
Abstract
The alien coral Tubastraea spp. has invaded Atlantic coral reefs since 1940s, but their effects on native coral assemblages are poorly understood. In this study the effects on coral assemblage structure and tissue mortality in native competitors by alien coral were investigated. We compared native coral cover in zones with and without T. tagusensis and evaluated the mortality of native coral species in natural encounters with T. tagusensis. Additionally, we evaluated the effects of contact with this invader on two native corals, Siderastrea stellata and Montastraea cavernosa, using a manipulative experiment. Multivariate analyses detected significant differences between coral assemblages in invaded and non-invaded zones. In the invaded zone, we observed greater cover of T. tagusensis on reef walls (34.9 ± 4.2 %) than on reef tops (18.5 ± 4.1 %). Madracis decactis and Mussismilia hispida were significantly less abundant in invaded zone than in non-invaded zones at the reef walls. Manipulative experiments showed a significant increase in tissue mortality of S. stellata (3.2 ± 1.4 cm2) after 60 days of contact with T. tagusensis and no effects on M. cavernosa after 90 days of contact. In natural encounters, 52 and 0.1 % of the S. stellata and M. cavernosa colonies, respectively, showed tissue mortality. These results indicate that competitive interactions with native competitors are important to understand alien coral establishment on coral reef. Our study documents the negative effects of the alien coral on coral reef assemblages and reinforces the urgent need for monitoring and management actions to control the expansion of this invader on Brazilian reefs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Biological invasion is a primary threat to global biodiversity and ecosystem function (Ruiz et al. 1997; Molnar et al. 2008; Seebens et al. 2013). Invasive species have had extensive effects on marine systems worldwide (Carlton and Geller 1993), and a large number of studies have demonstrated the negative impacts of invasive species on marine benthic assemblages (Hollebone and Hay 2007; Caralt and Cebrian 2013). However, coral reefs, one of the most biodiverse and productive marine systems in the world (e.g., Birkeland 1977; Connell 1978), have been the subject of relatively few rigorous studies evaluating the potential consequences of invasions on coral assemblages (Coles and Eldredge 2002; Fernández and Cortés 2005; Ávila and Carballo 2009; Pérez-Estrada et al. 2013). Coral reef systems are highly dependent on hermatypic corals which build complex three-dimensional physical structures (Jones et al. 1994; Graham and Nash 2012), creating a large variety of microhabitats and leading to a high diversity and abundance of organisms (Crowder and Cooper 1982; Coni et al. 2013; Leal et al. 2013; Graham 2014; Rogers et al. 2014; Leal et al. 2015). Thus, understanding how invasions affect these ecosystems is crucial for the well-being of the coral reefs and the organisms that depend of them.
The ahermatypic coral Tubastraea (Scleractinia, Dendrophylliidae), commonly known as cup coral, is a native genus from the Indo-Pacific region likely introduced by ships and/or oil platforms in the Atlantic Ocean in the 1940s (Cairns 2000; Castro and Pires 2001; Fenner 2001; Fenner and Banks 2004). Three species of Tubastraea genus were reported in the Atlantic (T. micranthus, T. coccinea and T. tagusensis). Tubastraea spp. is a successful invader due to its high fecundity (Glynn et al. 2008; De Paula et al. 2014), rapid growth (Wellington and Trench 1985), allelochemical defenses (Lages et al. 2010a, b) and competitive aggressiveness (Wellington and Trench 1985; Dos Santos et al. 2013). However, there are no reports of the ecological impacts of this invader on native coral species in coral reefs (Fenner and Banks 2004). Recent biological invasions by the species T. coccinea and T. tagusensis are threatening coral reefs along the northeast Brazilian coast in the southwestern Atlantic (Sampaio et al. 2012). These species were first reported on the subtropical southwest coast of Brazil (De Paula and Creed 2004) on rocky shores, where they can change the structure of benthic assemblages (Lages et al. 2011). However, there is an urgent need to assess the potential effects of these alien species on Brazilian corals, especially on the northeast coast, the region with the richest hermatypic coral fauna in the southwestern Atlantic (Leão et al. 2003). The orange cup coral represents a serious threat to the local marine biodiversity (Creed 2006) because Brazilian reefs are relatively poor in coral species but have high endemism (Castro and Pires 2001; Leão and Kikuchi 2001; Neves et al. 2006; Nunes et al. 2008). Direct interactions of T. tagusensis and T. coccinea with a Brazilian endemic coral (Mussismilia hispida) result in the tissue necrosis of native coral, but the effect is not reciprocal (Creed 2006; Dos Santos et al. 2013). These evidences suggest that the alien coral Tubastraea spp. can reduce or exclude the native coral species by space competition (Dos Santos et al. 2013).
Competition for space is an important process on coral reefs determining patterns of distribution, abundance and species diversity in benthic communities (Connell et al. 2004). Competitive processes regulate reef structure, since the space on hard substratum is limiting resource for the settlement, growth and reproduction of reef-building corals (Lang and Chornesky 1989; Connell et al. 2004; Chadwick and Morrow 2011). Competition among corals can occur (1) indirectly by overtopping, when growth by one colony (e.g., branching, tabular or foliaceous forms) overshadows another or (2) directly, when tissues of different colonies come into contact, resulting in necrosis of one or both competitors (Connell 1973). Direct interactions among corals are frequent in coral communities, especially those dominated by massive corals (e.g., Leão et al. 2003). These interactions may involve several mechanisms including, sweeper tentacles (Hidaka and Yamazato 1984), sweeper polyps (Peach and Hoegh-Guldberg 1999), mesenterial filaments (Lang 1973) and the secretion of mucus (Chadwick 1988). The competitive outcomes are complex and can lead to fitness reduction of the subordinate coral, cessation of growth along the margin of damaged and/or even competitive reversal. In the cases, the most aggressive coral species may impact the structure of reef communities (Dai 1990).
Dos Santos et al. (2013) showed that tissue necrosis caused by the alien coral species T. tagusensis and T. coccinea on the native coral M. hispida occurred mainly due to physical mechanisms in direct encounters. Thus, effects of direct encounters between Tubastraea spp. and native corals should be considered to understand biological invasions on Brazilian coral reefs. In this study, we evaluate whether T. tagusensis negatively affected the hermatypic coral assemblage in Brazil. To achieve this, we tested whether (1) sites with a high cover of T. tagusensis would have different structure of native coral assemblages compared with non-invaded sites and (2) whether direct encounter with T. tagusensis would cause tissue damage on native coral species. Therefore, we combined mensurative and manipulative experiments to provide an important baseline that will serve as a starting point for future monitoring and management actions.
Materials and methods
Study area
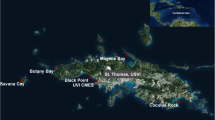
The study was conducted at Cascos Reef (13°07′46″S, 38°38′31″W) in the outer part of Todos os Santos Bay (TSB), east coast of Brazil (Fig. 1). Cascos Reef is the first coral reef site where T. tagusensis was documented in the southwestern Atlantic (Sampaio et al. 2012). This reef, at depths of approximately 20 m, consists of reef patches 11–13 m in height and 1–100 m in length (Fig. 1). Two main habitats are clearly distinguished on Cascos Reef: the reef top (horizontal surfaces; 11–13 m deep) and the walls (vertical surfaces; 12–20 m deep).

Location map of coral reefs in Todos os Santos Bay, Brazil (a): (b) Cascos Reef, with sampling zones (I = invaded zone; NI1 and NI2 = non-invaded zones) and (c) reef assemblages of the invaded zone
Eight species of native hermatypic coral occur on Cascos Reef, the scleractinians Montastraea cavernosa, Madracis decactis, Phyllangia americana, M. hispida, Mussismilia leptophylla, Siderastrea stellata, and Meandrina braziliensis (the last four species are endemic to Brazilian reefs: e.g., Leão et al. 2003; Neves et al. 2006; Nunes et al. 2008; Budd et al. 2012) and the hydrozoan fire coral Millepora alcicornis. There is only one zone on Cascos Reef where the population of T. tagusensis is aggregated over approximately 800 m2 (Fig. 1b, “Invaded Zone”). This population is dense, with patches measuring approximately 4 m2. T. coccinea was not found in field samples in the study area, and therefore, only T. tagusensis was used in our study.
Assemblage structure in invaded and non-invaded zones
We compared the structure of coral assemblages in the invaded zone (hereafter indicated as I) with that in non-invaded zones (NI1 and NI2, hereafter denoted NIs; Fig. 1b) to evaluate whether the presence of T. tagusensis affected the native coral assemblage structure. We selected the non-invaded zones based on their similarity in size (800 m2) and environmental conditions (i.e., wave exposure and depth) to the invaded zone. The non-invaded zones, where T. tagusensis did not occur, were 20 m apart. We sampled two distinct habitats, reef walls and reef tops, within each zone because of the marked differences in abiotic conditions (e.g., light incidence and substrate inclination, Francini-Filho et al. 2013).
We used an asymmetrical design to compare the coral cover between I and NIs. This design, with two non-invaded (“control”) locations (NIs), produces better estimates of natural variability than the use of a single“control” area, which was appropriate for this situation in which there was only a single impacted location (I) that could not be replicated (see Underwood 1992; Glasby 1997; Terlizzi et al. 2005a, b). Therefore, our design had three factors: Treatment (I and NIs, fixed and orthogonal), Zone (I, NI1 and NI2 random, nested in Treatment) and Habitat (Wall and Top fixed, nested in Zone).
During the summer of 2012/2013 (December–March), we characterized the coral assemblages using photo-quadrats (0.25 m2) of the reef tops and walls. We haphazardly took 80 photographs per zone (I, NI1 and NI2), 40 in each habitat (top and wall). We estimated the percentage of coral cover per species through 30 randomly distributed points per photo-quadrat (2400 points per zone, 1200 per habitat) using the Coral Point Count with Excel Extensions Software (CPCe) (http://www.nova.edu/ocean/cpce/) (Kohler and Gill 2006).
We analyzed the data using a combination of multivariate and univariate procedures to evaluate the variations in the structure of coral assemblages and native hermatypic species, respectively. We used a permutational multivariate analysis of variance (PERMANOVA, Anderson 2001) to test for differences between assemblages of corals in I and NIs, based on Bray–Curtis dissimilarities using 9999 random permutations, with arc-sine square root transformation of the data to achieve homogeneity of dispersions using the software PRIMER 6. The significance level (α) adopted in this analysis and the other analyses was 0.05.
We used analysis of variance (ANOVA) to test for differences in cover of each native coral species in I and NIs using the software STATISTICA version 8.0. A Cochran’s C test (Underwood 1997) and the Kolmogorov–Smirnov test were used to check for homogeneity of variance and normality, respectively. If necessary, data were arc-sine square root transformed to remove variance heterogeneity. If there was no suitable transformation, we performed analyses on the untransformed data. Previous studies argued that ANOVA was robust to departures from the assumption of homogeneous variances when there were many independent estimates of sample variance (e.g., Underwood 1997; Terlizzi et al. 2005a), as in the present study.
Natural encounters
We investigated the natural encounters between the alien coral and any native coral species in I (described above and shown in Fig. 1b) to estimate the percentage of native colonies with tissue damage. We estimated the total number of colonies of each native species contacting T. tagusensis in 80 photographs taken as previously described. We considered it a natural encounter when one colony of T. tagusensis was in contact with, or closer than 5 cm to, a native coral colony and when tissue lesions and algae overgrowth were observed (Dai 1990; Van Veghel et al. 1996). The distance between 1 and 5 cm was defined based on the different mechanisms of competition (e.g., sweeper tentacles, sweeper polyps, elongated polyps and mesenteric filaments) that can cause tissue damage on neighboring corals within this range (Lapid et al. 2004; Lapid and Chadwick 2006; Dos Santos et al. 2013). We also conducted a night dive to observe coral competitive morphological structures (e.g., sweeper tentacles and elongated polyps) in open polyps acting during natural encounters.
Manipulative experiment
We selected two well-distributed coral species of the Brazilian reefs (M. cavernosa and S. stellata) to evaluate the direct competitive encounter effects of the invasive coral T. tagusensis. These two native species were chosen because they occurred in the study area and were abundant in several other coral reefs (non-invaded, see Ferreira and Maida 2006; Cruz et al. 2009) susceptible to invasion in the northeastern and northern regions of the Brazilian coast. We established a manipulative experiment in the field (Cascos Reef) using paired species (alien-native) and corresponding controls (single colony of one species) attached to polyethylene plates (20 cm × 20 cm) as experimental units. Overall, we collected 42 colonies (18 of T. tagusensis, 12 of M. cavernosa and 12 of S. stellata) with diameters between 8 cm and 12 cm to minimize potential size effects (e.g., Zilberberg and Edmunds 2001). We attached the colonies to experimental plate units with non-toxic marine epoxy (Tubolit Men) immediately after collection. After allowing the epoxy to harden in a tank for 1 h, we attached the plate units to 20 cm × 20 cm × 40 cm cement blocks, which we had previously placed on the coral reef substrate at a depth of 21 m, with cable ties. We distributed the experimental units randomly on the substrate. We used two treatments: (1) Contact, where paired alien-native colonies (T. tagusensis with M. cavernosa and T. tagusensis with S. stellata) were placed 5 mm apart to avoid skeletal contact, but allowing tissue contact when their tentacles were expanded; (2) No Contact, where a single colony of each of the three species was placed alone one in the experimental unit. Each treatment had six replicates (n = 6).
The experiment lasted for 90 days, and our observations were made on days 5, 15, 30, 60 and 90 using SCUBA diving and underwater photography (high-resolution digital images). After 3 months, we performed a night dive to observe competitive morphological structures when the coral polyps were expanded. We quantified tissue necrosis of the native species by measuring the necrosed area (cm2) with photographs taken at each time, using CPCe (Kohler and Gill 2006). We used a one-way repeated measures ANOVA to assess the possible differences in the mortality of native species with the following factors: Treatment (fixed, two levels: Contact and No Contact) and Time (fixed, five levels: 5, 15, 30, 60 and 90 days), with time as a repeated measurement factor. To test the homogeneity of variance and normality, we used Cochran’s C and the Kolmogorov–Smirnov statistic, respectively, using STATISTICA software, version 8.0. We applied a square root transformation to the mortality data prior to the analysis to remove variance heterogeneity. A post hoc Tukey test was used when significant differences were found.
Results
Assemblage structure in invaded and non-invaded zones
The top five most abundant coral species (mean cover ≥1 %) in the Cascos Reef were M. cavernosa (14.1 ± 1.3, mean ± SE), T. tagusensis (8.9 ± 1.3), M. decactis (5.1 ± 0.7), M. hispida (2.8 ± 0.4) and S. stellata (1.4 ± 0.2). In the invaded zone, T. tagusensis was dominant (26.7 ± 3.1) and its cover was higher on reef wall (34.9 ± 4.2) than on reef top (18.5 ± 4.1). In the non-invaded zones, the native coral M. cavernosa was dominant (11.6 ± 1.7 in NI1 and 16.8 ± 2.2 in NI2) and more abundant on reef top (18.6 ± 2.7 in NI1 and 27.8 ± 3.4 in NI2) than on reef wall (4.5 ± 1.5 NI1 and 5.7 ± 1.6 NI2). The PERMANOVA showed significant differences in the cover pattern of coral assemblages between I and NIs (Table 1). No significant differences were detected between NIs, although the assemblages differed significantly between habitats (top and wall, Table 1; Fig. 2 “Native Hermatypic Corals”).

Mean coral cover (±SE) within habitats in invaded and non-invaded zones. The different letters denote significant differences. Native Hermatypic Corals represent the sum of native species covers
The ANOVA analysis showed significant differences for M. decactis and M. hispida between reef walls at I and NIs (F (1,239) = 4.96, p < 0.05 and F (1,239) = 7.53, p < 0.01, respectively), and four native corals showed differences between habitats (M. decactis, F (2,237) = 31.38, p < 0.01; M. hispida, F (2,237) = 7.43, p < 0.01; M. cavernosa, F (2,237) = 45.75, p < 0.01; and S. stellata, F (2,237) = 13.63, p < 0.01, Fig. 2).
Natural encounters
We observed 141 natural encounters between the invasive coral and the four most abundant native coral species (M. cavernosa, M. decactis, M. hispida and S. stellata). Colonies of native corals showed partial mortality on the side facing the neighboring invader in 21 % of the encounters (n = 30). S. stellata and M. hispida had the highest percentage of colonies with partial mortality, 52 % (n = 10) and 50 % (n = 7), respectively (Fig. 3). M. cavernosa and M. decactis had the lowest percentage of colonies with partial mortality, 0.1 % (n = 2) and 20 % (n = 11). At night, we observed M. cavernosa and M. decactis using sweeper tentacles against the elongated polyps of T. tagusensis (Fig. 4).

Percentage of natural encounters with mortality in each native coral species on the side facing Tubastraea tagusensis: a Siderastrea stellata, b Madracis decactis, c Montastraea cavernosa and d Mussismilia hispida. The number at the base of each bar represents the total number of colonies of each species in contact with T. tagusensis

Sweeper tentacles (arrows) observed in natural encounters involving competition: a–c Montastraea cavernosa and d Madracis decactis
Manipulative experiment
During the first 15 days of the experiment, the native coral S. stellata was healthy and no significant mortality was found between Contact and No Contact treatments (Fig. 5). However, after the 30th day, the area with tissue necrosis in S. stellata became more pronounced (1.3 ± 1.1 cm2), and bleaching and turf algae growth was also observed in the Contact treatment. At this time, we frequently observed hermit crabs in the contact zones between corals. At night, we also recorded elongated polyps of open T. tagusensis between dead and living zones in the S. stellata colonies. The mortality area of S. stellata increased significantly on the 60th and 90th days (3.2 ± 1.4 and 3.4 ± 1.3 cm2, respectively, Fig. 5; Table 2) and no tissue necrosis was observed in the colonies of T. tagusensis on the side facing S. stellata. Furthermore, all colonies of the No Contact treatment were healthy during the entire experiment.

Mortality (mean and SE) of native species in treatments (Contact and No Contact) in a manipulative experiment over 90 days (n = 6)
On the 5th day of the experiment, the mortality of M. cavernosa was 0.04 ± 0.03 cm2 (Fig. 5) and no necrosis was observed in T. tagusensis during this period in the Contact treatment. On day 15, the necrosis in M. cavernosa increased slightly (0.1 ± 0.04 cm2), followed by the growth of turf and fleshy algae; however, this pattern was also observed in T. tagusensis. Over time, mortality area on M. cavernosa was insignificant (Table 2) and only a small portion of its colony tissue was lost during the entire experiment (0.2 ± 0.1 cm2). We also observed that the number of polyps increased (one to seven polyps) by budding in all six replicates. These observations indicated that M. cavernosa remained in good condition during the experiment. In addition, M. cavernosa showed the ability to attack the alien coral. We recorded the sweeper tentacles of M. cavernosa reaching the living edge of T. tagusensis colonies at night (Fig. 4) that resulted in progressive tissue necrosis in the alien coral over time. By day 30, we observed the growth of turf algae in the mortality area of T. tagusensis colonies and occurrences of hermit crabs in contact zones. At 60–90 days, additional T. tagusensis polyp mortality occurred together with coenosarc discoloration, deterioration of the skeleton, growth of epibionts (crustose coralline algae and turf algae) and occurrences of polychaetes and hermit crabs.
Discussion
We showed that the structure of coral assemblages in Cascos Reef were different in reef zones with and without the presence of the alien coral. The invaded zone in Cascos Reef was dominated by T. tagusensis, especially on the vertical surfaces (i.e., reef wall), and this zone showed significantly lower abundances of the native hermatypic corals, M. decactis and M. hispida. We also found natural differences in patterns of native coral cover between habitats (as observed by Glynn 1976; Francini-Filho et al. 2013) with higher cover of the native zooxanthellate coral on horizontal surfaces, indicating that the alien coral likely found less free space in this habitat than on vertical surfaces. On vertical surfaces, we found a high number of natural encounters between alien and native corals with tissue mortality in colonies of S. stellata, M. hispida and M. decactis, indicating a large number of competitive interactions. A manipulative experiment confirmed that the tissue mortality on S. stellata was significantly higher when in contact with T. tagusensis than without contact. This was similarly observed for M. hispida by Dos Santos et al. (2013). However, our experiment also showed that M. cavernosa had insignificant mortality and reacted to the presence of T. tagusensis developing sweeper tentacles and damaging the alien colonies. This native coral was dominant on all reef tops and its cover did not differ between invaded and non-invaded zones, suggesting possible resistance to the invader effects.
A similar pattern was reported on the rocky shores of southeast Brazil, where the high cover of the alien coral (e.g., on vertical surfaces) was positively correlated with changes in native assemblage structure (Lages et al. 2011). Vertical surfaces, with less light than horizontal surfaces, seems to have optimal conditions for the rapid growth of azooxanthellate T. tagusensis in Brazilian rocky and coral reefs (De Paula and Creed 2005; Mizrahi et al. 2014; present study). Light is a key source of energy for reef-building, zooxanthellate corals via photosynthesis (autotrophy) for successful growth and competition in horizontal habitats (Muscatine 1990). This could explain the preference of the alien coral for shaded habitats where they can find greater release from competition with native zooxanthellate corals than in high-irradiance habitats (Mizrahi et al. 2014). The traditional niche-based models of interspecific competition propose that one species can dominate a community, leading to either resource partitioning between the species, or elimination of the weaker competitor from the habitat (Colwell and Fuentes 1975; Diamond 1978). However, this hypothesis needs further testing.
Our results showed that alien coral dominance in the invaded zone (e.g., reef wall) can be related, at least in part, to their competitive abilities. We observed in natural encounters a high percentage of colonies of the native corals M. hispida and S. stellata with partial mortality on the side facing the neighboring invader. Several authors have suggested that cup coral species from the Dendrophylliidae family (i.e., T. tagusensis, T. coccinea, T. micrantha and Balanophyllia elegans) are aggressive competitors using tentacular contact that can frequently cause tissue damage to neighboring corals (Wellington and Trench 1985; Creed 2006; Dos Santos et al. 2013) or prevent overgrowth by ascidians (Bruno and Witman 1996). Creed (2006) and Dos Santos et al. (2013) demonstrated by mensurative and manipulative experiments that despite the ability of M. hispida to extrude mesenteric filaments a defense mechanism, high mortality rates were observed on this native coral species when in direct contact with the alien corals T. tagusensis and T. coccinea. This may explain why cover of M. hispida was significantly lowest in the invaded zone. These observations corroborate previous suggestions that the endemic Brazilian coral M. hispida can be reduced or competitively excluded by T. tagusensis (Creed 2006; Dos Santos et al. 2013).
Our experiments also confirmed a negative effect of alien coral contact on the endemic coral S. stellata. We registered tentacular contact of T. tagusensis on native S. stellata by the use mechanism of elongated polyps which likely caused a significant increase of the mortality area observed in this native species 60 days after contact. Despite the fact that there was no test for the effects of mechanical contact on tissue necrosis (coral mimic control as in Box and Mumby 2007; Diaz-Pulido et al. 2011; Lages et al. 2012), our experiment strongly suggests that competition was the important mechanism causing tissue necrosis. The polyps of S. stellata are relatively small, and this may explain its lower ability to compete against the large and extensive polyps of T. tagusensis. This native species showed the lowest cover in the invaded zone, along with M. hispida and M. decactis, but cover was also very low at reef walls on non-invaded zones (Fig. 2), which might impede the detection of significant differences. An alternative explanation for the absence of significant variation in cover variation of S. stellata between zones is the compensation of high adult mortality by high settlement and recruitment rates. Reproductive characteristics of S. stellata (e.g., early reproductive age and brooding) enable mature colonies of small size to release planulae larvae able to settle rapidly on substrate, favoring high recruitment rates in shallow reefs (Barros et al. 2003). This supports the competition–colonization trade-off model (Levins and Culver 1971; Horn and MacArthur 1972). This model proposes that species that are inferior competitors are better colonizers and vice versa (Levins and Culver 1971; Horn and MacArthur 1972). Following this model, inferior competitors are better at colonizing vacant space; however, superior competitors can generally displace the subordinates in time (Amarasekare et al. 2004). Furthermore, the high reproductive capacity of the alien coral (Glynn et al. 2008; De Paula et al. 2014) would accelerate the displacement of S. stellata by strong competition between recruits, and therefore additional studies are needed to assess this issue.
T. tagusensis seemed competitively superior to most of the endemic species when competing for space; however, their aggressive mechanism had no effect on M. cavernosa. This native species had a low percentage of colonies with mortality in natural encounters and the tissue necrosis of this native species did not significantly increase when it was in contact with the invasive coral in the manipulative experiment. This result was most likely due to the ability of M. cavernosa to use sweeper tentacles (Richardson et al. 1979), as observed in the present study. It is probable that these tentacles developed in response to contact with the neighboring invasive coral (Richardson et al. 1979). In fact, 30 days after the beginning of the experiment, the alien coral began to show tissue necrosis. Several authors recorded the development of sweeper tentacles 20–30 days after contact with a neighbor (Wellington 1980; Chornesky 1983; Langmead and Chadwick-Furman 1999; Lapid et al. 2004; Lapid and Chadwick 2006). This ability to retaliate against invasive corals can explain why this species did not show differences in cover between invaded and non-invaded zones, as observed in our manipulative experiment. The cosmopolitan species M. cavernosa appears to offer resistance to competition from this invasive species. However, development of competitive mechanisms such as sweeper tentacles requires high energetic demands to coral which in turn cause a trade-off between the allocation of energy to aggressive interactions versus coral growth or reproduction (Tanner 1997). Thus, long-term competition against the alien coral could alter fitness of the native M. cavernosa.
Overall, our study confirmed negative effects of an alien coral on endemic hard corals as suggested by Creed (2006) and Dos Santos et al. (2013). This may be the trigger for a change in the functional benthic group dominance (Cruz et al. 2014) from hermatypic to ahermatypic coral. This would be dramatic for reef integrity, because hermatypic corals are ecological engineers accumulating high rates of calcium carbonate (Birkeland 1977) that increase reef structural complexity and facilitate the occurrence of a large number of species (Graham and Nash 2012; Graham 2014; Rogers et al. 2014). Thus, alien ahermatypic corals could indirectly affect the physical structure and biodiversity of the reef ecosystem. Due to the importance of biodiversity for ecosystem properties (Hooper et al. 2005), these impacts may represent the loss of ecological function and services (Micheli et al. 2014; Pratchett et al. 2014). Future experiments must address how this invasive species affects coral reef functioning.
Several studies suggested that highly diverse communities were more resistant to invasion than communities with low diversity (Elton 1958; Levine and D’Antonio 1999; Stachowicz et al. 1999). The biotic resistance’ is the ability of a native community to resist change in the face of an invasion (Elton 1958; Kimbro et al. 2013). When compared with Caribbean or Indo-Pacific reefs, Brazilian coral reefs may have low resistance to alien coral impacts due to lower species richness and a higher level of endemism. These characteristics may facilitate the establishment of Tubastraea spp. in other Brazilian reefs and explain the absence of impacts of Tubastraea spp. on the coral assemblages of Caribbean reefs (Sammarco et al. 2012).
Our findings showed that the expansion of the invader to other Brazilian coral reefs along the coast could be extremely negative for the most important coral system on the South Atlantic Ocean. Species distribution modeling showed that habitats occupied by native species along the entire Brazilian coast were suitable for Tubastraea spp. (Riul et al. 2013). The Abrolhos reef complex, the most important marine hotspot of the southern Atlantic (the largest and richest coral reef system, Leão et al. 2003), has not yet been invaded by Tubastraea spp., but a recent study reported this invader on oil platforms south of the Abrolhos Bank (Costa et al. 2014). The recent introduction of this species on Brazilian coral reefs requires the implementation of immediate eradication or control protocols with long-term monitoring in order to better understand the effects on Brazilian reefs. In addition, given that the introduction of Tubastraea spp. in Brazil most likely occurred by ships and oil/gas platforms (Fenner 2001; De Paula and Creed 2004), there is a need for the appropriate rules to supervise maritime trade to reduce the flow of this invasive species to the region.
References
Amarasekare P, Hoopes MF, Mouquet N, Holyoak M (2004) Mechanisms of coexistence in competitive metacommunities. Am Nat 164(3):310–326
Anderson MJ (2001) A new method for non-parametric multivariate analyses of variance. Austral Ecol 26:32–46. doi:10.1111/j.1442-9993.2001.01070.pp.x
Ávila E, Carballo JL (2009) A preliminary assessment of the invasiveness of the Indo-Pacific sponge Chalinula nematifera on coral communities from the tropical Eastern Pacific. Biol Invasions 11:257–264. doi:10.1007/s10530-008-9230-5
Barros MML, Pires DO, Castro CB (2003) Sexual reproduction of the Brazilian reef coral Siderastrea stellata Verrill 1868 (Anthozoa, Scleractinia). Bull Mar Sci 73(3):713–724
Birkeland C (1977) The importance of rate biomass accumulation in early successional stages of benthic communities to the survival of coral recruits. In: Proceedings of 3rd International Coral Reef Symposium, vol 1, pp 15–21
Box SJ, Mumby PJ (2007) Effect of macroalgal competition on growth and survival of juvenile Caribbean corals. Mar Ecol Prog Ser 342:139–149. doi:10.3354/meps342139
Bruno JF, Witman JD (1996) Defense mechanisms of scleractinian cup corals against overgrowth by colonial invertebrates. J Exp Mar Biol Ecol 207:229–241. doi:10.1016/S0022-0981(96)02648-2
Budd AF, Fukami H, Smith ND, Knowlton N (2012) Taxonomic classification of the reef coral family Mussidae (Cnidaria: Anthozoa: Scleractinia). Zool J Linn Soc 166:465–529. doi:10.1111/j.1096-3642.2012.00855.x
Cairns SD (2000) Revision shallow-water azooxanthellate scleractinia western Atlantic. Stud Nat Hist Caribb 75:1–240
Caralt S, Cebrian E (2013) Impact of an invasive alga (Womersleyella setacea) on sponge assemblages: compromising the viability of future populations. Biol Invasions 15:1591–1600. doi:10.1007/s10530-012-0394-7
Carlton JT, Geller JB (1993) Ecological roulette: the global transport of nonindigenous marine organisms. Science 80:78–82. doi:10.1126/science.261.5117.78
Castro CB, Pires D (2001) Brazilian coral reefs: what we already know and what is still missing. Bull Mar Sci 69:357–371
Chadwick NE (1988) Competition and locomotion in a free-living fungiid coral. J Exp Mar Biol Ecol 123:189–200. doi:10.1016/0022-0981(88)90041-X
Chadwick NE, Morrow KM (2011) Competition among sessile organisms on coral reefs. In: Dubinsky Z, Stambler N (eds) Coral reef: an ecosystem in transition. Springer, Dordrecht, pp 347–371
Chornesky EA (1983) Induced development of sweeper tentacles on the reef coral Agaricia agaricites: a response to direct competition. Biol Bull 165:569–581. doi:10.2307/1541466
Coles SL, Eldredge LG (2002) Nonindigenous species introductions on coral reefs: a need for information. Pac Sci 56:191–209
Colwell RK, Fuentes ER (1975) Experimental studies of the niches. Annu Rev Ecol Syst 6:281–310
Coni EOC, Ferreira CM, Moura RL, Meirelles PM, Kaufman L, Francini-Filho RB (2013) An evaluation of the use of branching fire-corals (Millepora spp.) as refuge by reef fish in the Abrolhos Bank, eastern Brazil. Environ Biol Fish 96:45–55. doi:10.1007/s10641-012-0021-6
Connell JH, Hughes TP, Wallace CC, Tanner JE, Harms KE, Kerr AM (2004) A long-term study of competition and diversity of corals. Ecol Monogr 74(2):179–210
Connell JH (1973) Population ecology of reef building corals. In: Jones OA, Endean R (eds) Biology and ecology of coral reefs. Academic Press, New York, pp 205–245
Connell JH (1978) Diversity in tropical rain forests and coral reefs. Science 199:1302–1310. doi:10.1126/science.199.4335.1302
Costa TJF, Pinheiro HT, Teixeira JB, Mazzei EF, Bueno L, Hora MSC, Joyeux JC, Carvalho-Filho A, Amado-Filho G, Sampaio CLS, Rocha LA (2014) Expansion of an invasive coral species over Abrolhos Bank, Southwestern Atlantic. Mar Pollut Bull 85(1):252–253. doi:10.1016/j.marpolbul.2014.06.002
Creed JC (2006) Two invasive alien azooxanthellate corals, Tubastraea coccinea and Tubastraea tagusensis, dominate the native zooxanthellate Mussismilia hispida in Brazil. Coral Reefs 25:350. doi:10.1007/s00338-006-0105-x
Crowder LB, Cooper WE (1982) Habitat structural complexity and the interaction between bluegills and their prey. Ecology 63:1802–1813. doi:10.2307/1940122
Cruz ICS, Kikuchi RKP, Leão ZMAN (2009) Caracterização dos Recifes de Corais da Unidade de Conservação Baía de Todos os Santos para fins de manejo, Bahia, Brasil. J Integr Coast Zone Manag 9:16–36
Cruz ICS, Kikuchi RPK, Creed JC (2014) Improving the construction of functional models of alternative persistent states in coral reefs using insights from ongoing research programs: a discussion paper. Mar Environ Res 97:1–9. doi:10.1016/j.marenvres.2014.01.003
Dai C (1990) Interspecific competition in Taiwanese corals with special reference to interactions between alcyonaceans and scleractinians. Mar Ecol Prog Ser 60:291–297
De Paula AF, Creed JC (2004) Two species of the coral Tubastraea (cnidaria, scleractinia) in Brazil: a case of accidental introduction. Bull Mar Sci 74:175–183
De Paula AF, Creed JC (2005) Spatial distribution and abundance of nonindigenous coral genus Tubastraea (Cnidaria, Scleractinia) around Ilha Grande, Brazil. Braz J Biol 65:661–673. doi:10.1590/S1519-69842005000400014
De Paula AF, Pires DO, Creed JC (2014) Reproductive strategies of two invasive sun corals (Tubastraea spp.) in the southwestern Atlantic. JMBAUK 94(3):481–492
Diamond JM (1978) Niche shifts and the rediscovery of interspecific competition. Am Sci 66:322–331
Diaz-Pulido G, Gouezo M, Tilbrook B, Dove S, Anthony KRN (2011) High CO2 enhances the competitive strength of seaweeds over corals. Ecol Lett 14:156–162. doi:10.1111/j.1461-0248.2010.01565.x
Dos Santos LAH, Ribeiro FV, Creed JC (2013) Antagonism between invasive pest corals Tubastraea spp. and the native reef-builder Mussismilia hispida in the southwest Atlantic. J Exp Mar Biol Ecol 449:69–76. doi:10.1016/j.jembe.2013.08.017
Elton CS (1958) The ecology of invasions by animals and plants. Methuen, London
Fenner D (2001) Biogeography of three Caribbean corals (Scleractinia) and the invasion of Tubastraea coccinea into the Gulf of Mexico. Bull Mar Sci 69:1175–1189
Fenner D, Banks K (2004) Orange cup coral Tubastraea coccinea invades Florida and the Flower Garden Banks, Northwestern Gulf of Mexico. Coral Reefs. doi:10.1007/s00338-004-0422-x
Fernández C, Cortés J (2005) Caulerpa sertularioides, a green alga spreading aggressively over coral reef communities in Culebra Bay, North Pacific of Costa Rica. Coral Reefs 24:10. doi:10.1007/s00338-004-0440-8
Ferreira B, Maida M (2006) Monitoramento dos Recifes de Coral do Brasil. Ministério do Meio Ambiente, Brasília
Francini-Filho RB, Coni EOC, Meirelles PM, Amado-Filho GM, Thompson FL, Pereira-Filho GH, Bastos AC, Abrantes DP, Ferreira CM, Gibran FZ, Güth AZ, Sumida PYG, Oliveira NL, Kaufman L, Minte-Vera CV, Moura RL (2013) Dynamics of coral reef benthic assemblages of the Abrolhos Bank, eastern Brazil: inferences on natural and anthropogenic drivers. PLoS One. doi:10.1371/journal.pone.0054260
Glasby TM (1997) Analysing data from post-impact studies using asymmetrical analyses of variance: a case study of epibiota on marinas. Aust J Ecol 22:448–459. doi:10.1111/j.1442-9993.1997.tb00696.x
Glynn PW (1976) Some physical and biological determinants of coral community structure in the Eastern Pacific. Ecol Monogr 46:431–456. doi:10.2307/1942565
Glynn PW, Colley SB, Maté JL, Cortés J, Guzman HM, Bailey RL, Feingold JS, Enochs IC (2008) Reproductive ecology of the azooxanthellate coral Tubastraea coccinea in the equatorial eastern pacific: part V. Dendrophylliidae. Mar Biol 153:529–544. doi:10.1007/s00227-007-0827-5
Graham NAJ (2014) Habitat complexity: coral structural loss leads to fisheries declines. Curr Biol 24:359–361. doi:10.1016/j.cub.2014.03.069
Graham NAJ, Nash KL (2012) The importance of structural complexity in coral reef ecosystems. Coral Reefs 32:315–326. doi:10.1007/s00338-012-0984-y
Hidaka M, Yamazato K (1984) Intraspecific interactions in a scleractinian coral, Galaxea fascicularis: induced formation of sweeper tentacles. Coral Reefs 3:77–85. doi:10.1007/BF00263757
Hollebone AL, Hay ME (2007) An invasive crab alters interaction webs in a marine community. Biol Invasions 10:347–358. doi:10.1007/s10530-007-9134-9
Hooper DU, Chapin FS, Ewel JJ, Hector A, Inchausti P, Lavorel S, Lawton JH, Lodge DM, Loreau M, Naeem S, Schmid B, Setälä H, Symstad AJ, Vandermeer J, Wardle DA (2005) Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol Monogr 75(1):3–35. doi:10.1890/04-0922
Horn HS, MacArthur RH (1972) Competition among fugitive species in a harlequin environment. Ecology 53(4):749–752
Jones CG, Lawton JH, Shachak M (1994) Organisms as ecosystem engineers. Oikos 69:373–386. doi:10.1007/978-1-4612-4018-1_14
Kimbro DL, Cheng BS, Grosholz ED (2013) Biotic resistance in marine environments. Ecol Lett 16:821–833. doi:10.1111/ele.12106
Kohler KE, Gill SM (2006) Coral point count with excel extensions (CPCe): a visual basic program for the determination of coral and substrate coverage using random point count methodology. Comput Geosci 32:1259–1269. doi:10.1016/j.cageo.2005.11.009
Lages BG, Fleury BG, Pinto AC, Creed JC (2010a) Chemical defenses against generalist fish predators and fouling organisms in two invasive ahermatypic corals in the genus Tubastraea. Mar Ecol 31:473–482. doi:10.1111/j.1439-0485.2010.00376.x
Lages BG, Fleury BG, Rezende CM, Pinto AC, Creed JC (2010b) Chemical composition and release in situ due to injury of the invasive coral Tubastraea (Cnidaria, Sclaractinia). Braz J Oceanogr 58:47–56
Lages B, Fleury B, Menegola C, Creed JC (2011) Change in tropical rocky shore communities due to an alien coral invasion. Mar Ecol Prog Ser 438:85–96. doi:10.3354/meps09290
Lages B, Fleury BG, Hovell AMC, Rezende CM, Pinto AC, Creed JC (2012) Proximity to competitors changes secondary metabolites of non-indigenous cup corals, Tubastraea spp., in the southwest Atlantic. Mar Biol 159:1551–1559. doi:10.1007/s00227-012-1941-6
Lang J (1973) Interspecific aggression by scleractinian corals II. Why the race is not only to the swift. Bull Mar Sci 23:260–279
Lang JC, Chornesky EA (1989) Competition between reef corals: a review of mechanisms and effects. In: Dubisky Z (ed) Ecosystems of the world: coral reefs. Elsevier, Amsterdam, pp 209–252
Langmead O, Chadwick-Furman NE (1999) Marginal tentacles of the corallimorpharian Rhodactis rhodostoma. 2. Induced development and long term effects on coral competitors. Mar Biol 34:491–500. doi:10.1007/s002270050565
Lapid ED, Chadwick NE (2006) Long-term effects of competition on coral growth and sweeper tentacle development. Mar Ecol Prog Ser 313:115–123. doi:10.3354/meps313115
Lapid ED, Wielgus J, Chadwick-Furman NE, Gan R (2004) Sweeper tentacles of the brain coral Platygyra daedalea: induced development and effects on competitors. Mar Ecol Prog Ser 282:161–171. doi:10.3354/meps282161
Leal ICS, Pereira PHC, Araújo ME (2013) Coral reef fish association and behaviour on the fire coral Millepora spp. in north-east Brazil. J Mar Biol Assoc UK 93(6):1703–1711. doi:10.1017/S0025315413000234
Leal ICS, Araújo ME, Cunha SR, Pereira PHC (2015) The influence of fire-coral colony size and agonistic behaviour of territorial damselfish on associated coral reef fish communities. Mar Environ Res 108:45–54
Leão ZMAN, Kikuchi RKP (2001) The Abrolhos reefs of Brazil. In: Seeliger U, Kjerfve B (eds) Coastal marine ecosystems of Latin America. Springer, Berlin, pp 83–96
Leão ZMAN, Kikuchi RKP, Testa V (2003) Corals and coral reefs of Brazil. In: Cortés J (ed) Latin America corals reefs. Elsevier, Amsterdam, pp 9–52
Levine JD, D’Antonio CM (1999) Elton revisited: a review of evidence linking diversity and invasibility. Oikos 87:15–26
Levins R, Culver D (1971) Regional coexistence of species and competition between rare species. Proc Nat Acad Sci 68(6):1246–1248
Micheli F, Mumby PJ, Brumbaugh DR, Broad K, Dahlgren CP, Harborne AR, Holmes KE, Kappel CV, Litvin SY, Sanchirico JN (2014) High vulnerability of ecosystem function and services to diversity loss in Caribbean coral reefs. Biol Conserv 171:186–194. doi:10.1016/j.biocon.2013.12.029
Mizrahi D, Navarrete SA, Flores AAV (2014) Uneven abundance of the invasive sun coral over habitat patches of different orientation: an outcome of larval or later benthic processes? J Exp Mar Biol Ecol 452:22–30. doi:10.1016/j.jembe.2013.11.013
Molnar JL, Gamboa RL, Revenga C, Spalding MD (2008) Assessing the global threat of invasive species to marine biodiversity. Front Ecol Environ 6:485–492. doi:10.1890/070064
Muscatine L (1990) The role of symbiotic algae in carbon and energy flux in reef corals. In: Dubinsky Z (ed) Ecosystems of the world: coral reefs. Elsevier, Amsterdam, pp 75–87
Neves E, Johnsson R, Sampaio C (2006) The occurrence of Scolymia cubensis in Brazil: revising the problem of the Caribbean solitary mussids. Zootaxa 54:45–54
Nunes F, Fukami H, Vollmer SV, Norris RD, Knowlton N (2008) Re-evaluation of the systematics of the endemic corals of Brazil by molecular data. Coral Reefs 27:423–432. doi:10.1007/s00338-007-0349-0
Peach MB, Hoegh-Guldberg O (1999) Sweeper polyps of the coral Goniopora tenuidens (Scleractinia: Poritidae). Invertebr Biol 118(1):1–7. doi:10.2307/3226906
Pérez-Estrada CJ, Rodríguez-Estrella R, Palacios-Salgado DS, Paz-García DA (2013) Initial spread of the invasive green alga Caulerpa verticillata over coral reef communities in the Gulf of California. Coral Reefs 32:865. doi:10.1007/s00338-013-1045-x
Pratchett MS, Hoey AS, Wilson SK (2014) Reef degradation and the loss of critical ecosystem goods and services provided by coral reef fishes. Curr Opin Environ Sustain 7:37–43. doi:10.1016/j.cosust.2013.11.022
Richardson CA, Dustan P, Lang JC (1979) Maintenance of living space by sweeper tentacles of Montastrea cavenosa, a Caribbean reef coral. Mar Biol 186:181–186. doi:10.1007/BF00396816
Riul P, Targino C, Júnior L, Creed J, Horta P, Costa G (2013) Invasive potential of the coral Tubastraea coccinea in the southwest Atlantic. Mar Ecol Prog Ser 480:73–81. doi:10.3354/meps10200
Rogers A, Blanchard JL, Mumby PJ (2014) Vulnerability of coral reef fisheries to a loss of structural complexity. Curr Biol 24:1000–1005. doi:10.1016/j.cub.2014.03.026
Ruiz GM, Carlton JT, Grosholz ED, Hines AH (1997) Global invasions of Marine and estuarine habitats by non-indigenous species: mechanisms, extent, and consequences. Am Zool 37:621–632. doi:10.1093/icb/37.6.621
Sammarco PW, Brazeau DA, Sinclair J (2012) Genetic connectivity in scleractinian corals across the Northern Gulf of Mexico: oil/gas platforms, and relationship to the Flower Garden Banks. PLoS One. doi:10.1371/journal.pone.0030144
Sampaio CLS, Miranda RJ, Maia-Nogueira R, Nunes JACC (2012) New occurrences of the nonindigenous orange cup corals Tubastraea coccinea and T. tagusensis (Scleractinia: Dendrophylliidae) in Southwestern Atlantic. Check List 8:2–4
Seebens H, Gastner MT, Blasius B (2013) The risk of marine bioinvasion caused by global shipping. Ecol Lett 16:782–790. doi:10.1111/ele.12111
Stachowicz JJ, Whitlatch RB, Osman RW (1999) Species diversity and invasion resistance in a marine ecosystem. Science 286:1577–1579. doi:10.1126/science.286.5444.1577
Tanner JE (1997) Interspecific competition reduces fitness in scleractinian corals. J Exp Mar Biol Ecol 214:19–34
Terlizzi A, Benedetti-Cecchi L, Bevilacqua S, Fraschetti S, Guidetti P, Anderson M (2005a) Multivariate and univariate asymmetrical analyses in environmental impact assessment: a case study of Mediterranean subtidal sessile assemblages. Mar Ecol Prog Ser 289:27–42. doi:10.3354/meps289027
Terlizzi A, Scuderi D, Fraschetti S, Anderson MJ (2005b) Quantifying effects of pollution on biodiversity: a case study of highly diverse molluscan assemblages in the Mediterranean. Mar Biol 148:293–305. doi:10.1007/s00227-005-0080-8
Underwood AJ (1992) Beyond BACI: the detection of environmental impacts on populations in the real, but variable world. J Exp Mar Biol Ecol 161:145–178. doi:10.1016/0022-0981(92)90094-Q
Underwood AJ (1997) Experiments in ecology: their logical design and interpretation using analysis of variance. Cambridge University, Cambridge, pp 1–504
Van Veghel MLJ, Cleary DFR, Bak RPM (1996) Interspecific interactions and competitive ability of the polymorphic reef-building coral Montastrea annularis. Bull Mar Sci 58:792–803
Wellington GM (1980) Reversal of digestive interactions between Pacific reef corals: mediation by sweeper tentacles. Oecologia 47:340–343. doi:10.1007/BF00398527
Wellington GM, Trench RK (1985) Persistence and coexistence of a nonsymbiotic coral in open reef environments. Proc Natl Acad Sci 82:2432–2436
Zilberberg C, Edmunds P (2001) Competition among small colonies of Agaricia: the importance of size asymmetry in determining competitive outcome. Mar Ecol Prog Ser 221:125–133. doi:10.3354/meps221125
Acknowledgments
We thank the Non-Governmental Organization PRÓ-MAR, EM Chaves, R Macedo, L Porto, IF Buda Andrade, FL Lorders, M Lima, RD Villegas, T Albuquerque, J Bacafu, GC Lessa and JA Reis-Filho for help in the field; JACC Nunes and M Loiola for several helpful comments on earlier drafts of the manuscript; and Y Costa and A Dorea for help with the map. We also thank N Menezes for the identification of S. stellata. This study was supported by The Rufford Foundation (Small Grant No. 13119-1, Projeto Corais da Baía). RJ Miranda was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, ICS Cruz by Conselho Nacional de Desenvolvimento Científico e Tecnológico - CNPq (scholarship, No. 556755/2010-3) and F Barros by CNPq fellowships (303897/2011-2, 239978/2012-9) and Projeto BTS.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: D. Gochfeld.
Reviewed by Undisclosed experts.
Rights and permissions
About this article

Cite this article
Miranda, R.J., Cruz, I.C.S. & Barros, F. Effects of the alien coral Tubastraea tagusensis on native coral assemblages in a southwestern Atlantic coral reef. Mar Biol 163, 45 (2016). https://doi.org/10.1007/s00227-016-2819-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-016-2819-9