
Yatin SM, Varadarajan S, Butterfield DA (2000) Vitamin E prevents Alzheimer’s amyloid beta-peptide (1–42)-induced neuronal protein oxidation and reactive oxygen species production. J Alzheimers Dis 2:123–131
Acknowledgments
This study was supported by The Scientific and Technological Research Council of Turkey (TÜBİTAK; SBAG-HD-228). The suthors wish to thank Ph.D. students Ömer Çelik and A Cihangir Uğuz, Department of Biophysics, Suleyman Demirel University, for help with analyses.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Nazıroğlu, M., Kutluhan, S. & Yılmaz, M. Selenium and Topiramate Modulates Brain Microsomal Oxidative Stress Values, Ca2+-ATPase Activity, and EEG Records in Pentylentetrazol-Induced Seizures in Rats. J Membrane Biol 225, 39–49 (2008). https://doi.org/10.1007/s00232-008-9132-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00232-008-9132-6