
Abstract
Between 2002 and 2008, samples of the cold-water scleractinian coral Lophelia pertusa were collected from the Trondheim Fjord in Norway to examine reproductive periodicity. Collections were made from three locations: Tautra, (63°35.36′N, 10°31.23′E at 40–70 m), Stokkbergneset (63°28.18′N, 09°54.73′E at 110–500 m), and Røberg (63°28.88′N, 09°59.50′E at 250 m). Populations of L. pertusa from the Trondheim Fjord initiated oogenesis in January and spawning occurred from late January to early March the following year. Gametogenic cycles of the female L. pertusa samples overlapped by approximately 2 months, with oogonia visible in January, but this was not evident in the males. This paper provides the most complete gametogenic cycle to date and spawning observations for this important structure-forming species. The results from fjord populations are compared with published and preliminary data from other regions and are discussed in the context of regional differences in physical and biological variables, particularly food supply. Differences in gametogenic cycles within a single species provide a rare opportunity (especially in deep-sea species) to examine potential drivers of reproduction.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The most widespread and extensively studied of the cold-water, reef-building scleractinian corals is Lophelia pertusa (Linnaeus 1758). This branching stony coral colonizes hard substrata and can form large complex structures, generally at upper slope depths of 200–600 m. The highest known densities of L. pertusa ecosystems occur in the North Atlantic Ocean, along both eastern and western continental margins, and these have provided the basis for most of the current knowledge of the biology and ecology of cold-water corals (Rogers 1999; Freiwald and Roberts 2005; Ross and Nizinski 2007; Brooke and Schroeder 2007; Roberts et al. 2009). Some of the most extensive L. pertusa ecosystems occur along the mid-Norwegian shelf and slope at 200–400 m depths (Mortensen et al. 1995; Freiwald et al. 2002; Fosså et al. 2002; Hovland 2008). Norway is home to both the largest and the shallowest documented L. pertusa communities; the Røst Reef was discovered in 2002 and is over 40 km long (Thorsnes et al. 2004), whereas the shallowest documented (39 m) L. pertusa communities were discovered decades earlier in the Trondheim Fjord (Strømgren 1971).
Cold-water coral ecosystems are threatened globally by anthropogenic activities, such as bottom-fishing, energy industry activities, and the effects of climate change. Highly localized nutrient input from terrestrial discharge (Johannessen and Dahl 1996) and mariculture operations (Asche et al. 1999) may also damage Norway’s shallow fjord coral ecosystems. These threats may result in direct physical damage as from bottom fishing (Koslow and Gowlett-Jones 1998; Rogers 1999; de Juan and Lleonart 2010; Clark and Tittensor 2010), mortality from sediment and toxins (Loya and Rinkevich 1979; Rinkevich and Loya 1979; Rogers 1999; Brooke et al. 2009; Larsson and Purser 2011), changes in calcification from reduced aragonite saturation state (de Putron et al. 2011) or proliferation of coral-encrusting organisms such as sponges (J. Jarnegren, pers. obs.). The tolerance of these deep-sea coral ecosystems to environmental variability is poorly understood (Pandolfi et al. 2011), nor is there adequate information on their ability to recover from natural or anthropogenic damage. Assuming availability of suitable substrate and appropriate environmental conditions, re-colonization of damaged L. pertusa reefs will be strongly influenced by the reproductive output (fecundity) of the adult coral and the dispersal, recruitment and post-settlement survival of the coral larvae (planulae). Understanding connectivity among these deep ecosystems is imperative for effective management, and yet with the exception of a few species (Brooke and Young 2003; Burgess and Babcock 2005; Waller and Tyler 2005), little is known of the reproductive and larval biology of cold-water reef-building species (Roberts et al. 2009; Morrison et al. 2011).
Colonial scleractinian corals grow via asexual replication of polyps, which in L. pertusa occurs by unequal intra-tentacular budding (Cairns 1979, 1995; Roberts et al. 2009). Uncommon modes of asexual reproduction, such as polyp bail-out and parthenogenesis, have been documented for shallow-water corals (Sammarco 1982; Krupp 1983; Stoddart 1983; McFadden et al. 2001; McFadden and Hutchinson 2004), but have not been reported for any cold-water scleractinian species. Many species of coral can also reproduce through fragmentation where pieces of the parent colony break off through mechanical stresses and continue to grow, thereby establishing new colonies that are genetically identical to the parent colonies (clones) (Highsmith 1982). Fragmentation, however, does not account for the establishment of new widely separated coral ecosystems, nor is it a mechanism for genetic exchange between established ecosystems; these require dispersive planktonic larvae produced through sexual reproduction (Harrison 2011). Most of the research on coral reproductive biology has focused on shallow tropical reef-dwelling corals (see reviews by Fadlallah 1983; Baird et al. 2009; Harrison 2011), whereas corals living in temperate, cold, and deep habitats have been neglected by comparison. Although scientific and conservation interests in cold-water corals have expanded rapidly, information on their basic life history patterns is still lacking, partly because obtaining samples throughout the year from cold-water coral ecosystems poses significant logistical challenges. Information on 444 species of scleractinians indicates that hermaphroditic broadcast spawning of gametes is the dominant fertilization mechanism (Harrison 2011). To date, all cold-water, structure-forming scleractinians are also broadcast spawners, but appear to be gonochoristic rather than hermaphroditic (Brooke and Young 2003; Burgess and Babcock 2005; Waller 2005; Waller and Tyler 2005). The duration of larval life is unknown for cold-water scleractinians, with one exception. Laboratory studies on Oculina varicosa, a shelf edge scleractinian species that forms large bioherms off the Florida Atlantic coast, documented a larval lifespan of approximately 14–21 days (Brooke and Young 2005).
Despite a proliferation of research in the last decade, information on the reproductive biology of L. pertusa was not published until quite recently. Using samples collected from the northeast (NE) Atlantic in March, July, August, September, and October, Waller and Tyler (2005) reported a seasonal reproductive cycle with one cohort per year, culminating in a spawning event, probably in February. They also reported equivocal information on gametogenic cycles of M. oculata, suggesting two cohorts per year. In contrast to structure-forming colonial species, which appear to be uniformly gonochoristic broadcast spawners, cold-water solitary scleractinians, such as Flabellum sp., (Waller and Tyler 2011; Mercier et al. 2011), Caryophyllia sp. (Waller et al. 2005), and Fungiacyathus sp. (Waller et al. 2002; Flint et al. 2007), have various reproductive strategies, including hermaphroditism, gonochorism, brooding, and broadcast spawning. The evolutionary significance of these differences is not clear, but larger eggs generally represent a larger maternal investment in the offspring and shorten the larval life and need for an exogenous food supply (Strathmann 1985; Havenhand 1993). Lophelia pertusa has a widespread distribution and can colonize hard substrata in many different habitat types (including oil rigs, shipwrecks, fjords, seamounts, and continental shelves and slopes), over a relatively large temperature range (4–14 °C). Recent genetic work on this species has shown that populations in the North Atlantic are relatively isolated, indicating some limit to dispersal potential (Morrison et al. 2011). More work is needed on larval dynamics and physical oceanography models to resolve the apparent contradictions between the widespread distribution of this species and the relatively limited dispersal rates derived from population structure.
This manuscript presents new data on the reproductive periodicity of L. pertusa using samples from the Trondheim Fjord in Norway. The sheltered conditions and relatively shallow depths of the fjord permitted a more complete temporal sampling and description of gametogenesis than was previously possible. This data provide detailed information on initiation of gametogenesis, periods of maximum yolk deposition (vitellogenesis), and timing and duration of spawning for this important structure-forming species.
Materials and methods
Field sampling
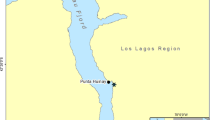
Situated in the middle of Norway’s west coast, the Trondheim Fjord is located between 63°30′–64°N and 09°30′–11°30′E, is 135 km long, and has a maximum depth of 617 m (Fig. 1). The fjord is fed by six large rivers (Smelror 2000) and is typically U-shaped in cross section, with steep walls and a flat base. The fjord is comprised of three basins separated by relatively shallow sills (100–195 m) (Jacobson 1983). The deeper water layers are a mixture of Atlantic Ocean water and the Norwegian Coastal Current, and during late winter and early spring, high salinity water penetrates the deeper parts of the fjord. Current speeds, which to a large extent are tidally driven, reach up to 1.0 m s−1 at the sills (Jacobson 1983).

Collection sites for Lophelia pertusa in Trondheim Fjord, Norway (indicated by circles): Tautra, Stokkbergneset and Røberg
Three different study sites were sampled within the fjord: Tautra (63°35.36′N, 10°31.23′E) at 40–70 m, Stokkbergneset (63°28.18′N, 09°54.73′E) at 110–500 m, and Røberg (63°28.88′N, 09°59.50′E) at ~250 m depth (Fig. 1). The Tautra reef is a shallow sill-reef consisting of large, abundant, and compact cauliflower-shaped L. pertusa colonies (Fig. 2a). At Stokkbergneset and Røberg, the coral colonies are attached to the steep rock walls and hang from the vertical faces or beneath overhangs, creating dense elongated structures (Fig. 2b). On these walls, live L. pertusa was found from 100 m to near the base of the wall at 500 m. Adjacent to the base of the wall are piles of predominantly dead coral colonies, molluscs, sponges, and other fauna that have fallen from the wall onto the soft sediment.

a Colonies of Lophelia pertusa at the Stokkergneset study site are attached to the steep wall at ~250 m, b Colonies at the shallow Tautra site are clustered on the sill at <100 m
Collections were made using the R/V Gunnerus and the remotely operated vehicle (ROV) Minerva, owned by the Norwegian University of Science and Technology (NTNU). The ROV can operate to depths of 700 m and is equipped with four cameras and two manipulator arms (one 5 function hydraulic and one single function electric). The ROV navigation system is the Ultra-short Baseline system Kongsberg HiPAP 500, with EIVA NaviPac and CMAP maps. A total of 30 separate cruises were conducted between November 2002 and March 2008. Fragments of 76 different colonies from the three study sites were processed for histology. Most of the samples were collected from Tautra (16 site visits, 44 samples, depth range 44–120 m), followed by Stokkbergneset (6 site visits, 27 samples, depth range 119–500 m), Røberg (2 site visits, 5 samples, depth range 250–396 m) (Table 1). Samples were collected from every month, except July and September, at least once during the 2002–2008 study period, with more collections focused on the months when spawning was most likely (January, February, and March).
During each collection cruise, between one and eight (mean = 3.17, SD = 1.95), samples (each consisting of 10–20 branches) were taken from different colonies. Coral samples were collected using a landing net attached to the ROV, which could accommodate only one or two samples per dive. Both of the two L. pertusa colony colors (orange and white) were present at all of the study sites. In order to identify each individual colony after collection, either a single colony or one of each color was collected per dive. Sample colors were tracked throughout processing to assess whether there were significant differences in reproductive cycles between the two morphotypes. Gender cannot be determined in the field, but previous collections by the authors indicated that females were more prevalent than males in the fjord populations. Samples were maintained in cold (~6–7 °C) seawater until returned to NTNU, where small fragments (10–15 polyps) of each sample were preserved for histological analysis in 10 % buffered formalin solution.
Histology
Small polyps at the tips of each fragment were not used for histological examination, as they were potentially immature and non-reproductive (Rinkevich and Loya 1987). The remaining polyps (~8–10) were transferred to 5 % hydrochloric acid for 6–10 h to decalcify the skeleton, leaving the tissue intact for further processing. Polyps were rinsed briefly in distilled water and then dehydrated through a series of ethanol concentrations (70, 85, 95, 100 % × 2 changes) and transferred to a clearing agent (toluene or Histoclear©). The polyps were then embedded in paraffin wax (3–5 per block), serially sectioned into 8 μm slices, mounted on glass microscope slides, and stained using Mayer’s Hematoxylin/Eosin B stains. Images of the microscope slides were taken using an Optronics digital camera attached to an Olympus BX50 compound microscope. These images were used for the description of gametogenic status for each individual coral fragment.
For each female fragment, 100 oocytes were measured from three to five polyps (occasionally more polyps were needed to meet the required number of oocytes). Only those oocytes with a visible nucleolus were measured. This ensured that the same egg was not measured more than once, as the nucleolus is small (approximately 9 μm diameter) that it only appears in one 8 μm slice. The area of each oocyte was measured and recorded using Image Tool (UTHSCSA) image analysis software. Oocyte ‘feret’ diameter was calculated, which uses the measured area of the oocyte and estimates the diameter is if it were a circle (Brooke and Young 2003; Waller and Tyler 2005). Feret diameters were used to generate means and standard deviations of oocyte diameters, and size–frequency distributions for each sampling date, which were then used to infer the timing of the female gametogenic cycle. The male reproductive periodicity was documented more qualitatively by developmental stage, as size of spermatocysts does not directly reflect their maturity. The male gametogenic cycle was documented by stages (after Waller and Tyler 2005) as follows: Stage I (early spermatogenesis), spermatocysts are lined with spermatocytes but lumens are empty; Stage II (maturation phase), thick layer of spermatocytes with some spermatozoa present, but with mostly empty lumens; Stage III (mature), spermatocyst lumens are filled with spermatozoa; Stage IV (post-spawn), spermatocysts are empty of spermatozoa, except occasional remnants of spawning.
Results
Gametogenesis
The samples were comprised of both males and females of each color; therefore, colony color was not an indicator of gender in L. pertusa. The gender ratio within the collections was unequal, with more females than males (f:m = 1.27); however, a Chi-squared analysis showed no significant difference between the numbers of each gender (χ2 = 19.76, p = 0.47). Twelve of the samples were devoid of gametes, therefore their genders could not be established (Table 1).
There were insufficient samples from the same time, date, depth, and year to test for statistical differences in mean oocyte feret diameter among sites. Differences among years were tested using one-way ANOVA when sufficient data were available, and no significant differences were found. Table 2 presents a summary of comparisons and statistical outcomes. For this study, discussion of gametogenesis and reproductive periodicity apply to data pooled across color morphs, study sites, and years.
Female gametogenesis
During this project (2002–2008), female samples were collected at least once for each month except May, July, September, and December. Oocytes were found in samples from every collection, indicating continuous reproductive cycles with no intermission between spawning and onset of gametogenesis. Mature oocytes were present from January to March, with the peak in middle to late February. The samples kept alive in the laboratory were observed to spawn from January to March, with each sample releasing gametes multiple times. The smallest oogonia (<10 μm) and pre-vitellogenic oocytes (10–30 μm) were also observed in samples from January, February and March (see Fig. 3a, b for images of different stages of oogenesis), indicating the onset of the next gametogenic cycle. The gametogenic cycle, therefore, began at the end of January and terminated in February–March of the following year so the cycle was approximately 13–14 months in duration, with a one- to two-month overlap between cycles.

a Early stages of Lophelia pertusa gametogenesis: A March: large oocytes in left image are remnants from spawning, and right image shows oogonia signaling the start of the next cycle, B April: oogonia and pre-vitellogenic oocytes, C June: pre-vitellogenic oocytes, D August: early vitellogenesis. Lvo late vitellogenic oocyte, og oogonia, pvo previtellogenic oocyte, vo vitellogenic oocyte. Scale bars A, B = 100 μm; C, D = 50 μm. b Late stages of Lophelia pertusa oogenesis A October: vitellogenic oocytes B November: vitellogenic oocytes become more mis-shapen as they fill the mesenteries C January: mature oocytes with nucleoli near periphery of nucleus D February: pre-spawning oocytes. vo vitellogenic oocyte, mvo mature vitellogenic oocyte, no nucleolus. Scale bars 100 μm
The oogonia developed within the mesenteries and in the early pre-vitellogenic stages were generally aligned along the mesenterial lamellae. As the oocytes grew, they migrated away from the mesenterial lamellae and increased in size, forming cohesive groups, generally toward the posterior end of the mesentery. There were fewer samples collected during the middle part (April–September) of the reproductive cycle than the onset and maturation periods, so oocyte growth rates could not be measured continuously. Average oocyte diameter data indicated little size increase between February and June (~4 μm month−1), with rapid vitellogenesis from June to October (~14 μm month−1) and slower growth toward the end of the cycle from October to February (~9 μm month−1) (Fig. 4). The largest average oocyte diameter from a single colony was 133.7 μm (SD = 19.31) (February 9, 2006), and the largest oocyte found was 180 μm (February 14, 2006). Monthly size–frequency histograms (Fig. 5) provided a detailed presentation of the variability in oocyte development over time, particularly around the spawning period. March samples displayed the most variable oocyte size range as they contained a combination of mature oocytes from the current gametogenic cycle and developing oocytes from the next cohort. These frequency distributions indicate protracted spawning periods (both within each individual and for the population), rather than highly synchronous spawning where there would be a much smaller range of oocyte size classes, especially in the more mature stages.

Mean oocyte diameters of Lophelia pertusa from different sampling periods, with samples combined across years, showing the progression of the gametogenic cycle from oogonia (late Jan–early Mar) to pre-vitellogenic (Apr–Jun), vitellogenic (Aug–Dec) and mature vitellogenic (late Jan–early March) oocytes. Error bars represent one standard deviation from the mean. Numbers above the bars = number of females. Week # represents 7 day increments from January 1st


a–e Size–frequency distributions of Lophelia pertusa oocytes by sample period: a Mature vitellogenic oocytes of Lophelia pertusa and the following cohort of oogonia and early pre-vitellogenic oocytes occurred simultaneously, indicating overlapping gametogenic cycles; (f, g) April and June samples consisted primarily of pre-vitellogenic oocytes; (h–j) Between June and August, oocytes transitioned from pre-vitellogenic to vitellogenic and increased in size steadily as they matured. Error bars represent one standard deviation from the mean percent number of oocytes in each size category
Male gametogenesis
Between 2002 and 2008, male samples were collected for each month except April and July. Early spermatogenesis (stage I) was not observed in our samples; from May to November, the spermatocysts were in the maturation phase (stage II) with darkly stained spermatocytes and no spermatozoa present (Fig. 6a, b). Waller and Tyler (2005) reported Stage II as large open lumens; however, the spermatocysts from our samples were almost filled with spermatocytes, even in the earlier part of the cycle (May and June). From December to March, the spermatocysts were mature (Stage III) and filled with spermatozoa (Fig. 6c, d). As spermatogenesis progressed, the spermatocysts increased in size, often completely filling the mesoglea.

a–d Spermatocysts in the mesenteries of male Lophelia pertusa colonies: a early Stage II spermatocysts mostly filled with spermatocytes from samples collected in June. b late Stage II spermatocysts, completely filled with dense spermatocytes from September samples. c Samples collected in December showing early Stage III spermatocysts with spermatocytes and some spermatozoa. d February samples with mature spermatocysts showing primarily spermatozoa with some spermatocytes. Sc spermatocyte, sz spermatozoa. Scale bars 100 μm
Twelve of the samples collected during the spawning period could not be assigned a gender since there were no visible gametes. Females examined during this period either had remnant mature oocytes or oogonia; therefore, these 12 samples may have been post-spawning males. If so, this indicates that, unlike the females, male L. pertusa colonies do not have overlapping gametogenic cycles.
Discussion
This study describes a more complete gametogenic cycle of L. pertusa than previous work, with samples that encompass both initial gamete production and the spawning period. These data clearly show that the Norwegian fjord populations have overlapping gametogenic cycles. In contrast, Waller and Tyler (2005) suggested that the NE Atlantic (785–980 m) populations had a resting phase after the spawning period, with initiation of gametogenesis in late summer to coincide with food influx to the benthos (late June–August) from the spring phytoplankton bloom. Our data show that oogenesis not only began earlier in the Trondheim Fjord than in the NE Atlantic populations, but that successive cycles overlapped by one to 2 months. Oocytes from NE Atlantic populations of L. pertusa (Waller and Tyler 2005) in August were smaller than those collected from the Trondheim Fjord (means = 41.3 vs. 57.8 μm, respectively), but by October the oocyte sizes were similar (88.7 vs. 79.8 μm, respectively) between areas. The rapid oocyte growth (vitellogenesis) that we measured between summer and late fall was therefore even more pronounced in the NE Atlantic populations.
In summary, there are differences in female gametogenic cycles between the NE Atlantic offshore and the fjord populations of L. pertusa, including the presence/absence of a resting phase between cycles and the rate of vitellogenesis. Maximum oocyte diameter and the timing and duration of spawning cannot be compared as these data were not available for the offshore populations. The reasons for these differences are unclear, but could be linked to external environmental variables (temperature, food supply etc.) or differences in endogenous rhythms.
Male L. pertusa colonies, unlike the females, showed no indication of overlapping gametogenic cycles; however, it is not uncommon for anthozoan gametogenic stages to vary between males and females. Energetic investment in sperm is considerably less than for eggs, and sperm production and maturation is more rapid than egg development (Goffredo et al. 2002; Baillon et al. 2011).
There is ample evidence for exogenous control of reproduction in a variety of marine invertebrates, both in terms of both proximal cues (immediate) and ultimate drivers (evolutionary) (Giese and Pearse 1974, Tyler et al. 1994). These controlling factors include temperature, salinity, food supply, lunar phase, tidal cycles and daily light/dark cycles, solar insolation, rainfall patterns, and periods of calm that enhance fertilization (Himmelman 1980; Babcock et al. 1986; Yankson 1986; Sasaki and Shepherd 1995; Hardege and Bentley 1997; Mangubhai and Harrison 2008; Mercier et al. 2011, Harrison 2011). Seasonal change in sea temperature has been cited frequently as an important environmental factor that controls gametogenic cycles and planular release in shallow-water scleractinian corals (Szmant-Froelich et al. 1980; Kojis and Quinn 1981; Tranter et al. 1982; Fadlallah 1985; Stoddart and Black 1985; Babcock et al. 1986; Harrison and Wallace 1990). However, how environmental factors affect reproduction in marine organisms is still poorly understood and relies on correlations derived from field observations. Controlled experiments are needed to determine the extent to which environmental factors control scleractinian reproduction (Harrison 2011). This body of research has also focused primarily on shallow water, most likely for logistical reasons, rather than deep-sea ecosystems, so information on the latter is lacking.
The factors that drive reproductive periodicity in the deep sea are even less clear than for shallower depths. Synchronized seasonal or periodic reproductive cycles are quite common among invertebrate species that live shallower than 3,000 m (these species have a high degree of synchrony within species with short spawning periods), whereas those that live deeper tend to reproduce continuously (Gooday 2002). Temperatures are much less variable than in shallow waters, and potential reproductive cues linked to lunar, tidal, or diurnal cycles also attenuate with depth. Only one study has systematically investigated the relationships between lunar cycles and reproduction in deep-sea benthic invertebrates, observing that spawning and larval release in six species of echinoderms (living between 100 and 1,400 m) showed significant increase in activity around the new and full moons (Mercier et al. 2011). The authors suggested the lunar phase was a proxy for some other environmental factor such as cyclic currents or deposition of particulate matter (food) to the seafloor. Environmental data for deep-sea coral ecosystems are limited, and these topographic features often show dynamic environmental conditions on various temporal and spatial scales (Duineveld et al. 2007; White 2007; Davies et al. 2009; Roberts et al. 2009). It is possible that cold-water corals are exposed to environmental fluctuations of predictable timing, duration, and intensity, and one or more of these may influence various aspects of their reproduction.
In the deep sea, long-term environmental data are limited to a few locations where instruments have been deployed, and studies on reproductive cycles are also rare, limiting our ability to generate strong correlations. In the NE Atlantic from Norway to the Porcupine Seabight, temperatures at cold-water coral depths ranged from 6.5 to 9.9 °C (Dullo et al. 2008), but with no information on temporal variability. Measurements over a 1-year period near L. pertusa banks on the Rockall Trough (NE Atlantic) revealed an average bottom temperature of 8–9 °C, with maximum daily fluctuations of 2.6 °C (Mienis et al. 2007). These temperature fluctuations correlated with tidal cycles, but there were no consistent seasonal signals that might drive reproductive cycles. Water temperatures in the Trondheim Fjord (near our sample sites) showed virtually no temporal variation (7.5–8 °C) at 100 m depth (Børsheim et al. 1999). In the western north Atlantic, extensive L. pertusa reefs occur off the southeastern USA (Ross and Nizinski 2007) and Gulf of Mexico (Brooke and Schroeder 2007). Limited environmental data from these sites (Brooks and Bane 1983; Bane et al. 2001; Continental Shelf Associates 2007; Mienis et al. 2012) show no clear connection between temperature and the timing of L. pertusa reproductive cycles.
The timing of food supply may influence various aspects of deep-sea invertebrate reproduction (Gage and Tyler 1991; Tyler et al. 1992, 1993; Campos-Creasey et al. 1994; Eckelbarger and Watling 1995; Gooday 2002; Young 2003), including initiation and timing of gametogenesis and vitellogenesis, timing of spawning or larval release, and timing of larval recruitment. Surface productivity blooms occur on a predictable seasonal schedule in many locations (Deuser 1986), with some inter-annual variation in timing and intensity. Delivery of labile carbon from the surface to the seafloor can occur in a few days at depths above 1,000 m in the NE Atlantic (Gage and Tyler 1991), creating seasonal food pulses to the benthos (Billett et al. 1983; Lampitt 1985; Rice et al. 1986; Thiel et al. 1989; Gooday 2002) that may also vary in timing, quantity, and composition (Deuser 1986; Newton et al. 1994; Wong et al. 1999; Takahashi et al. 2000, Lampitt et al. 2001). Vitellogenesis is energetically expensive as oocytes have high lipid (20–50 % of the egg composition) and protein content (Hoegh-Guldberg and Emlet 1997; Sewell and Manahan 2001, Moran and Manahan 2004; Byrne et al. 2008). Deep-sea species (especially those with large eggs or high fecundity) may therefore coordinate vitellogenesis with periods of high food delivery to the benthos (Eckelbarger and Watling 1995). For example, seasonal influx of organic material to the bottom was implicated in the initiation of gametogenesis or vitellogeneis in some cold-water scleractinians (Waller and Tyler 2005; Mercier et al. 2011) and octocorals (Sun et al. 2010; Mercier and Hamel 2011).
Food supply may influence different aspects of reproduction, including timing of spawning and larval release. Marine invertebrate species that produce large lipid-rich eggs generally produce lecithotrophic larvae, which do not need an exogenous food supply. Species that produce small eggs have a lower energy reserve and usually produce small planktotrophic larvae (Jaeckle 1995; Sewell and Manahan 2001; Prowse et al. 2008). Larval development mode has not yet been determined for L. pertusa, but this species produces small eggs (maximum = 140 μm) (Waller and Tyler 2005). To facilitate larval growth and development, species with planktotrophic larvae may therefore time their spawning to coincide with high food levels in the water column (Tyler et al. 1982). However, timing of larval recruitment with seasonal food pulses has only been observed in opportunistic macrofaunal species that have rapid reproductive cycles (Blake and Watling 1994; Gooday et al. 1996; Snelgrove et al. 1996). There are no documented examples of this response in megafauna, which produce eggs more slowly than the smaller macrofauna. Slower egg production disconnects food influx and the presence of mature larvae that are ready to recruit (Eckelbarger and Watling 1995).
Information on influx of food to the deep sea is limited and is restricted to a few locations. In the temperate North Atlantic, the spring phytoplankton bloom is usually well defined and results in a strong flux of phytodetritus that accumulates on the seafloor at bathyal and abyssal depths in the late spring and early summer (Lampitt 1985; Rice et al. 1994; Lampitt and Antia 1997; Longhurst 1998; Lampitt et al. 2001, Duineveld et al. 2007). Norwegian fjords also exhibit a spring (March/April to May/June) phytoplankton bloom, much of which sinks to the bottom (Burrell 1988; Wassmann et al. 1996), in a similar pattern to the offshore NE Atlantic. The Trondheim Fjord exhibits a variable seasonal pattern in dissolved organic carbon (DOC) between years, but in general DOC is highest at depth during the summer months (Børsheim et al. 1999). For more northerly Norwegian fjords, the deposition of particulate organic carbon (POC) peaked in April and May, coincident with the spring bloom; however, there were two additional pulses observed later in the year (June and September) in response to freshwater influx (Børsheim et al. 1999). These signatures are complicated by inter-annual variation in freshwater runoff, benthic re-suspension by currents and horizontal advection of water masses between the fjords and offshore areas. In fjord systems, the relationship between surface phytoplankton blooms and delivery of organic material to the benthos is more complex than for many open ocean or continental margin systems (Børsheim et al. 1999). These differences in food dynamics between open ocean and fjord systems may account for the observed differences in L. pertusa gametogenesis, but more data are needed before such correlations can be tested.
There is an apparent difference of several months in the onset of gametogenesis between our study in Norway (January) and that estimated by Waller and Tyler (2005) from the NE Atlantic (‘late summer’). Endogenous or genetic factors cannot explain differences between conspecific populations that are in the same region and are genetically similar (Morrison et al. 2011). Exogenous factors seem more likely to account for the observed differences in the timing and duration of L. pertusa gametogenic cycles; however, there are not enough environmental data from the two study areas to draw conclusions on what these might be. Both studies showed a similar rapid increase in oocyte diameters from early summer through December, which coincides with the seasonal influx of particulate organic carbon from the spring and (smaller) fall phytoplankton blooms. The largest phytodetrital pulse disappears before the completion of the reproductive cycle (in both fjord and offshore locations), so either the corals have the capability of long-term energy storage or they may use a different food source to complete vitellogenesis. The large polyps of L. pertusa are well equipped with batteries of nematocysts that enable them to capture large, active planktonic food items (Freiwald 2002). Although there are few controlled feeding experiments or in situ observations of L. pertusa feeding, isotope studies indicate that this species has a mixed diet of zooplankton and phytodetrital material (Duineveld et al. 2004; Kiriakoulakis et al. 2005) and in some cases phytoplankton material (Duineveld et al. 2012). Consideration of zooplankton dynamics at depth adds a layer of complexity to the role of food as a driver of reproductive activity. Zooplankton populations increase in response to phytoplankton blooms, and many zooplankton species undergo both diurnal as well as seasonal vertical migrations that reach the deep coral communities, especially in the shallow fjord habitats. Benthic zooplankton also exhibit diurnal vertical migrations in the vicinity of L. pertusa mounds, putting them within reach of the coral polyps (Davies et al. 2010; Mienis et al. 2012). In the Trondheim Fjord, the annual migration cycle of the abundant copepod Calanus finnmarchicus corresponds to the gametogenic cycle of L. pertusa, and it is possible that C. finnmarchicus constitutes an important food source for the corals at the onset of gametogenesis as well as throughout gonadal development. The dynamics of C. finnmarchicus in the north sea are very complex, and there may be differences in the amount and timing of the vertical migration of C. finnmarchicus in the offshore NE Atlantic (Heath et al. 2004) and the fjord systems that could account for the difference in reproductive timing between this study and Waller and Tyler (2005).
Reproductive cycles of L. pertusa from the southeastern US (S. Brooke, unpubl. data) and the Gulf of Mexico (Brooke et al. 2007) are offset by several months from those populations in the NE Atlantic. Spawning appears to occur in late September to early November in the southeastern US and Gulf of Mexico, rather than late January to early March in the NE Atlantic. The discussion above indicates that seasonal organic input may influence the reproductive cycles of L. pertusa; however, the exact mechanisms by which such environmental factors stimulate physiological responses are unknown (Roberts et al. 2009). In addition, while exogenous factors may influence invertebrate reproductive biology, there are also genetically constrained endogenous factors that may play a role in reproductive processes (Eckelbarger and Watling 1995). Large regional differences in reproductive timing within a single species, as occurs with L. pertusa, provide a rare opportunity to study exogenous factors that may influence the drivers of reproduction and identify which factors are constrained endogenously.
When corals are under stress, they may allocate energy away from reproduction into metabolic maintenance, tissue repair, or growth (Brown and Howard 1985; Richmond 1987; Ward 1995). Although this phenomenon has not been directly studied in cold-water corals, Waller and Tyler (2005) noted that samples of L. pertusa from their heavily trawled sites on the Darwin Mounds did not contain any reproductive material, whereas samples from the less impacted sites were packed with gametes. Population genetic analysis of L. pertusa from the heavily trawled Darwin Mounds also indicated very low levels of sexual reproduction (Le Goff-Vitry et al. 2004). Changes in reproductive output would not be immediately obvious, but could have a long-term influence on ecosystem resilience and possibly genetic diversity. This has important implications for deep coral reefs impacted by destructive human activities and future widespread changes in ocean conditions resulting from global climate change.
References
Asche F, Guttormsen AG, Tveteras R (1999) Environmental problems, productivity and innovations in Norwegian salmon aquaculture. Aquac Econ Manag 3:19–29
Babcock RC, Bull GD, Harrison PL, Heyward AJ, Oliver JK, Wallace CC, Wills BL (1986) Synchronous spawnings of 105 scleractinian coral species on the Great Barrier Reef. Mar Biol 90:379–394
Baillon S, Hamel J-F, Mercier A (2011) Comparative study of reproductive synchrony at various scales in deep-sea echinoderms. Deep-sea Res I 58:260–272
Baird AH, Guest JR, Willis BL (2009) Systematic and biogeographical patterns in the reproductive biology of scleractinian corals. Ann Rev Ecol Evol Syst 40:551–571
Bane JM Jr, Attkinson LP, Brooks DA (2001) Gulf Stream physical oceanography at the Charleston bump: deflection, bimodality, meanders, and upwelling. In: Sedberry GR (ed) Island in the Stream: oceanography and fisheries of the Charleston bump. American fisheries society symposium 25, Bethesda, pp 25–36
Billett DSM, Lampitt RS, Rice AL, Mantoura RFC (1983) Seasonal sedimentation of phytoplankton to the deep-sea benthos. Nature 302:520–522
Blake JA, Watling L (1994) Life history studies of deep-sea benthic infauna: Polychaeta, Aplacophora, and Cumacea from the continental slope off Massachusetts. In: Young CM, Eckelbarger KJ (eds) Reproduction, larval biology and recruitment of the deep-sea benthos. Columbia University Press, New York, pp 243–260
Børsheim KY, Myklestad SM, Sneli JA (1999) Monthly profiles of DOC, mono- and polysaccharides at two locations in the Trondheimsfjord (Norway) during two years. Mar Chem 63:255–272
Brooke S, Schroeder WW (2007) Ch 7. State of deep coral ecosystems in the Gulf of Mexico region: Texas to the Florida straits. In: The state of deep coral ecosystems of the United States. Lumsden SE, Hourigan TF, Bruckner AW (eds) NOAA Tech Memo NOS-CRCP-3, Silver Spring, pp 271–306
Brooke S, Young CM (2003) Reproductive ecology of a deep-water scleractinian coral, Oculina varicosa. Cont Shelf Res 23:847–858
Brooke S, Young CM (2005) Embryogenesis and larval biology of the ahermatypic scleractinian Oculina varicosa. Mar Biol 146:665–675
Brooke S, Young CM, Holmes M (2007) Ch 6. Biological characterization and studies. In: Continental shelf associates. Characterization of Northern Gulf of Mexico deepwater hard bottom communities with emphasis on Lophelia coral. Final Report to US Dept Interior, Minerals Management Service, Gulf of Mexico Region. OCS Study 2007-044, pp 119–147
Brooke S, Holmes M, Young CM (2009) Effects of sediment on two morphotypes of Lophelia pertusa from the Gulf of Mexico. Mar Ecol Prog Ser 390:137–144
Brooks DA, Bane JM Jr (1983) Gulf stream meanders off North Carolina during winter and summer 1979. J Geophys Res 88:4633–4650
Brown BE, Howard LS (1985) Assessing the effects of ‘stress’ on reef corals. Adv Mar Biol 22:1–63
Burgess S, Babcock RC (2005) Reproductive ecology of three reef-forming, deep-sea corals in the New Zealand region. In: Freiwald A, Roberts JM (eds) Cold-water corals and ecosystems. Springer, New York, pp 701–713
Burrell DC (1988) Carbon flow in fjords. Oceanogr. Mar Biol Rev 26:143–226
Byrne M, Prowse TAA, Sewell MA, Dworjanyn S, Williamson JE, Vaitilingon D (2008) Maternal provisioning for larvae and larval provisioning for juveniles in the toxopneustid sea urchin Tripneustes gratilla. Mar Biol 155(5):473–482
Cairns SD (1979) The deep-water scleractinia of the Caribbean Sea and adjacent waters. Studies on the fauna of Curacao and other Caribbean Islands, vol 57(180), p 341
Cairns SD (1995) The marine fauna of New Zealand: Scleractinia (Cnidaria: Anthozoa). NZ Oceanogr Inst Mems 103:210
Campos-Creasey LS, Tyler PA, Gage JD, John AWG (1994) Evidence for coupling the vertical flux of phytodetritus to the diet and seasonal life history of the deep-sea echinoid Echinus affinis. Deep-Sea Res 41:369–388
Clark MR, Tittensor DP (2010) An index to assess the risk to stony corals from bottom trawling to seamounts. Mar Ecol 31(Suppl 1):200–211
Continental Shelf Associates (2007) Characterization of northern Gulf of Mexico deepwater hard bottom communities with emphasis on Lophelia coral. US Dept Interior, Minerals Management Service, Gulf of Mexico OCS Region, New Orleans, LA OCS Study MMS 2007–044, p 169+ appendices
Davies AJ, Duineveld GCA, Lavaleye MSS, Bergman MJN, van Haren H, Roberts JM (2009) Downwelling and deep-water bottom currents as food supply mechanisms to the cold-water coral Lophelia pertusa (Scleractinia) at the Mingulay Reef complex. Limnol Oceanogr 54:620–629
Davies AJ, Duineveld GCA, van Weering TCE, Mienis F, Quattrini AM, Seim HE, Bane JM, Ross SW (2010) Short-term environmental variability in cold-water coral habitat at Viosca Knoll, Gulf of Mexico. Deep-Sea Res PTI 57:199–212
De Juan S, Lleonart J (2010) A conceptual framework for the protection of vulnerable habitats impacted by fishing activities in the Mediterranean high seas. Ocean Coast Manag J 53:717–723
de Putron SJ, McCorkle DC, Cohen AL, Dillon AB (2011) The impact of seawater saturation state and bicarbonate ion concentration on calcification by new recruits of two Atlantic corals. Coral Reefs 30:321–328
Deuser WG (1986) Seasonal and interannual variations in deep water particle fluxes in the Sargasso Sea and their relation to surface hydrography. Deep-Sea Res 33:225–246
Duineveld GCA, Lavaleye MSS, Berghuis EM (2004) Particle flux and food supply to a seamount cold-water coral community (Galicia Bank, NW Spain). Mar Ecol Prog Ser 277:13–23
Duineveld GCA, Lavaleye MSS, Bergman MJN, Stigter H, Mienis F (2007) Trophic structure of a cold water coral mound community (Rockall Bank, NE Atlantic) in relation to the near bottom particle supply and current regime. Bull Mar Sci 81:449–467
Duineveld GCA, Jeffreys RM, Lavaleye MSS, Davies AJ, Bergman MJN, Watmough T, Witbaard R (2012) Spatial and tidal variation in food supply to shallow cold-water coral reefs of the Mingulay Reef complex (Outer Hebrides, Scotland). MEPS 444:97–115
Dullo W-C, Flögel S, Rüggeberg A (2008) Cold-water coral growth in relation to the hydrography of the Celtic and Nordic Eurpoean continental margin. Mar Ecol Prog Ser 371:165–176
Eckelbarger KJ, Watling L (1995) Role of phylogenetic constraints in determining reproductive patterns in deep-sea invertebrates. Invertebr Biol 114:256–269
Fadlallah YH (1983) Sexual reproduction, development and larval biology of scleractinian corals. A review. Coral Reefs 2:129–150
Fadlallah YH (1985) Reproduction in the coral Pocillopora verrucosa on the reefs adjacent to the industrial city of Yanbu, Red Sea, Saudi Arabia. In: Proceedings of the 5th international coral reef congress, vol 4, Tahiti, pp 313-318
Flint H, Waller R, Tyler P (2007) Reproductive ecology of Fungiacyathus marenzelleri from 4100 m depth in the Northeast Pacific Ocean. Mar Biol 151:843–849
Fosså JH, Mortensen PB, Furevik DM (2002) The deep-water coral Lophelia pertusa in Norwegian waters: distribution and fishery impacts. Hydrobiologia 471:1–12
Freiwald A, Roberts JM (eds) (2005) Cold-water corals and ecosystems. Springer, Berlin
Freiwald A, Hűhnerbach V, Lindberg B, Wilson JB, Campbell J (2002) The Sula Reef complex, Norwegian Shelf. Facies 47:179–200
Freiwald A (2002) Reef-forming cold-water corals. In: Wefer G, Billett D, Hebbeln D et al (eds) Ocean margin systems. Springer, Berlin, pp 365–385
Gage JD, Tyler PA (1991) Deep-sea biology: a natural history of organisms at the deep-sea floor. Cambridge University Press, Cambridge, p 504
Giese AC, Pearse JS (1974) Introduction: general principles. In: Giese AC, Pearse JS (eds) Reproductive ecology of marine invertebrates. Wiley, New York
Goffredo S, Arnone S, Zaccanti F (2002) Sexual reproduction in the Mediterranean solitary coral Balanophyllia europaea (Scleractinia, Dendrophylliidae). Mar Ecol Prog Ser 229:83–94
Gooday AJ (2002) Biological responses to seasonally varying fluxes of organic matter to the ocean floor: a review. J Oceanogr 58:305–332
Gooday AJ, Pfannkuche O, Lambshead PJD (1996) An apparent lack of response by metazoan meiofauna to phytodetritus deposition in the bathyal northeast Atlantic. J Mar Biol Assoc UK 16:297–310
Hardege JD, Bentley MG (1997) Spawning synchrony in Arenicola marina: evidence for sex pheromonal control. Proc R Soc Lond Ser B 264(1384):1041–1047
Harrison PL (2011) Sexual reproduction of scleractinian corals. In: Dubinsky Z, Stambler N (eds) Coral reefs: an ecosystem in transition. Springer, Berlin, pp 59–85
Harrison PL, Wallace CC (1990) Reproduction, dispersal and recruitment of Scleractinian corals. In: Dubinsky Z (ed) Ecosystems of the World No. 25. Elsevier, New York, pp 133–207
Havenhand JN (1993) Egg to juvenile period, generation time, and the evolution of larval type in marine invertebrates. Mar Ecol Prog Ser 97:247–260
Heath MR, Boyle PR, Gislason A, Gurney WSC, Hay SJ, Head EJH, Holmes S, Ingvarsdottir A, Jonasdottir SH, Lindeque P, Pollard RT, Rasmussen J, Richards K, Richardson K, Smerdon G, Speirs D (2004) Comparative ecology of over-wintering Calanus finmarchicus in the northern North Atlantic, and implications for life-cycle patterns. ICES J Mar Sci 61:69
Highsmith RC (1982) Reproduction by fragmentation in corals. Mar Ecol Prog Ser 7:207–226
Himmelman JH (1980) Synchronization of spawning in marine invertebrates. In: Clark WH Jr, Adams TS (eds) Advances in invertebrate reproduction. Elsevier, New York, pp 3–19
Hoegh-Guldberg O, Emlet RB (1997) Energy use during the development of a lecithotrophic and a planktotrophic echinoid. Biol Bull 192:27–40
Hovland M (2008) Deep-water coral reefs: unique biodiversity hotspots. Springer-Praxis Books, Springer, Netherlands, pp 278
Jacobson P (1983) Physical oceanography of the Trondheimsfjord. Geophys Astrophys Fluid Dyn 26:3–26
Jaeckle WB (1995) Variation in the size, energy content and biochemical composition of invertebrate eggs: correlates to the mode of larval development. In: McEdward L (ed) Ecology of Marine Invertebrate Larvae. CRC Press, Boca Raton, pp 49–77
Johannessen T, Dahl E (1996) Declines in oxygen concentrations along the Norwegian Skagerrak coast, 1927–1993: a signal of ecosystem changes due to eutrophication? Limnol Oceanogr 41:766–778
Kiriakoulakis K, Fisher E, Wolff GA, Freiwald A, Grehan A, Roberts JM (2005) Lipids and nitrogen isotopes of two deep-water corals from the North-East Atlantic: initial results and implications for their nutrition. In: Freiwald A, Roberts JM (eds) Cold-water corals and ecosystems. Springer, New York, pp 715–729
Kojis BL, Quinn NJ (1981) Aspects of sexual reproduction and larval development in the shallow water hermatypic coral Goniastera australensis (Edwards and Haime, 1857). Bull Mar Sci 31:558–573
Koslow JA, Gowlett-Jones K (1998) The seamount fauna off southern Tasmania: benthic communities, their conservation and impacts of trawling. Final Report to Environment Australia and Fisheries Research Development Corporation. Australia, p 104
Krupp DA (1983) Sexual reproduction and early development of the solitary coral Fungia scutaria (Anthozoa, scleractinia). Coral Reefs 2(3):159–164
Lampitt RS (1985) Evidence for the seasonal deposition of detritus to the deep-sea floor and its subsequent resuspension. Deep-Sea Res 32A:885–897
Lampitt RS, Antia AN (1997) Particle flux in deep seas: regional characteristics and temporal variability. Deep-Sea Res PTI 44:1377–1403
Lampitt RS, Brett BJ, Kiriakoulis K, Popova EE, Ragueneau O, Vangriesheim A, Wolff GA (2001) Material supply to the abyssal seafloor in the northeast Atlantic. Prog Oceanogr 50:27–63
Larsson AI, Purser A (2011) Sedimentation on the cold-water coral Lophelia pertusa: cleaning efficiency from natural sediments and drill cuttings. Mar Pollut Bull 62:1159–1168
Le Goff-Vitry MC, Pybus OG, Rogers AD (2004) Genetic structure of the deep-sea coral Lophelia pertusa in the northeast Atlantic revealed by microsatellites and internal transcribed spacer sequences. Mol Ecol 13:537–549
Longhurst A (1998) Ecological geography of the sea. Academic Press, San Diego, p 398
Loya Y, Rinkevich B (1979) Abortion effect in corals induced by oil pollution. Mar Ecol Prog Ser 1:77–80
Mangubhai S, Harrison PL (2008) Asynchronous coral spawning patterns on equatorial reefs in Kenya. Mar Ecol Prog Ser 360:85–96
McFadden CS, Hutchinson MB (2004) Molecular evidence for the hybrid origin of species in the soft coral genus Alcyonium (Cnidaria: Anthozoa: Octocorallia). Mol Ecol 13:1495–1505
McFadden CS, Donahue R, Hadland BK, Weston R (2001) A molecular phylogenetic analysis of reproductive trait evolution in the soft coral genus Alcyonium. Evolution 55:54–67
Mercier A, Hamel J-F (2011) Reproductive periodicity, spawning and development of the deep-sea scleractinian coral Flabellum angulare. Mar Biol 158:371–380
Mercier A, Sun Z, Hamel J-F (2011) Reproductive periodicity, spawning and development of the deep-sea scleractinian coral Flabellum angular. Mar Biol 158:371–380
Mienis F, de Stigter HC, White M, Duineveld G, de Haas H, van Weering TCE (2007) Hydrodynamic controls on cold-water coral growth and carbonate-mound development at the SW and SE Rockall Trough margin, NE Atlantic Ocean. Deep-Sea Res PTI 54:1655–1674
Mienis F, Duineveld GCA, Davies AJ, Ross SW, Seim H, Bane J, van Weering TCE (2012) The influence of near-bed hydrodynamic conditions on cold-water corals in the Viosca Knoll area, Gulf of Mexico. Deep-Sea Res PTI 60:32–45
Moran AL, Manahan DT (2004) Physiological recovery from prolonged ‘starvation’ in larvae of the Pacific oyster Crassostrea gigas. J Exp Mar Biol Ecol 306:17–36
Morrison CL, Ross SW, Nizinski MS, Brooke S, Järnegren J, Waller RG, Johnson RL, King TL (2011) Genetic discontinuity among regional populations of Lophelia pertusa in the North Atlantic Ocean. Conserv Genet 12:713–729
Mortensen PB, Hovland M, Brattegard T, Farestveit R (1995) Deep water bioherms of the scleractinian coral Lophelia pertusa (L.) at 64°N on the Norwegian shelf: structure and associated megafauna. Sarsia 80:145–158
Newton PP, Lampitt RS, Jickells TD, King P, Boutle C (1994) Temporal and spatial variability of biogenic particle fluxes during JGOFS Northeastern Atlantic process studies at 47°N, 20°W. Deep Sea Res PTI 41:1617–1642
Pandolfi JM, Connolly SR, Marshall DJ, Cohen AL (2011) Projecting coral reef futures under global warming and ocean acidification. Science 333:418
Prowse TAA, Sewell MA, Byrne M (2008) Fuels for development: evolution of maternal provisioning in asterinid sea stars. Mar Biol 153:337–349
Rice AL, Billett DS, Fry J, John AWG, Lampitt RS, Mantoura RFC, Morris RJ (1986) Seasonal deposition of phytodetritus to the deep-sea floor. Proc Roy Soc Edinb 88B:265–279
Rice AL, Thurston MH, Bett BJ (1994) The IOS DEEPSEAS programme: introduction and photographic evidence for presence and absence of seasonal input of phytodetritus at contrasting abyssal sites in the northeastern Atlantic. Deep Sea Res PTI 41:1305–1320
Richmond RH (1987) Energetic relationships and biogeographical differences among fecundity, growth and reproduction in the reef coral Pocillopora damicornis. Bull Mar Sci 41:594–604
Rinkevich B, Loya Y (1979) Laboratory experiments on the effects of crude oil on the Red Sea coral Stylophora pistillata. Mar Pollut Bull 10:328–330
Rinkevich B, Loya Y (1987) Variability in the pattern of sexual reproduction of the coral Stylophora pistillata at Eilat, Red Sea: a long-term study. Biol Bull 173:335–344
Roberts JM, Wheeler AJ, Freiwald A, Cairns SD (2009) Coldwater corals: the biology and geology of deep-sea coral habitats. Cambridge University Press, Cambridge, p 352
Rogers AD (1999) The biology of Lophelia pertusa (Linnaeus 1758) and other deep-water reef-forming corals and impacts from human activities. Int Rev Hydrobiol 84:315–406
Ross SW, Nizinski MS (2007) State of deep coral ecosystems in the U.S. southeast region: Cape Hatteras to southeastern Florida, pp 233–270, Ch 6 In: Lumsden SE, Hourigan TF, Bruckner AW, Dorr G (eds) The state of deep coral ecosystems of the United States. NOAA Tech Memo CRCP-3. Silver Spring, p 365
Sammarco PW (1982) Polyp bail-out: an escape response to environmental stress and a new means of reproduction in corals. Mar Ecol Prog Ser 10:57–65
Sasaki R, Shepherd SA (1995) Larval dispersal and recruitment of Haliotis discus hannai and Tegula sp. on Miyagi coasts. Jpn Mar Freshwat Res 46:519–529
Sewell MA, Manahan DT (2001) Echinoderm eggs; biochemistry and larval biology, pp 55–58 in echinoderms 2000: proceedings of the 10th international conference, Dunedin. CRC Press, Boca Raton
Smelror M (2000) Hvordan ble Trondheimsfjorden til? In: Sakshaug E, Sneli J-A (eds) Trondheimsfjorden. Tapir forlag, Trondheim, pp 19–37
Snelgrove PVR, Grassle JF, Petrecca RF (1996) Experimental evidence for ageing food patches as a factor contributing to high macrofaunal diversity. Limnol Oceanog 41:605–614
Stoddart JA (1983) Asexual production of planulae in the coral Pocillopora damicornis. Mar Biol 76:279–284
Stoddart JA, Black R (1985) Cycles of gametogenesis and planulation in the coral Pocillopora damicornis. Mar Ecol Progr Ser 23:153–164
Strathmann RR (1985) Feeding and nonfeeding larval development and life-history evolution in marine invertebrates. Annu Rev Ecol Syst 16:339–361
Strømgren T (1971) Vertical and horizontal distribution of Lophelia pertusa (Linné) in Trondheimsfjorden on the west coast of Norway. Kongel Norske Vidensk Selsk Skrifter 6:1–9
Sun Z, Hamel J-F, Mercier A (2010) Planulation periodicity, settlement preferences and growth of two deep-sea octocorals from the northwest Atlantic. Mar Ecol Prog Ser 410:71–87
Szmant-Froelich AM, Yevich P, Pilson MEQ (1980) Gametogenesis and early development of the temperate coral Astrangia danae (Anthozoa: Scleractinia). Biol Bull 158:25–269
Takahashi K, Fujitani N, Yanada M, Maita Y (2000) Long term biogenic particle fluxes in the Bering Sea and central subarctic Pacific Ocean 1990–1995. Deep Sea Res I 47:1723–1759
Thiel H, Pfannkuche O, Schriever G, Lochte K, Gooday AJ, Hemleben C, Mantoura RFC, Turley CM, Patching JW, Riemann F (1989) Phytodetritus on the deep-sea floor in a central oceanic region of the northeast Atlantic. Biol Oceanogr 6:203–239
Thorsnes T, Fosså JH, Christensen O (2004) Deep-water coral reefs. Acoustic recognition and geological setting. Hydro Int 8(3):26–29
Tranter PRG, Nicholson DN, Kitchington D (1982) A description of the spawning and post gastrula development of the cool temperate coral Caryophillia smithii (Stokes and Broderip). J Mar Biol Assoc UK 62:845–854
Tyler PA, Pain SL, Gage JD (1982) The reproductive biology of the deep-sea asteroid Bathybiaster vexillifer. J Mar Biol Assoc UK 62:57–69
Tyler PA, Harvey R, Giles LA, Gage JD (1992) Reproductive strategies and diet in deep-sea nuculanid protobranchs (Bivalvia: Nuculoidea) from the Rockall Trough. Mar Biol 114:571–580
Tyler PA, Gage JD, Paterson GJL, Rice AL (1993) Dietary constraints on reproductive periodicity in two sympatric deep-sea astropectinid seastars. Mar Biol 115:267–277
Tyler PA, Campos-Creasey LS, Giles LA (1994) Environmental control of quasi-continuous and seasonal reproduction in deep sea invertebrates. In: Young CM, Eckelbarger KJ (eds) Reproduction, larval biology and recruitment of the deep sea benthos. Columbia University Press, New York, pp 158–178
Waller RG (2005) Deep-water Scleractinia (Cnidaria: Anthozoa): current knowledge of reproductive processes. In: Freiwald A, Roberts JM (eds) Cold-water corals and ecosystems. Springer, New York, pp 691–700
Waller RG, Tyler PA (2005) The reproductive biology of two deep-water, reef-building scleractinians from the NE Atlantic Ocean. Coral Reefs 24:514–522
Waller RG, Tyler PA (2011) Reproductive patterns in two deep-water solitary corals from the north-east Atlantic- Flabellum alabastrum and F. angulare (Cnideria: Anthozoa: Scleractinia). J Mar Biol Assoc UK 91:669–675
Waller RG, Tyler PA, Gage JD (2002) The reproductive ecology of the deep-sea scleractinian coral Fungiacyathus marenzelleri (Vaughan, 1906) in the northeast Atlantic Ocean. Coral Reefs 21:325–331
Waller RG, Tyler PA, Gage JD (2005) Sexual reproduction in three hermaphroditic deep-sea Caryophyllia species (Anthozoa: Scleractinia) from the NE Atlantic Ocean. Coral Reefs 24:594–602
Ward S (1995) Two patterns of energy allocation for growth, reproduction and lipid storage in the scleractinian coral Pocillopora damicornis. Coral Reefs 14:87–90
Wassmann P, Svendsen H, Keck A, Reigstad M (1996) Selected aspects of the physical oceanographic and particle fluxes in fjords of northern Norway. J Mar Syst 8:53–71
White M (2007) Benthic dynamics at the carbonate mound regions of the Porcupine Seabight continental margin. Int J Earth Sc 96:1–9
Wong CS, Whitney FA, Crawford DW, Iseki K, Matear RJ, Johnson WJ, Page JS, Timothy (1999) Seasonal and interannual variability in particle fluxes of carbon nitrogen and silicon from time-series sediment traps at Ocean Station P, 1982–1983: relationship to changes in subarctic primary productivity. Deep Sea Res PTII 46:2735–2760
Yankson K (1986) Reproductive cycles of Cerastoderma glaucum (Bruguiere) and C. edule (L.) with special reference to the effects of the 1981–82 severe winter. J Mollus Stud 52:6–14
Young CM (2003) Reproduction, development and life history traits. Ch 12. In: Tyler PA (ed) Ecosystems of the deep oceans. Ecosystems of the World 28. Elsevier, Amsterdam, pp 381–426
Acknowledgments
This research was funded by VISTA under contract #6153. Our sincere thanks go to Dr Craig Young at the Oregon Institute of Marine Biology for the generous use of his facilities for histological processing, to the crew of the R/V Gunnerus for their help during sample collections and to the staff at NTNU for providing excellent facilities for keeping the corals.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by U. Sommer.
Rights and permissions
About this article
Cite this article
Brooke, S., Järnegren, J. Reproductive periodicity of the scleractinian coral Lophelia pertusa from the Trondheim Fjord, Norway. Mar Biol 160, 139–153 (2013). https://doi.org/10.1007/s00227-012-2071-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-012-2071-x