
Abstract
Many corals obtain their obligate intracellular dinoflagellate symbionts from the environment as larvae or juveniles. The process of symbiont acquisition remains largely unexplored, especially under stress. This study addressed both the ability of Fungia scutaria (Lamarck 1801) larvae to establish symbiosis with Symbiodinium sp. C1f while exposed to elevated temperature and the survivorship of aposymbiotic and newly symbiotic larvae under these conditions. Larvae were exposed to 27, 29, or 31°C for 1 h prior to infection, throughout a 3-h infection period, and up to 72 h following infection. Exposure to elevated temperatures impaired the ability of coral larvae to establish symbiosis and reduced larval survivorship. At 31°C, the presence of symbionts further reduced larval survivorship. As sea surface temperatures rise, coral larvae exposed to elevated temperatures during symbiosis onset will likely be negatively impacted, which in turn could affect the establishment of future generations of corals.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Scleractinian corals and their unicellular dinoflagellate endosymbionts of the genus Symbiodinium form the foundation of coral reef ecosystems (Dubinsky 1990). Both partners have developed numerous cellular and physiological adaptations that maximize mutual benefits and support their association (Furla et al. 2005). Host corals gain organic nutrients that are produced by the dinoflagellates during photosynthesis (Muscatine and Hand 1958). Symbiotic dinoflagellates benefit from nutrients such as dissolved inorganic carbon and nitrogen excreted by the coral (Yellowlees et al. 2008). For many symbiotic associations, it remains uncertain how environmental variation, genotype, and condition of the partners influence the cost–benefit balance for the individuals in the association (Stachowicz 2001; Douglas 2008; Oliver et al. 2009). The coral–dinoflagellate symbiosis, which together with associated protists and microbes is known as the coral holobiont, is one symbiotic system that is well suited to explore how the partnership is affected by changes in environmental conditions.
The majority of scleractinian coral species broadcast spawn gametes free of dinoflagellates (Baird et al. 2009). For these species, horizontally transmitted symbionts are taken up from the environment by each host generation (Fadlallah 1983; Harrison and Wallace 1990). Horizontal transmission allows for flexibility in symbiotic associations, but environmental factors can affect symbiont availability and mechanisms controlling specificity are poorly known (Weis 2010). Symbiosis is typically established when dinoflagellates enter through the mouth of a coral larva or polyp and are subsequently phagocytosed by endodermal cells lining the gastrovascular cavity (Colley and Trench 1983; Fitt and Trench 1983; Schwarz et al. 1999), although direct ectodermal incorporation has also been reported (Marlow and Martindale 2007). Dinoflagellates ultimately reside in vacuoles of host gastrodermal cells (Wakefield et al. 2000).
Potential costs of the partnership are regulated through several mechanisms. Under stable circumstances, both partners manage dangerous reactive oxygen species (ROS) produced as a by-product of photosynthesis by making use of antioxidant enzymes such as superoxide dismutase (SOD) and catalase (Richier et al. 2005; Lesser 2006). When stressful circumstances such as increased temperature lead to excess ROS, the stable association can rapidly deteriorate as ROS-scavenging enzymes fail to keep up (Lesser 2006). This oxidative stress can eventually lead to coral bleaching, or collapse of the symbiosis, through one of several cellular mechanisms (Douglas 2003; Weis 2008).
The effects of global climate change, including elevated sea surface temperatures resulting from increased atmospheric levels of CO2 (Hoegh-Guldberg et al. 2007; Carpenter et al. 2008), threaten the health of corals, many of which live at temperatures near the upper limits of their thermal tolerance (Glynn 1993; Coles and Brown 2003). Bleaching in adult corals can be induced by short-term exposure (1–2 days) to temperatures 3–4°C above the normal summer ambient temperature or by long-term exposure (several weeks) to temperatures 1–2°C above ambient (Jokiel and Coles 1977, 1990). Since average sea surface temperatures (SSTs) are predicted to rise 1.8–4°C by the year 2100 with increasing atmospheric CO2 emissions (Meehl et al. 2007), there will likely be more frequent bleaching-related mortality of corals, unless corals can undergo rapid thermal acclimation or adaptation (Hoegh-Guldberg 1999; Jones 2008). Rising temperatures are predicted to have detrimental effects on tropical marine larvae by negatively impacting development and survivorship (Munday et al. 2009). Less clear is how an increase in temperature will affect early life history stages of corals that are undergoing the process of symbiosis establishment.
The importance of considering the coral holobiont and the early life history stages of corals when predicting coral adaptation to climate change is now recognized (Coles and Brown 2003; Day et al. 2008). Previous research on coral larvae and stress has examined aposymbiotic larvae (larvae without dinoflagellate symbionts) and thus focused almost exclusively on host responses, such as settlement and survivorship, in isolation. A pattern of increased larval settlement at higher temperatures with short-term (min–h) exposure (Edmondson 1929; Coles 1985; Nozawa and Harrison 2007) and decreased settlement at higher temperatures with longer durations (days–months) of exposure (Jokiel and Guinther 1978; Wilson and Harrison 1997; Bassim and Sammarco 2003; Nozawa and Harrison 2007; Randall and Szmant 2009a) emerged from these studies. Many experiments have found increased larval mortality in response to higher temperature (Bassim and Sammarco 2003; Nozawa and Harrison 2007; Randall and Szmant 2009a, b). Overall, these results indicate that increased temperatures ranging from 30 to 33°C affect larval development, survivorship, and settlement.
A small number of investigations of coral larvae from two genera of coral (Porites and Acropora) have considered the coral holobiont response to elevated temperature. The Porites studies had conflicting results; one experiment found no difference in symbiont density in response to high temperature (Edmunds et al. 2001); another found that larvae had the highest dinoflagellate densities at intermediate temperatures (Edmunds et al. 2005). In contrast, Baird et al. (2006) found no significant difference in survivorship between symbiotic and aposymbiotic larvae exposed to elevated temperatures for 3–7 days in Acropora, a result they attributed to low densities of symbionts in larvae at the start of the experiment. Yakovleva et al. (2009), however, found significantly decreased survivorship and increased SOD and malondialdehyde (MDA) levels (a measure of oxidative damage) in symbiotic larvae compared to aposymbiotic larvae after 3 days of high temperature exposure. In another study of Acropora, Baird et al. (2010) found that initial infection rates of two out of three Symbiodinium strains decreased at the highest temperature (the third strain had low infection at all temperatures). They also found that at high temperature, larvae with high symbiont density had lower survival. The contrasting results of these studies demonstrate our limited understanding of how thermal stress conditions affect the coral–dinoflagellate relationship during the earliest stages of the association and were the inspiration for exploring this topic in a different coral–dinoflagellate model system.
We used larvae of the solitary Hawaiian coral, Fungia scutaria, to carry out laboratory-based experiments of the holobiont response to elevated temperature. This species obtains its symbionts via horizontal transmission and was developed as a model for studies of symbiosis onset over a decade ago (Schwarz et al. 1999). In this investigation, the goals were (1) to quantify the effects of elevated temperatures on the establishment of symbiosis; (2) to quantify the effects of elevated temperatures on larval survivorship; and (3) to compare the survivorship of newly symbiotic and aposymbiotic larvae exposed to thermal stress. Our results demonstrate the following: (1) significantly reduced levels of larval infection due to temperature and duration of thermal exposure; (2) significantly decreased overall larval survival due to elevated temperature; and (3) significantly decreased survivorship of newly symbiotic larvae compared to aposymbiotic larvae exposed to high temperature. We have added to the understanding of elevated temperature impacts on early life history stages of corals by demonstrating that both larval infection and survivorship, as well as survivorship of newly symbiotic larvae compared with aposymbiotic larvae, are reduced when exposed to elevated temperature. Our work also highlights the need to better understand the underlying mechanisms involved in these responses.
Materials and methods
Collection and maintenance of coral larvae
Adult Fungia scutaria were collected from multiple reefs in Kaneohe Bay (Oahu, Hawaii) and maintained year-round in seawater tables at the Hawaii Institute of Marine Biology, University of Hawaii’s marine laboratory. Fungia scutaria broadcast spawn aposymbiotic gametes between 17:00 and 19:00 h, 2–4 days following the full moon in the summer months (Krupp 1983). Preliminary experiments were performed during the July 2007 and July 2008 spawning events; the final experiment was performed during the July 2009 spawning event. Eggs from individual females were kept in separate bowls with 11 of 0.22 μm filter-sterilized seawater (FSW) at densities of about 50 eggs ml−1. Sperm from multiple males (>3) were mixed together prior to fertilizing each bowl of eggs and diluted as described in Krupp et al. (2006). Bowls were placed in outdoor tables with a continuous flow of seawater around the bowls at ambient temperature (average 27°C). Larvae that developed in each bowl were given a 100% water change daily with 0.22 μm FSW and checked for the development of an oral pore using a compound microscope.
Preparation of dinoflagellates
In Hawaii, F. scutaria associate solely with Symbiodinium sp. clade C, subclade 1f (Rodriguez-Lanetty et al. 2004). Since F. scutaria symbionts have not been cultured, dinoflagellates were extracted from adults. Coral tissue containing C1f symbionts was removed from the skeleton of adult F. scutaria using an oral hygiene device (Water Pik). The mixture of host tissue and dinoflagellates was further separated by homogenization with a glass tissue homogenizer and centrifugation at 6,000 × g. Homogenization of the dinoflagellate pellet in FSW was repeated several times to remove most host tissue. The isolated dinoflagellate cells were checked for host tissue debris and quantified using a hemocytometer. Cleaned dinoflagellates were used within 2 h of preparation.
Experimental design
Symbiotic versus aposymbiotic larvae at treatment temperature
Infections in the Fungia system are typically performed at 3–4 days post-fertilization after larvae have developed the ability to acquire symbionts through the oral opening and before settlement begins (Schwarz et al. 1999).
2008 pilot experiment
In July 2008, a preliminary exposure of F. scutaria larvae to elevated temperature was conducted to determine the best design for the study. Larvae and dinoflagellates were exposed to 27 or 32°C for 1 h prior to infection, throughout the 3-h infection period, and up to 48 h post-infection. We used a concentration of 200 larvae ml−1 and infected larvae with 75 dinoflagellates per larva (Table 1). Samples were taken at 5 time points starting immediately post-infection up to 48 h post-infection.
2009 experiment
Results from the 2008 pilot experiment (see ahead) led us to decrease the concentration of larvae and dinoflagellates for the 2009 experiment. Four days post-fertilization, larvae from 7 bowls were combined, concentrated, and counted using a Sedgewick-Rafter Counting Cell. Larvae were distributed among square dishes with dimensions of 4.5 × 4.5 × 1.5 cm (infection success experiment) or 24-well culture dishes (survivorship experiment) at a concentration of 97 larvae ml−1 (Table 1). These dishes were placed into one of three constant temperature waterbaths (PolyScience digital waterbath) used to maintain experimental temperatures of 27, 29, and 31°C.
Larvae and isolated dinoflagellates were incubated separately in their treatment temperature for 1 h prior to infection. In all cases, the temperatures during this pre-incubation period reflected the mean temperatures reported in Table 2. Following the pre-incubation period, isolated dinoflagellates were mixed with homogenized Artemia sp. to stimulate a feeding response in the larvae (Schwarz et al. 1999) and distributed to the “symbiotic treatment” wells of larvae at a concentration of 26 dinoflagellates per larva (Table 1). “Aposymbiotic treatment” wells of larvae received homogenized Artemia sp. only. After the 3-h infection period, all larvae were washed to remove any remaining dinoflagellates and other debris by concentrating larvae onto a 50-μm mesh filter, rinsing them with FSW at treatment temperature, and placing them into clean dishes. Once the washing was complete, this was considered to be 0 h post-infection.
Water was changed once a day in each dish, and dissolved oxygen levels in individual replicates were measured prior to each sampling point over the course of the experiment using a YSI 55 handheld meter. Waterbaths were equipped with HOBO Temperature Pendant data loggers (Onset) to record temperature every 10 min. Waterbaths were located inside a laboratory with standard overhead fluorescent lights with a 12:12-h light/dark photoperiod.
Infection success
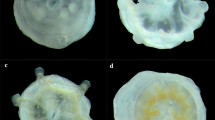
At each time point, larvae were collected from each replicate “symbiotic treatment” dish and fixed in 750 μl Z-fix (Anatech Ltd). To quantify infection success, larvae were rinsed 3 times in 1 × PBS and placed on a slide with a coverslip. Excess liquid was removed from under the coverslip, and larvae were visualized with a compound microscope (Weis et al. 2001) (Fig. 1). For each treatment replicate, infection success was determined as (1) percent of larvae infected in 100 larvae and (2) average density of dinoflagellates in 30 infected larvae.

Micrograph of fixed Fungia scutaria larvae showing how infection levels and symbiont density are visualized under a compound microscope for quantification. Algal symbionts appear as brown circles
Survivorship
To follow survivorship of symbiotic and aposymbiotic larvae at each treatment temperature, the number of larvae remaining in each well was measured at 7 time points post-infection. The entire 1 ml volume of each well was pipetted onto a Sedgewick-Rafter Counting Cell and photographed with a Macrofire digital camera attached to an Olympus light microscope using PictureFrame for Mac 2.0 software. Larvae were then pipetted back into their treatment well, and each culture dish was placed back in its treatment temperature. At the final time point (72 h), larvae were fixed in Z-fix to determine infection success as described above. Survivorship was measured by counting the number of larvae present in each photograph.
Statistical analysis
Statistical comparisons for percent infection success and symbiont density were computed using two-way analysis of variance (ANOVA) with temperature (3 levels) and duration of exposure (5 levels) as fixed factors. An additional one-way ANOVA (temperature, 3 levels, fixed) was performed for the infection success data from the survivorship experiment for the final time point, t = 72 h. All data were evaluated through graphical analyses and had acceptable normality and heteroscedasticity to meet the assumptions of ANOVA. Post hoc analyses using Tukey’s honestly significant difference (HSD) test were performed to determine which treatments differed when the ANOVA was significant at p < 0.05. The statistical program R version 2.10.1 (http://cran.r-project.org) was used for all analyses.
Survivorship data often have a right skewed distribution, violating normality assumptions necessary for traditional parametric statistical methods (Machin et al. 2006). Nonparametric survival analyses, like Kaplan–Meier (or product-limit) analyses, overcome this limitation and allow for “interval-censored” data where the exact time of death of each individual is not known (Machin et al. 2006). In this study, survivorship of larvae was determined at fixed time points, so that the number of larvae that died between each time point is known, but not their exact time of death. Therefore, Kaplan–Meier estimates were computed for all survivorship data, and survivorship curves were plotted. For all estimates, the census time at which the larva was first observed dead (i.e., absent from the photograph) was used as the time of death, so that mortality was not overestimated at earlier time points. Since nonparametric analyses cannot take replicates into account explicitly, data from replicate wells were pooled for the survivorship analyses. To examine the effect of temperature on overall survivorship, data for newly symbiotic and aposymbiotic larvae were combined for each temperature. Two or more survival curves can be compared with hypothesis tests to determine whether they differ statistically (Machin et al. 2006). Log-rank tests (also known as Cox–Mantel tests) were used to compare the Kaplan–Meier survival curves. The statistical program R version 2.10.1 was used for all analyses. Kaplan–Meier estimates were generated using the survfit function, and log-rank tests were conducted with the survdiff function of the survival package in R.
Results
Infection success
2008 pilot experiment
Infection success was dramatically reduced in larvae incubated at 32°C compared to 27°C starting immediately post-infection. Larvae in the control temperature maintained an average infection level of 67%, whereas larvae at the high temperature were never infected more than 7%. Larvae at 27°C had an average density of infection that started at 18 dinoflagellates per larva, then decreased to 14 dinoflagellates per larva, and leveled out at 15 dinoflagellates per larva for the final 3 time points, while larvae at 32°C had very low overall densities throughout the experimental period (average density of 2.8 dinoflagellates per larva immediately post-infection and 1.7 by 48 h post-infection).
2009 experiment
Proportion of larvae infected
Both the length of exposure (F 4,45 = 6.67, p < 0.001) and treatment temperature (F 2,45 = 45.20, p < 0.001) affected the proportion of larvae infected with symbionts (Fig. 2a; Table 3). There was no significant interaction between the effects of time and temperature on the proportion of larvae infected (F 8, 45 = 1.84, p = 0.094, Table 3). The overall effect of treatment temperature was that there were fewer larvae infected at higher temperature compared with larvae at the control temperature (Fig. 2a asterisks). The overall effect of length of exposure was a significant decrease in infection level for larvae at 29°C by 21.5 h post-infection (Fig. 2a letters). A similar effect of length of exposure was seen for larvae at 31°C by 21.5 h post-infection. In contrast, length of exposure did not result in lower infection levels for larvae at 27°C, where even at 36 h post-infection, infection levels remained high.

Infection success of Fungia scutaria larvae incubated at experimental temperatures for up to 40 h (36 h post-infection) in July 2009. a Changes in % infection over time at three temperatures. Values are mean ± SE [n(dishes per temperature) = 4]. The letters a, b and x, y denote a significant difference in percent infection within a temperature over time as determined by Tukey’s HSD post hoc test. Asterisks denote a significant difference in percent infection between that temperature and 27°C at a single time point as determined by Tukey’s HSD post hoc test. b Average density of dinoflagellates in larvae at three temperatures. Values are mean ± SE [n(dishes per temperature) = 4]
Density of symbionts
Neither temperature nor length of exposure significantly affected the density of symbionts (Fig. 2b). An average of 3.8 dinoflagellates per larva was maintained for larvae at 27°C. Larvae at 29°C had a slightly higher density of symbionts (5.3) at the earliest time point before decreasing to a final level of 3.4; the average density of symbionts for larvae at 29°C was 3.8. Larvae at 31°C had a slightly lower average density of 2.6 dinoflagellates per larva.
Survivorship
Exposure to elevated temperature decreased overall larval survival (Fig. 3a). Larvae at 27°C exhibited high levels of survivorship that decreased slightly over time. Survival of larvae at 29°C was significantly lower than larvae at 27°C (log-rank test, p = 0.0394). Larvae exposed to 31°C had the lowest survivorship, with steep declines apparent at the final three time points, and were significantly lower than larvae at 29°C (log-rank test, p < 0.001) and 27°C (log-rank test, p < 0.001).

Kalpan–Meier estimated survival curves for all replicates pooled of Fungia scutaria larvae for up to 76 h (72 h post-infection) in July 2009. a Survival curves for larvae at three temperatures (values for aposymbiotic and symbiotic larvae were combined for each temperature). b Survival curves for aposymbiotic (apo) and symbiotic (sym) larvae at 31°C
The presence of symbionts further reduced survivorship only in larvae in the 31°C treatment (Fig. 3b). Newly symbiotic larvae at 31°C had clearly lower survivorship than aposymbiotic larvae at 31°C at the final four time points, and the difference between the two curves was significant (log-rank test, p = 0.0382). Differences in survivorship due to symbiotic status were not found for larvae at 27°C (log-rank test, p = 0.153) or 29°C (log-rank test, p = 0.456).
Infection success (percent infection) was also measured using larvae from the survivorship experiment (at 72 h), and results indicated a significant difference in infection success among temperature treatments (one-way ANOVA, F 2,9 = 7.23, p = 0.013, Table 3). Larvae at 27°C had a 66% infection level, larvae at 28°C had a 57.5% infection level, and larvae at 31°C had a 42% infection level. The 24% difference in infection success between larvae at 27°C and larvae at 31°C was significant (Tukey’s HSD p = 0.011). These levels mirrored the results of the infection success experiment (Fig. 2a), reinforcing those results and making comparisons between the two experiments possible.
Environmental parameters
Parameters measured for each experiment, including the average temperatures and dissolved oxygen levels maintained, are summarized in Table 2. Dissolved oxygen levels ranged from 5.56 (88%) to 6.08 mg l−1 (92.8%) at 27°C and from 4.98 (81.3%) to 5.58 mg l−1 (86.2%) at 31°C (Table 2), indicating that larvae did not experience hypoxic conditions, which could have confounded the effect of temperature.
Discussion
In this study, we found that thermal stress negatively impacted symbiosis onset and larval survivorship and had a stronger impact on the survivorship of newly symbiotic larvae than aposymbiotic larvae. First, elevated temperature reduced the success of symbiont infection in coral larvae. A modest 2°C increase in temperature for a day and a half significantly impacted symbiosis onset (Fig. 2a). Furthermore, a 4°C increase in temperature above ambient for a period of only a few hours significantly reduced the proportion of larvae infected with symbionts (Fig. 2a). Our survivorship results indicate that infection rates decline at elevated temperature because larvae with symbionts are dying relatively faster than larvae without symbionts (Fig. 3).
The density of symbionts was low in all treatments, including controls, and did not change over time within and between temperature treatments at any time point (Fig. 2b). This result may be due to the amount of dinoflagellates provided for infection (26 dinoflagellates per larva), which was much less than previous studies with the Fungia-Symbiodinium sp. C1f system where infection with “saturating concentrations” of dinoflagellates resulted in higher density levels in controls (Weis et al. 2001; Rodriguez-Lanetty et al. 2006). The lower level of dinoflagellates provided for infection was chosen based on pilot temperature stress studies conducted in July 2008 where infections with higher concentrations of dinoflagellates (75 dinoflagellates per larva) and larvae (200 larvae per ml) led to extremely low infection rates in larvae at high temperature starting immediately post-infection (see “Results”). It is possible that the low infection rates in the pilot experiment could have resulted from low oxygen levels in treatment wells due to increased larval and dinoflagellate respiration at high temperature (Karako-Lampert et al. 2005; Edmunds et al. 2005). Future temperature stress experiments should test infection with higher amounts of dinoflagellates to obtain higher levels of symbiont density in controls.
The onset of symbiosis at ambient temperatures in corals likely involves a complex series of events including pre- and post-phagocytic host–symbiont recognition events (Fitt and Trench 1983; Chen et al. 2004; Rodriguez-Lanetty et al. 2006; Wood-Charlson et al. 2006; Dunn and Weis 2009). It is unclear how these events are affected by elevated temperature. Corals may recognize and reject compromised (e.g., heat stressed) dinoflagellates during these early stages. Recent studies have demonstrated that ROS produced by stressed dinoflagellates may be toxic to coral larvae (Lesser 2006; Yakovleva et al. 2009). Yakovleva et al. (2009) showed that symbiotic larvae exposed to high temperature had substantially higher SOD activity levels and more cellular damage (MDA) than aposymbiotic larvae at the same temperature. Interestingly, larvae at all three temperatures in our experiment had similar initial infection success, but the proportion of larvae infected decreased through time for larvae at elevated temperature, while infection levels for larvae at the control temperature remained stable. Larval survival also decreased through time in symbiotic larvae faster than in aposymbiotic larvae at high temperature. Together, these results suggest that symbionts may pose a liability for coral larvae under elevated temperatures, providing indirect evidence of a larval response to elevated dinoflagellate ROS or to other cellular mechanisms, such as a host innate immune response to a compromised symbiont (Weis 2008).
A proportion of larvae remained infected with the homologous symbiont type C1f at 31°C at 36 h post-infection (43%, Fig. 2a), indicating that although some symbiotic larvae are dying at high temperature, some symbionts are still tolerated at this time point. This result contrasts with previous studies of F. scutaria infected with heterologous types of Symbiodinium spp. at ambient temperature, which are initially taken up but subsequently completely eliminated from larvae by 14 h post-infection (Weis et al. 2001; Rodriguez-Lanetty et al. 2006; Dunn and Weis 2009). The complete elimination of heterologous Symbiodinium types at ambient temperature is due, at least in part, to the activation of post-phagocytic apoptosis (Dunn and Weis 2009). We conclude that there are differences in the timing and nature of cellular events of larvae responding to infection with a heterologous symbiont at ambient temperature (rapid apoptosis resulting in symbiont loss) compared to infection with a homologous symbiont at elevated temperature (slower larval death possibly due to elevated ROS and other mechanisms). Further studies designed to examine how larvae respond to thermal stress during infection with different Symbiodinium types will help tease apart these mechanisms. Rodriguez-Lanetty et al. (2009) explored the transcriptional response of aposymbiotic larvae to elevated temperature. Transcriptional responses to temperature when different types of Symbiodinium are present would be informative, particularly since genes involved in protecting cells from oxidative stress were not upregulated in the first study (Rodriguez-Lanetty et al. 2009).
The degree of larval survivorship was reduced by each temperature increase above the control temperature (Fig. 3a). Kaplan–Meier survival curves demonstrate that the cohort of larvae at 31°C had large losses in survivorship as the duration of exposure increased. Microscopic observations of larvae indicated that exposure to 31°C resulted in numerous sublethal effects that negatively impacted larval behavior, development, and settlement. At the start of the experiment, all larvae were approximately 100 μm long, ciliated, barrel-shaped, and swimming actively. Larvae in the control temperature followed a normal progression of developmental stages through the course of the experiment from swimming actively to creeping along the substrate to settling (Schwarz et al. 1999). After 76 h of exposure to elevated temperature, most of the larvae in the 31°C treatment were small and spherical or irregularly shaped and exhibited clear developmental abnormalities. All larvae exposed to 31°C exhibited increasingly limited movement as the experiment progressed, and by t = 72 h, although they were resting on the bottom of the culture well, most had not settled. These sublethal effects were similar to those found for larvae of other coral species (Randall and Szmant 2009a, b) and F. scutaria embryos (Krupp et al. 2006) and indicate that increased temperature impairs normal physiological processes and larval morphogenesis. Given these observations, it is highly unlikely that heat-stressed F. scutaria larvae would survive to settlement and metamorphosis and grow into healthy adult corals.
Survivorship levels between symbiotic and aposymbiotic larvae differed when exposed to high temperature, where the presence of symbionts in larvae at 31°C posed an increased risk of mortality (Fig 3b). This result implies that where onset of symbiosis coincides with an increase in seawater temperature, larvae may have to contend with costs associated with increased ROS production and oxidative cellular damage and decreased benefits such as less carbon being translocated from the symbiont to the host due to changes in energy production and demand (Iglesias-Prieto et al. 1992; Edmunds et al. 2001; Ferrier-Pages et al. 2007; Yakovleva et al. 2009).
The temperatures used in this investigation were chosen to reflect the local environment, since variation in normal summer temperature maxima that exists across regions leads to geographic disparity in coral responses to temperatures (Coles et al. 1976; Jokiel and Coles 1990). In Hawaii, there has been a warming trend in SSTs over the past several decades (Jokiel and Coles 1990; Jokiel and Brown 2004), and bleaching events in 1996 and 2002 have been attributed to temperature anomalies exceeding +1°C (Jokiel and Brown 2004). In Kaneohe Bay, the average summer monthly temperature is 27 ± 1°C (Jokiel and Coles 1977), and it regularly reaches temperatures of 29°C during midday on reef flats (Krupp et al. 2006). As the temperature of Hawaiian waters increases, larvae of corals that spawn during summer months in this region, including F. scutaria, will be subject to temperatures similar to those examined here.
Results from this study indicate that exposure to elevated temperatures during the initiation of symbiosis impairs the ability of coral larvae to maintain an endosymbiotic relationship with newly acquired dinoflagellate symbionts. The cost of establishing symbiosis may prove too great if temperatures are elevated beyond the ability of larvae to cope with increased levels of oxidative stress or to proceed through normal cellular development. If SSTs continue to increase over the next several decades as predicted, the symbioses that coral larvae form may change in response to changing environments (Baird et al. 2007), although most corals may not exchange Symbiodinium types (Goulet 2006) and the long-term stability of new relationships that do arise remains an open question (Coffroth et al. 2010, LaJeunesse et al. 2010). Thus, the potential for coral larvae to continue to form stable symbiotic relationships with homologous symbiont types and survive to establish the next generation of corals may be greatly diminished.
References
Baird AH, Gilmour JP, Kamiki T, Nonaka M, Pratchett MS, Yamamoto HH, Yamasaki H (2006) Temperature tolerance of symbiotic and non-symbiotic coral larvae. Proc 10th Int Coral Reef Symp 1:38–42
Baird AH, Cumbo VR, Leggat B, Rodriguez-Lanetty M (2007) Fidelity and flexibility in coral symbioses. Mar Ecol Prog Ser 347:307–309
Baird AH, Guest JR, Willis BL (2009) Systematic and biogeographical patterns in the reproductive biology of scleractinian corals. Annu Rev Ecol Evol Syst 40:551–571
Baird AH, Bhagooli R, Nonaka M, Yakovleva I, Yamamoto HH, Hidaka M, Yamasaki H (2010) Environmental controls on the establishment and development of algal symbiosis in corals. Proc 11th Int Coral Reef Symp 1:108–112 Session number 5
Bassim K, Sammarco P (2003) Effects of temperature and ammonium on larval development and survivorship in a scleractinian coral (Diploria strigosa). Mar Biol 142:241–252
Carpenter KE, Abrar M, Aeby G, Aronson RB, Banks S, Bruckner A, Chiriboga A, Cortes J, Delbeek JC, DeVantier L, Edgar GJ, Edwards AJ, Fenner D, Guzman HM, Hoeksema BW, Hodgson G, Johan O, Licuanan WY, Livingstone SR, Lovell ER, Moore JA, Obura DO, Ochavillo D, Polidoro BA, Precht WF, Quibilan MC, Reboton C, Richards ZT, Rogers AD, Sanciangco J, Sheppard A, Sheppard C, Smith J, Stuart S, Turak E, Veron JEN, Wallace C, Weil E, Wood E (2008) One-third of reef-building corals face elevated extinction risk from climate change and local impacts. Science 321:560–563
Chen M, Cheng Y, Hong M, Fang L (2004) Molecular cloning of Rab5 (ApRab5) in Aiptasia pulchella and its retention in phagosomes harboring live zooxanthellae. Biochem Biophys Res Commun 324:1024–1033
Coffroth MA, Poland DM, Petrou EL, Brazeau DA, Holmberg JC (2010) Environmental symbiont acquisition may not be the solution to warming seas for reef-building corals. PLoS ONE 5:e13258
Coles S (1985) The effects of elevated temperature on reef coral planulae settlement as related to power plant entrainment. Proc 5th Int Coral Reef Symp 4:171–176
Coles S, Brown B (2003) Coral bleaching—capacity for acclimatization and adaptation. Adv Mar Biol 46:183–223
Coles SL, Jokiel PL, Lewis CL (1976) Thermal tolerance in tropical versus subtropical Pacific reef corals. Pac Sci 30:159–166
Colley NJ, Trench RK (1983) Selectivity in phagocytosis and persistence of symbiotic algae by the scyphistoma stage of the jellyfish Cassiopeia xamachana. Proc R Soc Lond B Biol Sci 219:61–82
Day T, Nagel L, van Oppen MH, Caley MJ (2008) Factors affecting the evolution of bleaching resistance in corals. Am Nat 171:E72–E88
Douglas AE (2003) Coral bleaching–how and why? Mar Pollut Bull 46:385–392
Douglas AE (2008) Conflict, cheats and the persistence of symbioses. New Phytol 177:849–858
Dubinsky Z (1990) Coral reefs. Elsevier, New York
Dunn SR, Weis VM (2009) Apoptosis following post-phagocytic symbiont recognition in the coral, Fungia scutaria. Environ Microbiol 11:268–276
Edmondson CH (1929) Growth of Hawaiian corals. Bernice Bishop Mus Bull 58:1–38
Edmunds PJ, Gates RD, Gleason DF (2001) The biology of larvae from the reef coral Porites astreoides, and their response to temperature disturbances. Mar Biol 139:981–989
Edmunds PJ, Gates RD, Leggat W, Hoegh-Guldberg O, Allen-Requa L (2005) The effect of temperature on the size and population density of dinoflagellates in larvae of the reef coral Porites astreoides. Invertebr Biol 124:185–193
Fadlallah YH (1983) Sexual reproduction, development and larval biology in scleractinian corals. Coral Reefs 2:129–150
Ferrier-Pages C, Richard C, Forcioli D, Allemand D, Pichon M, Shick J (2007) Effects of temperature and UV radiation increases on the photosynthetic efficiency in four scleractinian coral species. Biol Bull 213:76–87
Fitt WK, Trench RK (1983) Endocytosis of the symbiotic dinoflagellate Symbiodinium microadriaticum Freudenthal by endodermal cells of the scyphistomae of Cassiopeia xamachana and resistance of the algae to host digestion. J Cell Sci 64:195–212
Furla P, Allemand D, Shick JM, Ferrier-Pages C, Richier S, Plantivaux A, Merle P-L, Tambutte S (2005) The symbiotic anthozoan: a physiological chimera between alga and animal. Integr Comp Biol 45:595–604
Glynn PW (1993) Coral reef bleaching: ecological perspectives. Coral Reefs 12:1–17
Goulet T (2006) Most corals may not change their symbionts. Mar Ecol Prog Ser 321:1–7
Harrison P, Wallace C (1990) Reproduction, dispersal and recruitment of scleractinian corals. In: Dubinsky Z (ed) Ecosystems of the World: coral reefs. Elsevier, New York, pp 133–207
Hoegh-Guldberg O (1999) Climate change, coral bleaching, and the future of the world’s coral reefs. Mar Freshw Res 50:839–866
Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS, Greenfield P, Gomez E, Harvell CD, Sale PF, Edwards AJ, Caldeira K, Knowlton N, Eakin CM, Iglesias-Prieto R, Muthiga N, Bradbury RH, Dubi A, Hatziolos ME (2007) Coral reefs under rapid climate change and ocean acidification. Science 318:1737–1742
Iglesias-Prieto R, Matta JL, Robins WA, Trench RK (1992) Photosynthetic response to elevated temperature in the symbiotic dinoflagellate Symbiodinium microadriaticum in culture. Proc Natl Acad Sci USA 89:10302–10305
Jokiel PL, Brown EK (2004) Global warming, regional trends and inshore environmental conditions influence coral bleaching in Hawaii. Global Change Biol 10:1627–1641
Jokiel PL, Coles SL (1977) Effects of temperature on the mortality and growth of Hawaiian reef corals. Mar Biol 43:201–208
Jokiel PL, Coles SL (1990) Response of Hawaiian and other Indo-Pacific reef corals to elevated temperature. Coral Reefs 8:155–162
Jokiel PL, Guinther EB (1978) Effects of temperature on reproduction in the hermatypic coral Pocillopora damicornis. Bull Mar Sci 28:786–789
Jones R (2008) Coral bleaching, bleaching-induced mortality, and the adaptive significance of the bleaching response. Mar Biol 154:65–80
Karako-Lampert S, Katcoff DJ, Achituv Y, Dubinsky Z, Stambler N (2005) Responses of Symbiodinium microadriaticum clade B to different environmental conditions. J Exp Mar Biol Ecol 318:11–20
Krupp DA (1983) Sexual reproduction and early development of the solitary coral Fungia scutaria (Anthozoa: Scleractinia). Coral Reefs 2:159–164
Krupp D, Hollingsworth L, Peterka J (2006) Elevated temperature sensitivity of fertilization and early development in the mushroom coral Fungia scutaria Lamarck 1801. Proc 10th Int Coral Reef Symp 1:71–77
LaJeunesse TC, Smith R, Walther M, Pinzón J, Pettay DT, McGinley M, Aschaffenburg M, Medina-Rosas P, Cupul-Magaña AL, López Pérez A, Reyes-Bonilla H, Warner ME (2010) Host-symbiont recombination versus natural selection in the response of coral-dinoflagellate symbioses to environmental disturbance. Proc R Soc B 277:2925–2934
Lesser MP (2006) Oxidative stress in marine environments: biochemistry and physiological ecology. Annu Rev Physiol 68:253–278
Machin D, Cheung YB, Parmar MKB (2006) Survival analysis: a practical approach, 2nd edn. Wiley, Chichester
Marlow HQ, Martindale MQ (2007) Embryonic development in two species of scleractinian coral embryos: Symbiodinium localization and mode of gastrulation. Evol Dev 9:355–367
Meehl G, Stocker T, Collins W, Friedlingstein P, Gaye A, Gregory J, Kitoh A, Knutti R, Murphy J, Noda A, Raper S, Watterson I, Weaver A, Zhao Z (2007) Global climate projections. Climate change 2007: the physical science basis. Cambridge University Press, Cambridge
Munday P, Leis J, Lough J, Paris C, Kingsford M, Berumen M, Lambrechts J (2009) Climate change and coral reef connectivity. Coral Reefs 28:379–395
Muscatine L, Hand C (1958) Direct evidence for the transfer of materials from symbiotic algae to the tissues of a coelenterate. Proc Natl Acad Sci USA 44:1259–1263
Nozawa Y, Harrison P (2007) Effects of elevated temperature on larval settlement and post-settlement survival in scleractinian corals, Acropora solitaryensis and Favites chinensis. Mar Biol 152:1181–1185
Oliver TH, Leather SR, Cook JM (2009) Tolerance traits and the stability of mutualism. Oikos 118:346–352
Randall CJ, Szmant AM (2009a) Elevated temperature reduces survivorship and settlement of the larvae of the Caribbean scleractinian coral, Favia fragum (Esper). Coral Reefs 28:537–545
Randall CJ, Szmant AM (2009b) Elevated temperature affects development, survivorship, and settlement of the Elkhorn coral, Acropora palmata (Lamarck 1816). Biol Bull 217:269–282
Richier S, Furla P, Plantivaux A, Merle P-L, Allemand D (2005) Symbiosis-induced adaptation to oxidative stress. J Exp Biol 208:277–285
Rodriguez-Lanetty M, Krupp DA, Weis VM (2004) Distinct ITS types of Symbiodinium in Clade C correlate with cnidarian/dinoflagellate specificity during onset of symbiosis. Mar Ecol Prog Ser 275:97–102
Rodriguez-Lanetty M, Wood-Charlson EM, Hollingsworth L, Krupp D, Weis V (2006) Temporal and spatial infection dynamics indicate recognition events in the early hours of a dinoflagellate/coral symbiosis. Mar Biol 149:713–719
Rodriguez-Lanetty M, Harii S, Hoegh-Guldberg O (2009) Early molecular responses of coral larvae to hyperthermal stress. Mol Ecol 18:5101–5114
Schwarz JA, Krupp DA, Weis VM (1999) Late larval development and onset of symbiosis in the scleractinian coral Fungia scutaria. Biol Bull 196:70–79
Stachowicz JJ (2001) Mutualism, facilitation, and the structure of ecological communities. Bioscience 51:235–246
Wakefield TS, Farmer MA, Kempf SC (2000) Revised description of the fine structure of in situ “zooxanthellae” genus Symbiodinium. Biol Bull 199:76–84
Weis VM (2008) Cellular mechanisms of cnidarian bleaching: stress causes the collapse of a symbiosis. J Exp Biol 211:3059–3066
Weis VM (2010) The susceptibility and resilience of corals to thermal stress: adaptation, acclimatization or both? Mol Ecol 19:1515–1517
Weis VM, Reynolds WS, deBoer MD, Krupp DA (2001) Host-symbiont specificity during onset of symbiosis between the dinoflagellates Symbiodinium spp. and planula larvae of the scleractinian coral Fungia scutaria. Coral Reefs 20:301–308
Wilson JR, Harrison PL (1997) Sexual reproduction in high latitude coral communities at the Solitary Islands, Eastern Australia. Proc 8th Int Coral Reef Symp 1:533–538
Wood-Charlson EM, Hollingsworth LL, Krupp DA, Weis VM (2006) Lectin/glycan interactions play a role in recognition in a coral/dinoflagellate symbiosis. Cell Microbiol 8:1985–1993
Yakovleva IM, Baird AH, Yamamoto HH, Bhagooli R, Nonaka M, Hidaka M (2009) Algal symbionts increase oxidative damage and death in coral larvae at high temperatures. Mar Ecol Prog Ser 378:105–112
Yellowlees D, Rees TAV, Leggat W (2008) Metabolic interactions between algal symbionts and invertebrate hosts. Plant Cell Environ 31:679–694
Acknowledgments
Funding was provided by a PADI Foundation grant; a Dr. Earl and Ethel Myers Oceanographic and Marine Biology Trust grant; and the Lerner-Gray Fund for Marine Research (American Museum of Natural History) to C.E. Schnitzler; and by National Science Foundation #IOS-0542452 and #IOS-0919073 to V.M. Weis. We thank Andrew Baird, Santiago Perez, Mauricio Rodriguez-Lanetty, and Cliff Ross and for helpful advice; Elisha Wood-Charlson, Jessi Kershner, and Olivier Detournay for assistance in the field; Jeremy Polk for help counting larvae; and Ruth Gates for generously providing equipment and space. This is Hawaii Institute of Marine Biology contribution # 1476.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M. Kühl.
Rights and permissions
About this article
Cite this article
Schnitzler, C.E., Hollingsworth, L.L., Krupp, D.A. et al. Elevated temperature impairs onset of symbiosis and reduces survivorship in larvae of the Hawaiian coral, Fungia scutaria . Mar Biol 159, 633–642 (2012). https://doi.org/10.1007/s00227-011-1842-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-011-1842-0