
Abstract
The northeastern offshore population of the pantropical spotted dolphin (Stenella attenuata) in the eastern tropical Pacific remains listed as depleted under the Marine Mammal Protection Act, and recent estimates of abundance show that recovery has been slow. One hypothesis for the slow recovery is that continued chase and encirclement by the tuna fishery negatively affects reproduction. Insufficient life-history sampling in this region over the last two decades makes traditional estimates of reproductive rates impossible. Here, we examine the current reproductive patterns of these dolphins by measuring blubber progesterone (BP) concentrations in biopsy samples to assess pregnancy state. BP was quantified in 212 biopsies from female offshore spotted dolphins sampled between 1998 and 2003 in the northeastern tropical Pacific, and we found that 11.5 % of the biopsied females (mature and immature) were pregnant. The relationship between pregnancy and fishery exposure was analyzed, and we found that pregnant females were exposed to significantly less fishery activity than non-pregnant ones (p = 0.022), suggesting that the fishery may have an inhibitive effect on pregnancy. Spatial analysis indicated that pregnancy was more aggregated than random (p < 0.05) at a scale up to 180-nmi, with the highest proportion pregnant in the mouth of the Gulf of California, an area with relatively low reported fishery activity.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In harvesting yellowfin tuna (Thunnus albacares), ETP purse-seine fishing vessels have exploited the common association between tuna and dolphins, as schools of dolphins are chased, herded, and encircled in order to catch the tuna swimming underneath (Perrin 1969). From the 1950s to the early 1990s, this practice generated high dolphin mortality and a substantial decline in abundance, especially in the northeastern stock (NE) of offshore pantropical spotted dolphins (Stenella attenuata) (Smith 1983; DeMaster et al. 1992; Dizon et al. 1994; Wade 1994), and consequently led to their listing as “highly depleted” under the Marine Mammal Protection Act. Although changes in seining methods in the early 1990s drastically reduced direct fishery mortality (Joseph 1994; Gosliner 1999), this stock has shown indications of a weak, slow, or delayed recovery (Wade et al. 2002; Gerrodette and Forcada 2005; Wade et al. 2007). One explanation for the slow recoveryFootnote 1 is that even though reported fishery mortality has dropped off precipitously, the continued chase and encirclement of these dolphins has unobserved detrimental effects on their health via chronic perturbation, including reducing reproduction (Archer et al. 2004b).
Historically, concern regarding their depleted status had prompted numerous studies into the biology of these animals, often focusing on reproductive rates (many reviewed by Perrin and Hohn (1995)). Early studies employed observers to collect biological samples, including the reproductive tracts of incidentally killed dolphins (Perrin et al. 1976, 1977; Hohn et al. 1985; Myrick et al. 1986). The tracts were examined for fetal presence and other indicators of pregnancy, and the resulting data were used to estimate reproductive rates for the entire population (Perrin et al. 1977; Myrick et al. 1986; Chivers and Myrick 1993; Chivers and Demaster 1994). Thousands of samples were collected in this way from NE offshore spotted dolphins, creating one of the largest collections of reproductive data for any cetacean population ever studied and providing the overwhelming majority of the life-history information we have for these dolphins today (Myrick et al. 1986). However, since 1993, with the decline in dolphin mortality and the cessation of the systematic collection of specimens, too few reproductive tracts have been collected to accurately estimate pregnancy rates using fishery-killed dolphins. In addition, there are concerns about age selectivity of the kill, as neonates and post-weaned juvenile animals are under-represented (Barlow and Hohn 1984; Archer and Chivers 2002). This sample selectivity has substantial effects on different measured reproduction parameters, including the proportion of mature dolphins sampled, which in turn influences estimates of reproductive output for these populations (Barlow 1985).
Recent studies into the relationship between pregnancy status and BP content provide an approach that, with sufficient sampling, could help re-assess pregnancy rates of these dolphins (Mansour et al. 2002; Kellar et al. 2006; Trego et al. 2013). The results from these studies document large non-overlapping differences in BP concentration between known pregnant and non-pregnant (mature and immature) female cetaceans including the pantropical spotted dolphin (Trego et al. 2013) and show that compared to the differences between pregnant and non-pregnant dolphins, BP is relatively homogeneous throughout an individual’s blubber layer (Kellar et al. 2006). In addition, information has been gleaned about the dynamics of BP (Kellar et al. 2006) indicating that delphinid BP levels lag behind those in the blood by less than 14 days; though not a precise, this estimate does help bound the maximum lag-in-timing of the hormone concentrations in the blubber. When coupled with projectile biopsying, in which hundreds of blubber samples can be collected during a routine abundance cruise, BP can be used as a pregnancy diagnostic appropriate for population-level estimates.
In this study, BP levels in the blubber collected with biopsy samples of NE pantropical spotted dolphins were quantified and pregnancy status was determined. We compared the proportion of pregnant females in our sample to those made from the incidental fishery-kill samples in efforts to provide context to the current observed values. We also examined geographic variation in reproduction and correlations between pregnancy status and indices of fishery exposure with the intention to assess whether there is evidence of an effect of the purse-seine fishery on spotted dolphin reproduction.
Methods
Data sources for pregnancy assessment
Historic fishery-kill samples
The historical data on incidental fishery-kills for this study were collected by scientific observers from the National Marine Fisheries Service and the Inter-American Tropical Tuna Commission between 1974 and 1993. The reproductive tracts of dolphins killed during tuna purse-seine trips (n = 1,146) were examined for the presence of a fetus, and both ovaries were collected for subsequent examination and characterization of associated corpora. For each specimen, the time and location of collection were noted along with additional biological data including total specimen length and developmental coloration phase. As with all sample sets covered in this study, we estimated the proportion pregnant as the fraction of all females sampled that were pregnant, regardless of maturity state. In most cetacean literature, “pregnancy rate” is based on the fraction of pregnant females of all sexually mature females. We use the term “proportion pregnant” here to distinguish our index from that parameter. The reason for this difference is that we could not determine the state of sexual maturity for biopsied dolphins. We did not biopsy obvious small calves or large animals accompanied by small calves, but we did target all other animals, which accordingly included juveniles not associated with a large female.
Biopsy samples
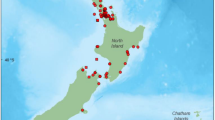
Two types of skin biopsy samples were used in this study. The first type was obtained via crossbow (projectile) darting with a 6-mm-diameter stainless steel collection tip similar to that described in Palsbøll et al. (1991). These biopsies had varying amounts of blubber, ranging from zero to 213 mg, with an average of 98.0 mg. They were most commonly obtained from the posterior flank and tailstock regions posterior to the dorsal fin, because they were collected from bowriding dolphins. The geographic sampling area was large, bounded by the Central American mainland, 111°W, 5 and 25°N (Fig. 1). The samples were collected during four research cruises from 1998 to 2003 (n = 417). This represents about a third of the range of NE pantropical spotted dolphins in the ETP.

Geographic distribution of the female pantropical spotted dolphin blubber samples acquired from projectile biopsying (dark circles) and roto-tagging (light circles) that were analyzed to determine pregnancy state using blubber progesterone concentration
The second type of biopsy was a by-product of attaching plastic identification tags (Roto-tags, Dalton Supplies, Nettlebed, England) to encircled dolphins as part of a 2001 study (Chase Encirclement Stress Studies or CHESS) tracking movement patterns and investigating the effects of chase and encirclement (Scott et al. 1990; Forney et al. 2002; Scott and Chivers 2009). With the application of a tag to the tailing edge of the dorsal fin, a small skin core (3–4 mm in diameter and 2–3 cm in length) was produced (Scott et al. 1990; Scott and Chivers 2009). The blubber from these cores ranged in mass from 0.052 to 0.188 mg (n = 215). The geographic area covered during the 2001 study was much smaller than our projectile biopsy sampling area, and it was centered around 12°N and 97°W (Fig. 1).
Reference samples
We also obtained nine blubber samples from NE pantropical spotted dolphins collected during the 1999–2001 International Dolphin Conservation Program Act necropsy program (Reilly et al. 2005). For these specimens, blubber was collected from the mid-thoracic region (ventral to the dorsal fin) along with observations about fetal presence within the reproductive tract. The blubber samples were trimmed into 100 mg full-thickness pieces (similar in size to a typical biopsy) and then processed and measured identically to all other samples.
Sample processing
Hormone extraction
A FastPrep automated homogenization instrument (Q-Biogene Inc., Irvine, CA) was used to pulverize blubber samples in 1,000 μl of 100 % ethanol following a previously described procedure (Kellar et al. 2006). Unlike in Kellar et al. (2006), after complete homogenization the entire contents of the homogenization tubes were poured (instead of aspirating only 500 μl for further processing) into 12 × 100 mm disposable glass tubes. The homogenization tubes were washed with an additional 500 μl of ethanol. The wash was then collected and added to the 12 × 100 mm collection tubes. A mixture of ethanol and acetone was added to this solution such that the final volume was 4 ml at 4:1. The rest of the hormone extraction procedures were identical to those described in Kellar et al. (2006). The final residue was stored at −20 °C until quantified.
Hormone quantification
A commercially available enzyme immunoassay (ADI-900-011, Enzo Life Sciences, Farmingdale, NY) was used to measure the progesterone in each extraction following a previously described protocol for blubber samples (Kellar et al. 2006). The coefficient of variation (CV) at approximately the 50 % binding concentration for inter-assay and intra-assay comparisons was 3.2 (n = 24) and 4.2 (n = 24), respectively. The manufacturer’s list cross reactivities include 5α-pregnane-3,20-dione (100 %), 17-OH-progesterone (3.46 %), and 5-pregnen-3β-o1-20-one (1.43 %). The rest, corticosterone, 4-androstene-3,17-dione, deoxycorticosterone, DHEA, 17-β estradiol, estrone, estriol, testosterone, and hydrocortisone, were all listed at values less than 1 %. We also empirically tested pregnenlone, a precursor to many of the hormones listed above, and one known to cause high non-targeted binding in other EIA assays (Hecker et al. 2006); we measured 0.16 % cross reactivity in progesterone EIA readings with straight addition of pregnenlone.
Six blubber samples of a single known non-pregnant spotted dolphin female (i.e., known from examinations of reproductive tracts) were spiked with known quantities of cold progesterone (0.0–45.0 ng) to estimate extraction efficiency for each group of extractions (groups ranged from 16 to 30 samples including these controls). The controls were extracted and quantified identically to the rest of the samples. Efficiency was calculated as the amount of measured progesterone minus the amount determined in the non-spiked sample, all divided by the amount of progesterone added (spiked) to the sample prior to extraction. The average efficiency at detection was 82.7 % (SE = 7.8 %).
Sex determination
A standard PCR assay (Berube and Palsboll 1996) was used to determine the sex of the biopsied animals. For 35 of the tagged animals, we had additional data from field determinations of gender (gross examination of external morphology). This information was kept blind until after molecular assaying was complete and therefore provided a check of assay accuracy.
Data analysis
General
All statistical analyses were performed using Matlab 7.0 (The Mathworks Inc., Natick, MA). Chi-squared contingency analyses were used to test the significance of comparisons of proportion pregnant. The estimates from biopsies were compared with those from the 1973–1992 fishery-kill. In addition, the biopsy sampling area was stratified into three equal regions according to the gaps in the biopsy sample distribution, and for each region, the preceding comparisons were re-analyzed.
Statistical significance of spatial variation in proportion pregnant as determined from the biopsies was analyzed using permutation tests of Ripley’s K (Ripley 1977; Besag and Clifford 1989). The K statistic was calculated for the observed samples and then each sample set was randomized with respect to pregnancy state while keeping the proportion pregnant the same as the observed. The resulting permutation tests were repeated for re-constructed datasets in a series of distances (0–1500 nmi); 5,000 replicates were used for each test. The significance of the spatial variation in proportion pregnant was tested against what would be expected at random given the sampling locations.
Fishery effort
A weighted relative index of fishing exposure (Archer et al. 2010) was estimated for each biopsied female using Inter-American Tropical Tuna Commission (IATTC) data indicating the time, date, and location of dolphin sets (i.e., purse-seine operations in which offshore pantropical spotted dolphins were encircled to capture tuna) in the days and months prior to the sampling event in which the biopsy was obtained. The index generally gives greater weight to sets the closer in time and space they were to the sampling event. The weighting system is based on a species-specific movement model (Archer et al. 2010) that incorporates tag-recapture data at different temporal and spatial scales. There was one minor difference in the effort calculation used in this study and the one presented in Archer et al. (2010). Though only sets in which spotted dolphins were observed were used in these calculations, we did not have information about the number of spotted dolphins in each set, and therefore, this weighting cofactor was not incorporated into the final index values. Consequently, the values used in this study are not directly comparable to the exposure values described by Archer et al. (2010).
Two indices were calculated for every biopsy, each reflecting a different temporal/spatial window or ambit (the duration prior to the sampling event in which weighted sets were added together). We used 140- and 180-day ambits in attempts to include a large enough duration to be physiologically meaningful (i.e., a duration in which we expect to be able to quantify any change in proportion pregnant within the population; here approximately equal to half the time of gestation) without making the duration so large that the power to differentiate individual fishery exposure estimates was unduly compromised. The estimated fishery exposure levels were then compared in pregnant and non-pregnant biopsied females using a difference-of-means permutation test (Manly 1991) (Δμ mean), in which the observed difference in mean exposure between pregnant and non-pregnant animals is compared to the distribution of difference-of-mean exposure in iterations (n = 5000) of permuted datasets. This statistical method is comparable to Student’s t test but does not require that parametric assumptions are met; an appropriate approach given that the observed distributions of the fishery exposure indices for our sample set were decidedly multimodal.
Spatial mapping
All maps were created using ArcMap 9.1 (ESRI, Redlands, CA). The ordinary Kriging prediction maps were created as approximate illustrations of the spatial patterns in proportion pregnant and the associated fishery index estimations. All default values were used in the parameter and model selection with the exception that the number of neighboring points which were used for interpolation was raised from 5 to 30 in an effort to capture broad regional patterns instead of local, likely ephemeral, heterogeneities.
Results
Pregnancy rate assessment
BP concentration was measured in nine reference female pantropical spotted dolphins of known pregnancy state (5 immature, 2 non-pregnant mature, and 2 pregnant). Concentrations in the pregnant animals (398.7 ± 128 SE) were on average 63 and 79 times greater than those in non-pregnant mature (5.84 ± 0.36 SE) and immature (8.29 ± 4.0 SE) females, respectively (Table 1). No overlap in BP concentrations was observed between pregnancy states, irrespective of state of sexual maturity. The highest BP measurement of a non-pregnant dolphin was 12.1 ng/g and the lowest measurement for a pregnant dolphin was 269 ng/g.
Of the 632 biopsy samples collected in our study area between 1999 and 2003, 320 were identified as females using the Fain and LaMay (1995) sexing assay; of these, 208 samples (125 projectile biopsy; 83 tagging biopsy) contained sufficient blubber for hormone extraction and analysis.Footnote 2 The progesterone concentration frequency distribution was bimodal, with no value observed between 48.1 and 89.3 ng/g, a finding consistent with the break between known pregnant and non-pregnant delphinids (Fig. 2). We therefore classified those females below 48.1 ng/g as non-pregnant (n = 184) and those above 89.3 ng/g as pregnant (n = 24). This translated into a mean BP concentrations for pregnant and non-pregnant females as 201 ng/g (SE = 24.8) and 6.77 ng/g (SE = 0.62), respectively (Table 1).

Frequency distribution of blubber progesterone concentrations (ng/gram of tissue extracted) in biopsied females pantropical spotted dolphins
Spatial patterns
The Ripley’s K permutation test indicated that, within the biopsy samples, pregnant individuals were more clumped spatially than one would expect at random at a scale from 6.0 to 180-nmi (Fig. 3). Proportion pregnant was greatest, by a substantial margin, in the northern portion of the study area (Figs. 4, 5).

Results of spatial pattern analysis of the distribution of 24 biopsied pregnant pantropical spotted dolphins relative to all females sampled (n = 212) using Ripley’s K statistic. The function K(d) was calculated for each 20-nmi interval (0–300 nmi) and 50-nmi interval (>300 nmi). The derived statistic K(d)/npregnant was plotted against distance (dotted lines). The solid lines represent the 95 % simulation envelope for total spatial randomness. The envelope was derived by permuting (5,000 simulations/distance) pregnancy state for the entire set of sampling sites while fixing the total number of simulated pregnant animals at 24 (equal to the observed number)

Prediction map from an ordinary Kriging interpolation of the proportion pregnant of our biopsies of female pantropical spotted dolphins. The open circles indicate the sampling locations and darker areas denote higher levels of proportion pregnant

Geographic distribution of the female pantropical spotted dolphin biopsies designated as pregnant (dark circles) and non-pregnant (light circles) as determined by blubber progesterone concentrations. Using these data, three geographic strata were delineated (light boxes at breaks in the sampling distribution. These strata were used for comparisons within the historic fishery-kill and between the biopsies and the historic fishery-kill
After discovering this potential spatial pattern in pregnancy, the historic fishery-kill data from our study area were divided into three subregions: north, central, and south (Fig. 4), delineated by breaks in the biopsy sampling distribution. This allowed us to test whether the historic fishery-killFootnote 3 had similar spatial patterns with respect to pregnancy as the biopsies and to assess whether any region had a significantly higher proportion pregnant in the fishery-kill data (Fig. 6). The subsequent 2 × 3 contingency analysis indicated that proportion pregnant in the historic fishery-kill varied significantly among the three regions (χ 2 = 11.81, p = 0.004). Individual 2 × 2 contingency tests showed that the northern region was significantly higher in proportion pregnant than either of the two more southern regions (central: χ 2 = 5.18, p = 0.02; southern: χ 2 = 12.00, p < 0.001), but we found no significant differences between the southern two (χ 2 = 0.80, p = 0.88).

Spatial distribution of pregnant and non-pregnant female pantropical spotted dolphins in the observed historic fishery-kill from 1974 to 1992, constrained to the same area and time of year covered by our biopsy sampling from 1997 to 2003. The light colored boxes indicate the strata of comparison used to (1) investigate spatial differences in the proportion pregnant within the historic kill and (2) assess regional differences in the proportion pregnant between the kill and the biopsies
Comparison of proportion pregnant between historic fishery-kill and current biopsies
We found the proportion of all females biopsied that were pregnant (likely mature plus non-neonate juveniles) was 0.115 (SE = 0.022). This result is significantly lower (χ 2 = 11.04, p < 0.001) than the overall proportion of pregnant females sampled from the historic fishery by catch (0.216 ± 0.011 SE) within the same area. However, after stratifying both datasets into the three geographic subregions, we found that in only two of the three areas were there significantly fewer proportion pregnant females in the biopsies (central χ 2 = 5.08, p = 0.023; southern χ 2 = 7.82, p = 0.004), the exception being the northernmost sampling subregion, at the mouth of the Gulf of California, which was not significantly different from the historic fishery-kill (χ 2 = 1.44, p = 0.22). However, the last can be perhaps ascribed to inadequate statistical power given the smaller sample sizes for the northern strata.
Fishery exposure
In our biopsy sample, pregnant animals had substantially lower fishery exposure indices than non-pregnant ones (Table 2). Permutation tests of the difference in mean exposure (Δμ mean) indicated that these differences were significant for both ambits, the 140-day ambit (Δμ mean = −39.3, p = 0.0220) and the 180-day ambit (Δμ mean = −37.8, p = 0.0188), suggesting that there was an inverse relationship between estimated fishery exposure and proportion pregnant. Moreover, there was also an apparent spatial pattern to the fishery exposure index and the density of sets, with lowest levels seen in the north [where we find the highest proportion pregnant (Fig. 5)] and the highest levels in the south between 10 and 12° latitude (Fig. 7).

Prediction map from an ordinary Kriging interpolation of Archer’s fishery exposure index (140-nmi ambit) for the biopsied female pantropical spotted dolphins. The open circles indicate sampling locations and darker areas indicate higher levels of exposure associated with the samples. This map is a composite of relative exposure levels across all time periods that biopsies were collected and is intended to show that there is geographic variation in these levels. It is not a necessarily an accurate representation of the individual values used for each point as they were collected during different years
Discussion
Pregnancy rate assessment
The frequency distribution of BP concentrations in the pantropical spotted dolphin biopsies was bimodal, with no value between 48.1 and 89.3 ng/g, similar to the values reported for this species in this and previous studies (Trego et al. 2013). This hiatus is similar across all cetacean species examined and in specimens of known reproductive state (Mansour et al. 2002; Kellar et al. 2006; Trego et al. 2013); it reflects the separation between non-pregnant and pregnant femalesFootnote 4 Based on the non-overlapping BP distributions, pregnant females had on average 60 times higher BP concentrations than putative non-pregnant females. Again, these values are similar to those reported for this specimens in animals of known pregnancy state (Trego et al. 2013). Together these pieces of evidence give credence to the use of BP concentration as a diagnostic of pregnancy in these dolphins.
A caveat of these findings is that there are non-pregnancy physiological conditions that could potentially lead to high progesterone levels and thus result in pregnancy misdiagnosis using this method. Corpora lutea associated with ovulation produce higher levels of serum progesterone than those observed in non-pregnant and non-cycling females; however, in the serum these levels are usually lower than average levels seen during pregnancy (Brook et al. 2004). Also, pseudopregnancy, a condition in which no fetus is present yet the reproductive tract functions similar to that of a viable pregnancy can produce elevated serum progesterone (Robeck et al. 2001), potentially at diagnostic levels. However, we believe that both of these conditions have only small effects on the calculated proportion pregnant. Studies examining the historic fishery-kill data found only 2.9 % of all female spotted dolphins with a CL (n = 242) had no associated pregnancy detected, i.e., no observed conceptus or fetus (Perrin et al. 1976; Benirschke et al. 1980). If we make the assumption that no small conceptus or fetus was overlooked in these studies, then we would expect that this 2.9 % was the maximum estimate of the percentage of pseudopregnancies and/or ovulations in these studies’ sample sets. Assuming that same 2.9 % applies to the current sampled population that would suggest our proportion pregnant estimate should be corrected lower but only marginally, from 11.5 to 11.2 %.
Fishery exposure
It has been hypothesized that fishery-induced stress from frequent repeated chase and encirclement of dolphin schools might lead to reduced reproductive success in the highly affected populations (Curry 1999; Archer et al. 2004b; Cramer et al. 2008). Our finding that pregnant animals were estimated to have less previous estimated fishery exposure than non-pregnant animals is consistent with this hypothesis. Individual dolphins are chased, captured, and released on average of eight times a year (2–50 times annually depending on school size) (Perkins and Edwards 1999)Footnote 5 with animals in larger schools exposed at a rate of approximately once a week. The high annual rate of chase and encirclement of dolphins during tuna fishing may constitute a chronic stressor that negatively affects reproductive physiology, altering the rate of pregnancy throughout this population.
These results are consistent with the Cramer et al. (2008) study that showed a negative correlation between the proportion of calves in pantropical spotted dolphin populations and fishing effort. In other Cetartiodactyla, numerous stressors (e.g., noise, isolation, restraint, social separation, and social aggression), including some that may be more benign than chase and encirclement, have been shown to affect all major aspects of reproductive endocrine activity, leading to irregular estrus, fewer ovulations, fewer successful fertilizations, implantation failure, spontaneous abortion, and elevated infant mortality (Moberg 1976, 1991; Hennessy and Williamson 1983; Wilson et al. 1998a, b). How this interference may affect the proportion pregnant is difficult to predict and depends on which stage of pregnancy or reproduction in general is most vulnerable to frequent chase and encirclement. We discuss three potential scenarios here:
-
1.
If the most vulnerable aspects of reproduction are ovulation, fertilization and/or early pregnancy, then one would expect to find a lower proportion pregnant in areas with higher fishing effort. It is well known in the mammalian literature that chronic stress has negative impacts on ovulation (Moberg 1976; Coubrough 1985). Any effect on ovulation rate would be difficult to quantify, but there is evidence that early fetal mortality of dolphin populations impacted by the fishery may be high (Perrin et al. 2003).
-
2.
If late pregnancy and neonate survival are more vulnerable, then we might expect a positive correlation between proportion pregnant and chase and encirclement frequency because mothers with calves are physiologically less likely to become pregnant (lactation has inhibitive effects on ovulation) (McNeilly 2001). If a calf is killed well before weaning or as a fetus before birth, then it is likely that its mother would become pregnant again earlier than had the calf or fetus not died. If this situation occurred at a sufficient rate, it would lead to a measurably higher proportion pregnant within the population (but a lower birth rate). Several studies have indicated that dolphin calves, especially neonates, are disproportionally more vulnerable to chase and recapture than dolphins of other demographic classes (Archer et al. 2001, 2004a; Edwards 2006; Noren and Edwards 2007).
-
3.
It could be some combination of ‘1’ and ‘2’, in which higher than normal prenatal and natal mortality effect both of these vulnerable life-history stages.
Given the fact that we found pregnant animals had significantly lower fishery exposure levels, the results of this study are more concordant with scenarios ‘1’ and ‘3’; they suggest that the main effect of fishery exposure is that it reduces the likelihood of a female being pregnant (i.e., either becoming pregnant or maintaining a pregnancy). There are several alternative explanations that could explain this result that might also have a similar spatial pattern, for example, any oceanographic/habitat conditions that increase the likelihood of becoming and staying pregnant might be poor for tuna fishing (see the Spatial Patterns section, for additional discussion). Second, the sampling in this study was not random; the biopsies were taken opportunistically, leading to clumped sampling in both time and space. This may result in some pseudo-replication characteristics within the sample set. Currently, there are too few samples, especially of pregnant animals (n = 24) to adequately address this concern.
Comparing proportion pregnant between historic fishery-kill and biopsy data
While the observed regional pattern in proportion pregnant from the fishery-kill specimens and the biopsy samples are similar, the absolute values differ. This likely reflects differences in sampling biases in the two datasets. It appears that differential age selectivity of the fishery lead to a disproportionately high proportion pregnant in the measured fishery-kill. Age-frequency distributions of spotted dolphins collected during the kill indicate that it is biased; juveniles (5–15 year olds) are substantially under-represented (Archer and Chivers 2002). This has potentially large ramifications for the historic fishery/biopsy comparison. Recall that the metric, “proportion pregnant” reported in this study, is the number of pregnant females as a fraction of all females sampled. By disproportionally sampling non-juvenile females, we expect that the proportion pregnant within the fishery-kill is biased higher than what we would find in the actual population. If biopsy sampling is less biased or differently biased with respect to age, this could easily account for the differences seen in proportion pregnant in this studyFootnote 6.
Other plausible explanations include (1) natural temporal variation in reproduction is such that biopsies were obtained during a particularly low pregnancy period, (2) changes in habitat during the intervening years leading to reduced reproductive capacity for the population, and (3) stochasticity in the data not accounted for in this investigation. However, given the known age selectivity of the fishery-kill, it is difficult to tease out the role that these factors might have in the disparity of proportion pregnant between the kill and the biopsies.
Spatial patterns
The spatial variability analyses indicated that pregnancy state was not randomly distributed with respect to space. Both the fishery and biopsy data indicated that pregnancy rates were highest in the mouth of the Gulf of California. The explanation may be as simple as that the different areas contain demographically distinct populations with different reproductive rates. There are several additional plausible explanations for this type of selective spatial clumping:
Pregnant females move to areas of higher forage density
Many different mammals segregate by demographic or reproductive condition by selecting habitats that can meet their specific nutritional requirements (Seagle and McNaughton 1992; Labisky and Fritzen 1998; Bowyer 2004; Millspaugh et al. 2004; Ciuti et al. 2006). Pregnant females, especially during mid- to late gestation, experience large increases in energy requirements (Millar 1977; Jonsson et al. 1995; Reynolds and Kunz 2000) and changes in dietary needs that can prompt movement to more productive or more nutritionally appropriate habitats (Bonenfant et al. 2002). In our case, we find the highest proportion pregnant in the mouth of the Gulf of California, an area with relatively high productivity (Collins et al. 1997; Bustos-Serrano and Castro-Valdez 2006) and an additional local maximum near the northwestern edge of the Costa Rica Dome another oceanographic feature associated with upwelling and relatively high productivity (Fiedler and Reilly 1994; Fiedler 2002). Therefore, “the differential nutritional requirements concept” appears to be an explanation consistent with our observations. However, we know that directly after pregnancy, the nutritional requirements are on average even higher while lactating (Hadjipie and Holmes 1966; Millar 1975, 1977; Bowen et al. 2001); in some mammals, the difference is as much as three times as great (Millar 1977). So although nutrition may be a driver for reproductive partitioning or differential location selectivity, we would not necessarily expect only pregnant animals to be disproportionately occupying the areas of highest nutritional output or highest prey concentration; lactating females would be subject to the same drive. In addition, this explanation presupposes that there is some drawback to these areas (that is currently unknown) such that when a post-weaned female is not either pregnant or lactating, she would more likely than chance move to an area of more limited nutritional resources; otherwise, one would expect that all animals, regardless of reproductive condition, would seek out areas of high productivity.
Mature females segregate spatially
Because we cannot differentiate maturity state in biopsied females, it also may be that mature females are disproportionately segregating from immature/post-weaned animals on a regional basis. If this is true, the factors delineated above may still contribute to the spatial variability we find in pregnancy state, but they would be acting on somewhat different physiological, behavioral, and/or social attributes associated with the different maturity groups instead of the different pregnancy states. Previous studies have observed selective maturity partitioning in several free-ranging dolphin and porpoise species. They show reproductively immature animals schooling differently and in different areas compared with mature dolphins (Kasuya 1972, 1976; Kasuya and Ogi 1987). However, as noted above, the fishery-kill data do not suggest strong segregation by demographic or reproductive state.
Chase and encirclement altered reproductive physiology regionally
Because fishing effort is not evenly distributed in space with respect to dolphin abundance, we expect that dolphins in certain areas experience different levels of chase and encirclement; this is evident in the fishery exposure analysis (Archer et al. 2010). If chase and encirclement is generating altered physiological condition at a level that negatively affects reproduction as suggested by the exposure analysis, it might alter the relative spatial distribution of pregnancy.
Again the specific effects of high fishery exposure on pregnancy rate could be complicated, depending on which aspects of reproduction are most vulnerable; for instance, we might not necessarily expect that high fishery exposure would lead to lower pregnancy rates, especially if the late pregnancy or early postnatal periods are most affected. As noted above, this is because mortality at these stages may allow mothers to become pregnant again faster than had they successfully raised and weaned their offspring.
It may be that more than one of these potential factors play some role in the geographic variation of pregnancy. However, irrespective of the potential causes, we reiterate that there does not appear to be strong segregation (though it is statistically significant) by either pregnancy or maturity state, for instance, in the historic fishery-kill, as we find all demographic classes are represented in all areas examined in the kill data. Instead, the signal suggests a disproportionate preference of certain areas by different reproductive/demographic groups.
Conclusions
Though the evidence we present does not establish a definitive causal relationship between purse-seining sets on dolphins and decreased pregnancy rates, it is consistent with the hypothesis that repeated chase and encirclement of dolphins by ETP tuna purse-seine fishery negatively impacts dolphin reproduction. This hypothesis is consistent with the lower numbers of pantropical spotted and eastern spinner dolphin (S. longirostris orientalis) calves observed during years with high fishing effort (Cramer et al. 2008) and in general, the relationship of anthropogenic stressors associated with reduced reproductive success in other free-ranging wildlife populations (Harrington and Veitch 1992; Ellenberg et al. 2007; French et al. 2011). If NE pantropical spotted dolphin reproduction is negatively impacted by tuna purse-seining, this would help explain the observed slow recovery in abundance of this depleted population (Gerrodette and Forcada 2005; Wade et al. 2007), as it would result in a lower rate of recruitment. Given the results of this and previous studies examining the potential impacts of the fishery on the recovery of the pantropical spotted dolphin population in the ETP, additional research examining the direct effects of chase and encirclement on dolphin reproduction is needed.
Notes
Other plausible explanations are summarized in Gerrodette and Forcada (2005) and include indirect reduction of dolphin survival, underreporting of dolphin by catch, and fundamental shifts in habitat conditions.
The ratio of usable samples (those with sufficient blubber) out of all females sampled, 212/320 was particularly low for this sample set because many of the samples have been subsampled for other studies (population genetics and immuno-histochemical staining to characterize protein composition).
Because there was no a priori stratification of the biopsy data, we did not compare the proportion pregnant between these regions with this sample set.
Given sufficient sampling, it is likely that one would find intermediate values indicating a transition between pregnancy states or perhaps as a signal of ovulation. However, because these events are short in duration relative to the amount of time a female spends in pregnancy, lactation, or as an immature animal over her lifetime, they would be rare events and therefore difficult to detect via biopsy sampling.
The duration of each dolphin set is on the order of one to 2 h with problematic sets lasting longer (Curry 1999).
Currently, we do not know how or whether biopsy sampling is selective relative to age. We do know that calves under a year old and mothers with calves under a year old were not targeted for biopsy sampling. With this restriction, one would expect that the biopsy data would be, like the fishery-kill, biased high compared to the fraction of pregnant females within the actual population because female calves cannot be pregnant and mothers with calves are disproportionally less likely to be pregnant compared to other adult females (lactation generally suppresses ovulation). However, it is unknown whether this bias is as large as that resulting from the fishery-kill; consequently, it is uncertain whether the differences in selectively of the two sampling methods (i.e., fishery by catch and biopsy) are solely responsible for the large differences in proportion pregnant.
References
Archer FI, Chivers SJ (2002) Age structure of the northeastern spotted dolphin incidental kill by year for 1971–1990 and 1996–2000. NOAA Admin Rep LJ-02-12
Archer F, Gerrodette T, Dizon A, Abella K, Southern S (2001) Unobserved kill of nursing dolphin calves in a tuna purse-seine fishery. Mar Mammal Sci 17:540–554
Archer F, Gerrodette T, Chivers S, Jackson A (2004a) Annual estimates of the unobserved incidental kill of pantropical spotted dolphin (Stenella attenuata) calves in the tuna purse-seine fishery of the eastern tropical Pacific. Fish Bull 102:233–244
Archer FI, Ballance LT, Barlow J, Chivers SJ, Cramer K, Dizon AE, Donahue M, Edwards EF, Gerrodette T, Kellar NM, Mesnick SL, Perrin WF, Perryman WL, Pitman RL, Reilly SB (2004b) A research plan to investigate fishery effects on reproduction and post-natal mortality. NOAA Admin Rep LJ-04-03
Archer FI, Redfern JV, Gerrodette T, Chivers SJ, Perrin WF (2010) Estimation of relative exposure of dolphins to fishery activity. Mar Ecol Prog Ser 410:245–255. doi:10.3354/meps08641
Barlow J (1985) Variability, trends, and biases in reproductive rates of spotted dolphins, Stenella attenuata. Fish Bull 83:657–669
Barlow J, Hohn A (1984) Interpreting spotted dolphin age distributions. NOAA Tech Memo NMFS-SWFSC-48
Benirschke K, Johnson ML, Benirschke RJ (1980) Is ovulation in dolphins, Stenella longirostris and Stenella attenuata, always copulation-induced? Fish Bull 78:507–528
Berube M, Palsboll P (1996) Identification of sex in cetaceans by multiplexing with three ZFX and ZFY specific primers. Mol Ecol 5:283–287. doi:10.1111/j.1365-294X.1996.tb00315.x
Besag J, Clifford P (1989) Generalized Monte-Carlo significance tests. Biometrika 76:633–642
Bonenfant C, Gaillard JM, Klein F, Loison A (2002) Sex- and age-dependent effects of population density on life history traits of red deer Cervus elaphus in a temperate forest. Ecography 25:446–458. doi:10.1034/j.1600-0587.2002.250407.x
Bowen WD, Iverson SJ, Boness DJ, Oftedal OT (2001) Foraging effort, food intake and lactation performance depend on maternal mass in a small phocid seal. Funct Ecol 15:325–334. doi:10.1046/j.1365-2435.2001.00530.x
Bowyer RT (2004) Sexual segregation in ruminants: definitions, hypotheses, and implications for conservation and management. J Mamm 85:1039–1052. doi:10.1644/Bbl-002.1
Brook F, Lim EHT, Chua FHC, Mackay B (2004) Assessment of the reproductive cycle of the Indo-Pacific humpback dolphin, Sousa chinensis, using ultrasonography. Aquatech Mamm 30:137–148
Bustos-Serrano H, Castro-Valdez R (2006) Flux of nutrients in the Gulf of California: geostrophic approach. Mar Chem 99:210–219. doi:10.1016/j.marchem.2005.09.012
Chivers SJ, Demaster DP (1994) Evaluation of biological indices for three eastern Pacific dolphin species. J Wildl Manage 58:470–478
Chivers SJ, Myrick AC Jr (1993) Comparison of age at sexual maturity and other reproductive parameters for two stocks of spotted dolphin, Stenella attenuata. Fish Bull 91:611–618
Ciuti S, Bongi P, Vassale S, Apollonio M (2006) Influence of fawning on the spatial behaviour and habitat selection of female fallow deer (Dama dama) during late pregnancy and early lactation. J Zool 268:97–107. doi:10.1111/j.1469-7998.2005.00003.x
Collins CA, Garfield N, Mascarenhas AS, Spearman MG, Rago TA (1997) Ocean currents across the entrance to the Gulf of California. J Geophys Res Oceans 102:20927–20936. doi:10.1029/97jc01302
Coubrough RI (1985) Stress and fertility: a review. Onderstepoort J Vet Res 52:153–156
Cramer KL, Perryman WL, Gerrodette T (2008) Declines in reproductive output in two dolphin populations depleted by the yellowfin tuna purse-seine fishery. Mar Ecol Prog Ser 369:273–285
Curry BE (1999) Stress in mammals: the potential influence of fishery-induced stress on dolphins in the eastern tropical pacific ocean. NOAA Tech Memo NMFS-SWFSC-260
DeMaster DP, Edwards EF, Wade PJ, Sisson JE (1992) Status of dolphin stocks in the Eastern tropical pacific. In: McCullough DR, Barrett RH (eds) Wildlife 2001: population. Elsevier Press, London, pp 1038–1050
Dizon AE, Perrin WF, Akin PA (1994) Stocks of dolphins (Stenella spp. and Delphinus delphis) in the Eastern tropical pacific: a phylogeographic classification. NOAA Technical Report NMFS 119
Edwards EF (2006) Duration of unassisted swimming activity for spotted dolphin (Stenella attenuata) calves: implications for mother-calf separation during tuna purse-seine sets. Fish Bull 104:125–135
Ellenberg U, Setiawan AN, Cree A, Houston DM, Seddon PJ (2007) Elevated hormonal stress response and reduced reproductive output in yellow-eyed penguins exposed to unregulated tourism. Gen Comp Endocrinol 152:54–63. doi:10.1016/j.ygcen.2007.02.022
Fiedler PC (2002) Environmental change in the eastern tropical Pacific Ocean: review of ENSO and decadal variability. Mar Ecol Prog Ser 244:265–283. doi:10.3354/Meps244265
Fiedler PC, Reilly SB (1994) Interannual variability in dolphin habitats in the eastern tropical Pacific. II: effects on abundances estimated from tuna vessel sightings, 1975–1990. Fish Bull 92:451–463
Forney KA, St. Aubin DJ, Chivers SJ (2002) Chase encirclement stress studies on dolphins involved in eastern tropical Pacific ocean purse-seine operation during 2001. NOAA Admin Rep LJ-02-32
French SS, Gonzalez-Suarez M, Young JK, Durham S, Gerber LR (2011) Human disturbance influences reproductive success and growth rate in California sea lions (Zalophus californianus). PLoS ONE 6:e17686. doi:10.1371/journal.pone.0017686
Gerrodette T, Forcada J (2005) Non-recovery of two spotted and spinner dolphin populations in the eastern tropical Pacific Ocean. Mar Ecol Prog Ser 291:1–21
Gosliner ML (1999) The tuna-dolphin controversy. In: John R, Twiss J, Reeves RR (eds) Conservation and management of marine mammals. Smithsonian Institution Press, Washington, pp 120–155
Hadjipie G, Holmes W (1966) Studies on feed intake and feed utilization by sheep: voluntary feed intake of dry pregnant and lactating ewes. J Agr Sci 66:217–223
Harrington FH, Veitch AM (1992) Calving success of woodland caribou exposed to low-level jet fighter overflights. Arctic 45:213–218
Hecker M, Newsted JL, Murphy MB, Higley EB, Jones PD, Wu R, Giesy JP (2006) Human adrenocarcinoma (H295R) cells for rapid in vitro determination of effects on steroidogenesis: hormone production. Toxicol Appl Pharmacol 217:114–124. doi:10.1016/j.taap.2006.07.007
Hennessy DP, Williamson P (1983) The effects of stress and of ACTH administration in hormone profiles, estrus and ovulation in pigs. Theriogenology 20:13–26
Hohn AA, Chivers SJ, Barlow J (1985) Reproductive maturity and seasonality of male spotted dolphins, Stenella attenuata, in the eastern tropical Pacific. Mar Mammal Sci 1:273–293. doi:10.1111/j.1748-7692.1985.tb00016.x
Jonsson KI, Tuomi J, Jaremo J (1995) On the consequences of pre- and post- breeding costs in the evolution of reproductive effort tactics. Ecoscience 2:311–320
Joseph J (1994) The tuna dolphin controversy in the eastern Pacific ocean: biological, economic, and political aspects. Ocean Devel Inter Law 25:1–30
Kasuya T (1972) Growth and reproduction of Stenella Coeruleoalba based on the age determination by means of dentinal growth layers. Sci Rep Whales Res Inst Tokyo 24:57–79
Kasuya T (1976) Reconsideration of life history parameters of the spotted and striped dolphins based on cemental layers. Sci Rep Whales Res Inst Tokyo 28:73–106
Kasuya T, Ogi H (1987) Distribution of mother-calf Dall’s porpoise pairs as an indication of calving grounds and stock identity. Sci Rep Whales Res Inst Tokyo 38:125–140
Kellar NM, Trego ML, Marks CI, Dizon AE (2006) Determining pregnancy from blubber in three species of delphinids. Mar Mammal Sci 22:1–16
Labisky RF, Fritzen DE (1998) Spatial mobility of breeding female white-tailed deer in a low-density population. J Wildl Manage 62:1329–1334. doi:10.2307/3801998
Manly BFJ (1991) Randomization and Monte Carlo methods in biology. Chapman and Hall, London
Mansour AAH, McKay DW, Lien J, Orr JC, Banoub JH, Oien N, Stenson G (2002) Determination of pregnancy status from blubber samples in minke whales (Balaenoptera acutorostrata). Mar Mammal Sci 18:112–120. doi:10.1111/j.1748-7692.2002.tb01022.x
McNeilly AS (2001) Lactational control of reproduction. Reprod Fertil Dev 13:583–590
Millar JS (1975) Tactics of energy partitioning in breeding Peromyscus. Canad J Zool 53:967–976
Millar JS (1977) Adaptive features of mammalian reproduction. Evolution 31:370–386. doi:10.2307/2407759
Millspaugh JJ, Brundige GC, Gitzen RA, Raedeke KJ (2004) Herd organization of cow elk in Custer State Park, South Dakota. Wildlife Soc B 32:506–514. doi:10.2193/0091-7648(2004)32[506:Hoocei]2.0.Co;2
Moberg GP (1976) Effects of environment and management stress on reproduction in dairy cow. J Dairy Sci 59:1618–1624. doi:10.3168/jds.S0022-0302(76)84414-1
Moberg GP (1991) How behavioral stress disrupts the endocrine control of reproduction in domestic animals. J Dairy Sci 74:304–311
Myrick AC, Hohn AA, Barlow J, Sloan PA (1986) Reproductive biology of female spotted dolphins, Stenella attenuata, from the eastern tropical Pacific. Fish Bull 84:247–259
Noren SR, Edwards EF (2007) Physiological and behavioral development in delphinid calves: implications for calf separation and mortality due to tuna purse-seine sets. Mar Mammal Sci 23:15–29. doi:10.1111/j.1748-7692.2006.00083.x
Palsbøll PJ, Larsen F, Hansen ES (1991) Sampling of skin biopsies from free-ranging large cetaceans in west Greenland: development of new biopsy tips and bolt designs. Rpt of Internat Whaling Comm. Special Issue 13:71–89
Perkins PC, Edwards EF (1999) Capture rate as a function of school size in pantropical spotted dolphins, Stenella attenuata, in the eastern tropical Pacific Ocean. Fish Bull 97:542–554
Perrin WF (1969) Using porpoise to catch tuna. World Fish 18:42–45
Perrin WF, Hohn AA (1995) Pantropical spotted dolphin Stenella attenuata. Handb Mar Mamm 5:71–97
Perrin WF, Coe JM, Zweifel JR (1976) Growth and reproduction of the spotted porpoise, Stenella attenuata, in the offshore eastern tropical Pacific. Fish Bull 74:229–269
Perrin WF, Miller RB, Sloan PA (1977) Reproductive parameters of the offshore spotted dolphin, a geographic form of Stenella attenuata, in the eastern tropical Pacific, 1973–75. Fish Bull 75:629–633
Perrin WF, Chivers SJ, Archer FI (2003) Fetal mortality in dolphins exploited by the tuna fishery abstracts of the 15th Biennial conference on the biology of marine mammals. The Society of Marine Mammalogy, Greensboro, NC
Reilly SB, Donahue MA, Gerrodette T, Forney K, Wade P, Ballance L, Forcada J, Fiedler P, Dizon A, Perryman WF, Archer FA, Edwards EF (2005) Report of the scientific research program under the international dolphin conservation program act. NOAA Tech Memo NMFS-SWFSC-372
Reynolds DS, Kunz TH (2000) Changes in body composition during reproduction and postnatal growth in the little brown bat, Myotis lucifugus (Chiroptera: Vespertilionidae). Ecoscience 7:10–17
Ripley BD (1977) Modeling spatial patterns. J Roy Stat Soc Ser B 39:172–212
Robeck TR, Atkinson SKC, Brook FM (2001) Reproduction. In: Dierauf LA, Gulland FMD (eds) Marine mammal medicine. CRC Press LLC, Boca Raton, pp 193–236
Scott MD, Chivers SJ (2009) Movements and diving behavior of pelagic spotted dolphins. Mar Mammal Sci 25:137–160. doi:10.1111/j.1748-7692.2008.00241.x
Scott MD, Wells RS, Irvine AB, Mate BR (1990) Tagging and marking studies of small cetaceans. In: Leatherwood S, Reeves RR (eds) The bottlenose dolphin. Academic Press, San Diego, pp 489–514
Seagle SW, McNaughton SJ (1992) Spatial variation in forage nutrient concentrations and the distribution of Serengeti grazing ungulates. Landsc Ecol 7:229–241
Smith TD (1983) Changes in size of three dolphin (Stenella spp.) populations in the eastern tropical Pacific. Fish Bull 81:1–13
Trego ML, Kellar NM, Danil K (2013) Validation of blubber progesterone concentrations for pregnancy determination in three dolphin species and a porpoise. PLoS One. In press
Wade P (1994) Revised estimates of incidental kill of dolphins (Delphinidae) by the purse-seine tuna fishery in the eastern tropical Pacific, 1959–1972. Fish Bull 93:345–354
Wade PR, Reilly SB, Gerrodette T (2002) Assessment of the population dynamics of the northeastern offshore spotted and the eastern spinner dolphin populations through 2002. NOAA Admin Rep LJ-02-13
Wade PR, Watters GM, Gerrodette T, Reilly SB (2007) Depletion of spotted and spinner dolphins in the eastern tropical Pacific: modeling hypotheses for their lack of recovery. Mar Ecol Prog Ser 343:1–14. doi:10.3354/Meps07069
Wilson SJ, Kirby CJ, Koenigsfeld AT, Keisler DH, Lucy MC (1998a) Effects of controlled heat stress on ovarian function of dairy cattle. 2. Heifers. J Dairy Sci 81:2132–2138. doi:10.3168/jds.S0022-0302(98)75789-3
Wilson SJ, Marion RS, Spain JN, Spiers DE, Keisler DH, Lucy MC (1998b) Effects of controlled heat stress on ovarian function of dairy cattle. 1. Lactating cows. J Dairy Sci 81:2124–2131. doi:10.3168/jds.S0022-0302(98)75788-1
Acknowledgments
We like to thank our amazing marine mammal observers and field personnel, especially Juan Carlos Salinas and Ernesto Vazquez, for obtaining many of the biopsies used in this study. Additional thanks go to the crew and officers of the NOAA ships David Star Jordan and the McArthur II for their adept service over these years. Also this project would not have been possible save for the tireless efforts of the tuna boat observers, who over many years brought home quality data despite arduous working conditions. Finally, we thank Wayne Perryman and Bill Perrin for their insightful recommendations of both the research and this manuscript. This research was funded by NOAA Fisheries under the International Dolphin Conservation Program Act.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by U. Siebert.
Rights and permissions
About this article
Cite this article
Kellar, N.M., Trego, M.L., Chivers, S.J. et al. Pregnancy patterns of pantropical spotted dolphins (Stenella attenuata) in the eastern tropical Pacific determined from hormonal analysis of blubber biopsies and correlations with the purse-seine tuna fishery. Mar Biol 160, 3113–3124 (2013). https://doi.org/10.1007/s00227-013-2299-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-013-2299-0