
Abstract
The results presented in this report are based on analyses of 16,721 days of data downloaded from 96 archival tags recovered from bigeye tuna (Thunnus obesus; 54–159 cm in length, 0.97–5.44 years of age) at liberty from 31 to 1,508 days in the equatorial eastern Pacific Ocean. Analyses of daily timed depth and temperature records resulted in the classification of the data into three daily behavior types: characteristic, associative (associated with floating objects), and other. There is a significant positive correlation between the proportion of time fish exhibit characteristic behavior and increasing length, and significant negative correlations between the proportion of time bigeye exhibit associative and other behavior with increasing length. For the smallest (54–80 cm) to largest (100–159 cm) length classes, the vertical habitats utilized when exhibiting non-associative behaviors were 99 and 98% of the time above the thermocline depth (60 m) during the night, at the same average depth of 34 m, and 60 and 72% of the time below the thermocline during the day at average depths of 163 and 183 m, respectively. For the same smallest to largest length classes, when exhibiting associative behavior, the average nighttime and daytime depths were 25 and 21, and 33 and 37 m, respectively. The apparent effects of the environment on the behavior of the fish are discussed.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
It has long been known that tunas and other large pelagics are attracted to flotsam and jetsam, and fisherman has taken advantage of this associative behavior by constructing and deploying both moored and drifting fish-aggregating devices (FADs; Scott et al. 1999; Le Gall et al. 2000).
Bigeye tuna (Thunnus obesus) are caught in the eastern Pacific Ocean (EPO) primarily by Asian distant water longline vessels targeting them from about 30°N to 30°S, and by industrial purse-seine vessels, of many nations, targeting tuna aggregations associated with drifting floating objects, mostly FADs, from about 5°N to 15°S (Anonymous 2008). Since 1994 catches of bigeye from longline fisheries have significantly decreased and those of the purse-seine fisheries have significantly increased. The purse-seine fishery captures smaller bigeye, and the consequences of that fishery over the past 16 years include reductions in the yield per recruit and maximum sustainable yield. Because the stock of bigeye in the EPO is overfished, there is uncertainty about the sustainability of recent levels of exploitation (Anonymous 2008; Aires-da-Siva and Maunder 2009).
Since changes in behavior and habitat of bigeye tuna affect its vulnerability to capture, understanding the behavior and habitat utilization of bigeye is crucial for improving stock assessments, by including quantitative information for standardization of purse-seine and longline catch per unit of effort (Brill et al. 2005; Maunder 2005; Maunder et al. 2006). Estimation of the occurrence and duration of associative and non-associative events of bigeye with floating objects, where large-scale industrial purse-seine and longline fisheries operate in the EPO, is important for estimation of their relative vulnerability. In addition, understanding the fine-scale behavior of bigeye in association with floating objects can be useful for evaluating potential modifications to purse-seine fishing techniques for reduction of the catch of undesirable sizes of tunas including bigeye, and also other bycatch species.
Historical studies using ultrasonic telemetry of fine-scale movements and short-term behavior of bigeye equipped with acoustic tags, while associated with anchored FADs off Hawaii (Holland et al. 1990) and while non-associated in pelagic waters off French Polynesia (Dagorn et al. 2000), described some associative and non-associative behavior patterns and provided some preliminary information on habitat utilization. Schaefer and Fuller (2005) investigated the fine-scale and short-term horizontal and vertical movements of bigeye and skipjack (Katsuwonus pelamis) tunas concurrently, within large multi-species aggregations, associated with moored buoys and a drifting vessel in the EPO, using ultrasonic telemetry and archival tags (ATs), coupled with acoustic imaging. The simultaneous behavior of bigeye, skipjack, and yellowfin (Thunnus albacares) tunas, while associated with drifting FADs, was also investigated in the western Pacific Ocean (WPO), using ultrasonic telemetry, by Matsumoto et al. (2006).
In recent years, tagging experiments utilizing ATs with free-ranging tunas, including bigeye, have provided extensive long-term data sets, and vital information toward further understanding of the movements, behavior, habitat, and physiology for the species investigated (Block 2005; Schaefer et al. 2009). A previous investigation into bigeye movements, behavior, and habitat in the EPO (Schaefer and Fuller 2002), based on a limited amount of AT data, included discrimination and classification of daily behavior types. Estimates in that study included the frequency and duration of events in which bigeye exhibited characteristic behavior, consisting of diel vertical migrations to daytime depths of about 350 m, and associative behavior with floating objects. The diurnal and seasonal variability in vertical habitat utilization distributions relative to environmental variables were also evaluated in that study. Within the Pacific Ocean, useful information has also been derived from AT data on the vertical movements, behavior, and habitat of bigeye in Hawaiian waters (Musyl et al. 2003) and the Coral Sea (Evans et al. 2008).
The objectives of this investigation are to quantify and elucidate the vertical movements, behavior, and habitat utilization of bigeye in the equatorial EPO, based on analyses of 16,721 days of data downloaded from 96 archival tags recovered from bigeye (54–159 cm in length; 0.97–5.44 years of age) at liberty from 31–1,508 days, collected during 2000–2009. The time-at-depth and temperature data are utilized for the daily classification of distinct behavior types and habitat utilization distributions. The behavioral data are evaluated with respect to length, age, and spatial and temporal characteristics. The results obtained have direct implications for consideration of their inclusion within stock assessments and resource management applications for bigeye in the EPO.
Materials and methods
Tag releases
Tagging was conducted on the chartered FV Her Grace, a 17.7-m live-bait pole-and-line fishing vessel. Bigeye associated with Tropical Atmosphere–Ocean (TAO) moorings, the tagging vessel, and drifting fish-aggregating devices (FADs), were captured, tagged, and released between 2°N and 2°S, and 95° and 97°W, during March to May of 2000 and 2002 through 2005. The details regarding the materials and methods utilized in the capture, tagging, and release of the fish are given by Schaefer and Fuller (2009).
The ATs deployed in 2000 were model Mk7 and in 2002 model Mk9, both manufactured by Wildlife Computers, Redmond, Washington, USA. The ATs deployed in 2003, 2004, and 2005 were model Mk9, and LTD_2310 manufactured by Lotek Wireless, Inc., St. John’s, Newfoundland, Canada. The configuration for each of the ATs deployed was designed for implantation into the peritoneal cavity of the fish, with the light and temperature sensor stalk protruding outside the fish through an incision in the abdominal wall. A label, printed in Spanish, with information about reporting the recovery of the tag and the associated reward (US$500) was encased in the main body of the instruments. The depth (pressure), ambient and internal temperatures, and light-level data were programmed to be stored in the memory of the tags every 4 min for the Mk7 tags deployed in 2000, and every 1 min for the Mk9 and LTD_2310 tags deployed in 2002–2005.
Tag recoveries
Data on the release and recapture dates, locations, and lengths for 96 bigeye for which there are complete data sets of greater than 30 days, along with the corresponding days of archival tag data utilized for each in this study are given in Table 1. Except for 9 of the fish that were recaptured by longline vessels (Table 1), all were recaptured by purse-seine vessels fishing in the vicinity of FADs. The 96 fish ranged from 54.1 to 159.2 cm in length and 0.97–5.44 years of age, and AT data were recovered from those fish while at liberty from 31.2 to 1,508.2 days (mean = 171.4 days), for a total of 16,721 days of AT data (Table 1).
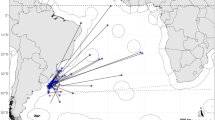
The most probable movement paths for each of those 96 bigeye are shown in Fig. 1. The unscented Kalman filter model (Lam et al. 2008), with sea surface temperature measurements integrated (UKFsst), was used to obtain the most probable tracks and improved estimates of geographic positions, as described by Schaefer and Fuller (2009), for 95 of the fish. The most probable horizontal movement paths for those 95 fish indicate that they spent about 95% of their time in a relatively restricted area between 5°N and 5°S and 85°W and 110°W (Fig. 1). The exception was a bigeye (Tag no. 590051; Table 1) at liberty for 4.1 years. Because of various computational problems encountered for processing of this very large archival tag data set with UKFsst, the most probable movement path was derived using the “trackit” model with SST incorporated (Lam et al. unpublished manuscript). During the first 2 years at liberty, the fish remained within 1,839 km of its release location, a restricted area similar to that occupied by the other 95 fish, but centered slightly to the east. However, during the third and fourth years at liberty the fish undertook two very similar cyclical movements to the central Pacific. The first movement began with a departure near early July in 2007, arriving at about 151°W near early November 2007, and returning to the east, at about 84°W, in early May 2008. The second movement began with a departure near early July 2008, arriving at about 162°W near early December 2008, and returning to the east, at about 84°W, in early May 2009. The fish was recaptured just 1,245 km from where it was tagged and released.

Most probable movement paths for the 96 bigeye tuna archival tag data sets used in this study (see Schaefer and Fuller 2009). Dashed lines represent the position from which a tag failed to the recapture positions
Data processing
Data were downloaded from the recovered tags, using software provided by the tag manufacturers. A Microsoft Access database is used for storage, queries, and some processing of the data. Queries are written to extract information from the database in required formats for use in other software packages. Two of the primary packages are Microsoft Excel for statistical analyses and the Environmental Systems Research Institute (ESRI) ArcView for mapping and spatial analyses. Computer programs were written for the processing of the time-series depth and temperature data, specifically for classification of daily behavior types, using the behavior criteria described elsewhere.
The data sets were separated into periods of nighttime and daytime by using the times of nautical twilight. Nighttime was classified as the period between the time of the first evening record after nautical twilight until the time of the last morning record before nautical twilight. The individual data sets for night and day were used in evaluations of diurnal differences in behavior and habitat utilization.
For each of the 96 fish, the daily behavior for each day at liberty was classified as characteristic, associative (with a floating object), or other, based on the time-series depth and ambient temperature data. A daily position was estimated along the movement paths for each of the 96 fish, based on the methodology described by Schaefer and Fuller (2009), except when no position was available due to the lack of a light-based position estimate, a position was interpolated for that day. For analyses of behavior types, the time-series data were truncated, and only those data recorded after fish had been separated from initial association with floating objects, including the tagging vessel, were utilized.
Characteristic behavior is herein defined as that in which the fish occupies primarily the mixed layer during the night, descends to a depth greater than 150 meters within 1 h of sunrise, remaining for the duration of the day and returning only to depths where temperatures are above 20°C, except if temperatures are below 20°C at depths at which temperatures are within 0.5 degrees of estimated SST, for brief periods not to exceed 30 min until within 1 h of sunset. The fish may not make more that 24 ascents during the day.
The classification of associative behavior, used herein, is based partially on the reported behavioral characteristics from bigeye AT data (Schaefer and Fuller 2002) and ultrasonic telemetry observations of bigeye associated with floating objects in this area (Schaefer and Fuller 2005). Associative behavior with floating objects was defined as that during which bigeye remained 84% or more of the time above the 20°C isotherm during daylight hours for the first day of an associative event, and with subsequent consecutive days classified as associative if 74% or more time was spent above 20°C. The threshold value for subsequent days following the start of an associative event was relaxed from 84 to 74% because there were known associative events for bigeye released and recaptured at the same floating objects in which 74% of their time was spent above 20°C for some subsequent days during that event. Those days in which the time-series data did not satisfy the classification criteria for characteristic or associative behavior were classified as “other”. For each fish, the numbers and durations of each behavior event were determined throughout the period at liberty.
Dives in excess of 500 m were classified, as deep dives, and the time, duration, and maximum depth of each event was determined, for each fish, throughout periods at liberty.
Estimation of the length and/or age of fish, for each day during their respective times at liberty, was done using the equations from Schaefer and Fuller (2006) for fish up to 149 cm, and the equations from Aires-da-Siva and Maunder (2009) for fish of greater length. The estimated lengths and ages along with their respective behavioral data for that day at liberty were utilized in the length and age-specific analyses in this report.
Chi-square goodness of fit analyses was used to compare the frequencies of behavior types, and analyses of variance were performed to compare the mean durations of behavioral events, by length classes, within quarters and years, between years, and for years combined. Pearson correlation analyses was used to test for relationships between behavior patterns and length, age, and event durations (Zar 1974). Hypotheses were rejected at a confidence level of 95% (5% significance level).
Results
Vertical movements and behavior
An example of characteristic behavior exhibited by a 126-cm fish for 1 week is shown in Fig. 2a, and for one day in Fig. 2b. There is a distinct diurnal vertical movement pattern observed, and previously described, from shallow at night to much greater depths, well below the thermocline, during the day (Holland et al. 1990; Schaefer and Fuller 2002). As seen in Fig. 2b, the fish descended before dawn and ascended before dusk, tracking and foraging on the vertically migrating deep-scattering layer (DSL) prey organisms (Josse et al. 1998). During this 24-h period (Fig. 2b), the fish remained for periods of up to 2 h 24 min at depths between 200 and 325 m, and undertook just 5 upward excursions to depths within the mixed layer, less than that of 20°C isotherm.

Depth and temperature records for an estimated 126-cm bigeye tuna (Tag. no. 1113, Table 1), exhibiting characteristic behavior for 1 week (8/29 to 9/4/2003) (a), and one day (9/3/2003) (b)
An example of associative behavior exhibited by a 96-cm fish for 1 week is shown in Fig. 3a, and for one day in Fig. 3b. The fish remained relatively shallow, primarily within the mixed layer depths, and associated with a floating object during this period as previously described (Holland et al. 1990; Schaefer and Fuller 2002). There is a distinct diurnal diving behavior observed for each day in Fig. 3a, shortly before dusk. This diving behavior is apparently timed to coincide with the ascending DSL, which, when present, rises to within mixed layer depths at night.

Depth and temperature records for an estimated 96-cm bigeye tuna (Tag. no. 2332, Table 1), exhibiting associative behavior for 1 week (8/19 to 8/25/2004) (a), and one day (8/22/2004) (b)
An example of other behavior type exhibited by a 62-cm fish for 1 week is shown in Fig. 4a, and for one day in Fig. 4b. There is a distinct diurnal vertical movement pattern observed, from shallow at night and in the morning, to repetitive diving and searching behavior between depths of the mixed layer and that of the DSL during the afternoon, as previously described (Schaefer and Fuller 2002). This behavior pattern apparently results from a different vertical distribution of prey organisms in locations where observed and is most likely an alternative type of foraging strategy.

Depth and temperature records for an estimated 62-cm bigeye tuna (Tag. no. 1082, Table 1), exhibiting other type behavior for 1 week (5/4 to 5/10/2005) (a), and one day (5/9/2005) (b)
The classifications of daily behavior for the 96 bigeye throughout their times at liberty as characteristic, associative, or other, by year and length class, are given in Table 2. Analyses of the percentages of days classified by daily behavior type indicated significant differences between length classes 80.0–99.9 and 100.0–137.0 cm within 2003 (Chi-square test, χ 22 = 25.6, P < 0.001), between length classes 80.0–99.9 and 100.0–135.6 cm within 2004 (Chi-square test, χ 22 = 14.7, P < 0.001), and between length classes 60.0–79.9 and 80.0–99.9 cm within 2005 (Chi-square test, χ 22 = 19.5, P < 0.001). The differences are attributed to the greater percentages of characteristic behavior exhibited in the larger length classes and associative and other behavior in the smaller length classes.
Analyses of the mean durations of events classified by behavior type indicated the only significant differences were for characteristic events between length classes 80.0–99.9 and 100.0–137.0 cm within 2003 (ANOVA, F (1,291) = 4.7, P = 0.03), and between length classes 60.0–79.9 and 80.0–99.9 cm within 2005 (ANOVA, F (1,356) = 6.4, P = 0.01). Analyses of the percentage of days classified by daily behavior type indicated significant differences within length class 80.0–99.9 cm between 2003, 2004, and 2005 (Chi-square test, χ 24 = 11.0, P < 0.05), and within length class 100.0–146.0 cm between 2000, 2003, and 2004 (Chi-square test, χ 24 = 148.5, P < 0.001).
Analyses of the mean durations of events classified by behavior type indicated the only significant difference was for associative events within length class 100.0–146.0 cm between 2000, 2003, and 2004 (ANOVA, F (2,297) = 5.2, P = 0.006).
The percentage of days and mean duration of events classified by behavior type within length class, for years combined, are given in Table 2. Analyses of the percentages of days classified by daily behavior type for years combined indicated significant differences (Chi-square test, χ 24 = 388.5, P < 0.001), among the 54–79.9, 80–99.9, and 100–159.2 cm length classes. The differences are attributed to opposing trends in the higher percentages of characteristic behavior and lower percentages of associative and other behavior with increasing length.
Analyses of the mean durations of events between length classes indicated significant differences for characteristic (ANOVA, F (2,1948) = 8.1, P = 0.0002), and other behavior (ANOVA, F (2,2215) = 4.3, P = 0.01), but not associative behavior (ANOVA, F (2,797) = 1.6, P = 0.20).
The summary statistics for the classification of daily behavior for the 96 bigeye throughout their times at liberty as characteristic, associative, or other, by quarter of the year and length class, are given in Table 3. Analyses of the percentages of days classified by daily behavior type indicated significant differences between length classes 54.0–79.9, 80.0–99.9, and 100.0–159.2 cm within quarter 2 (Chi-square test, χ 24 = 356.9, P < 0.001), and between length classes 80.0–99.9 and 100.0–150.0 cm within quarter 3 (Chi-square test, χ 22 = 17.4, P < 0.001). The differences being attributed to the greater percentages of characteristic behavior exhibited in the larger length classes, and associative and other behavior in the smaller length classes.
Analyses of the mean durations of events classified by behavior type indicated significant differences for characteristic (ANOVA, F (2,797) = 17.5, P < 0.0001), and other (ANOVA, F (2,924) = 6.7, P = 0.001) events between the three length classes within quarter 2. Analyses of the percentage of days classified by daily behavior type indicated significant differences within length class 80.0–99.9 cm between quarters 2 and 3 (Chi-square test, χ 22 = 51.8, P < 0.001), and within length classes greater than 100 cm between quarters 1, 2, 3, and 4 (Chi-square test, χ 26 = 120.7, P < 0.001). Analyses of the mean durations of events classified by behavior type indicated significant differences for characteristic events within length class 80.0–99.9 cm between quarters 2 and 3 (ANOVA, F (1,539) = 22.0, P < 0.0001) and for other events within length classes greater than 100 cm between quarters 1, 2, 3, and 4 (ANOVA, F (3,1228) = 3.9, P = 0.008).
There is a significant positive correlation (Pearson, r 2 = 0.82, n = 21, P < 0.0001) between proportion of time that bigeye exhibit characteristic behavior and increasing length (Fig. 5a). The distribution of the durations of events classified as characteristic (Fig. 6a) shows a peak at about 2 days, with a significant number of observations out to 10 days, and then only sporadic observations to a maximum of 45 days. There is a significant negative correlation (Pearson, r 2 = 0.55, n = 21, P < 0.0001) between the proportion of time that bigeye exhibit associative behavior and increasing length (Fig. 5b). The distribution of the durations of events classified as associative (Fig. 6b) shows a peak at about 2 days, with a significant number of observations out to 4 days, and then only sporadic observations to a maximum of 24 days. There is a significant negative correlation (Pearson, r 2 = 0.36, n = 73, P < 0.0001) between the number of days in which bigeye exhibited associative behavior and the number of days at liberty, utilizing the 73 data sets from Table 1 in which the numbers of days of archival data collected and days at liberty were the same. There is a significant negative correlation (Pearson, r 2 = 0.32, n = 21, P = 0.008) between the proportion of time that bigeye exhibit other behavior with increasing length (Fig. 5c). The distribution of the durations of events classified as other (Fig. 6c) shows a peak at about 2 days, with a significant number of observations out to about 6 days, and then only sporadic observations to a maximum of 33 days.

Proportion of time bigeye spent exhibiting characteristic (a), associative (b), and other (c) type behaviors, by length class, for 16,278 days of data

Distributions of the durations of all characteristic (a), associative (b) and other (c) events
A comparison was conducted of the daily behavior types for 4 bigeye recaptured by purse-seine vessels (Table 1, tag nos.: 787, 814, 821, 826), comprising 449 days of data, and 4 bigeye recaptured by longline vessels (Table 1, tag nos.: 793, 798, 812, 887), comprising 438 days of data. The data sets were truncated for the analyses, so the fish from both groups were of about the same lengths (120–130 cm), during the same period of time, in the same area. The percentages of time, and mean durations of events, in which the purse-seine and longline-caught fish exhibited characteristic behavior was 55.7 (4.2 days) and 61.0% (5.0 days), associative behavior was 4.2 (1.5 days) and 10.3% (2.1 days), and the other behavior type was 40.1 (3.2 days) and 28.7% (2.0 days), respectively. Analyses of the percentages of days classified by daily behavior type indicated significant differences between the two groups (Chi-square test, χ 22 = 20.8, P < 0.001). The differences being attributed to the lower percentage of associative and higher percentage of other behavior in the purse-seine caught group. Analyses of the mean durations of events classified by behavior type indicated no significant differences for characteristic (ANOVA, F (1,115) = 0.69, P = 0.41), associative (ANOVA, F (1,32) = 1.50, P = 0.23), and other (ANOVA, F (1,127) = 2.22, P = 0.14) events between the two groups.
The spatial and temporal patterns in the three different behavior types, and their durations, along the movement paths of three distinctly different sizes of bigeye are illustrated in Fig. 7. It is evident in this figure that the 64–77-cm bigeye (Fig. 7a), in comparison with the 115–128-cm bigeye (Fig. 7c), shows a higher proportion of days exhibiting associative behavior (22.4% versus 9.4%) and lower percentage of days exhibiting characteristic behavior (26.9% versus 76.2%). The distributions of the three behavior types along each of the movement paths show some clustering, indicative of the non-random distribution of floating objects leading to associative behavior, and DSL prey organisms leading to characteristic behavior.

Most probable movement paths, with behavior types for each daily position estimate, for three fish of different lengths: 64–77 cm (tag no. 0390070) (a), 80–106 cm (tag no. 0390057) (b), and 115–128 cm (tag no. 1113) (c)
The proportions of time which the 96 bigeye exhibited each of the three different behavior types within 5-degree areas by three length classes are illustrated in Fig. 8. A qualitative evaluation of these data indicates there do not appear to be any spatial differences, within the three length classes, of the distributions of behavior types. However, the majority of these data are distributed within a very restricted area. The higher proportions of characteristic behavior and lower proportions of associative and other behavior, with increasing size of the length class, are, however, apparent.

Proportion of time bigeye exhibit one of three distinctive behavior types within 5-degree areas by length classes, 54–79.9 cm (a), 80–99.9 cm (b), and ≥100 cm (c). The size of the circle in each map is scaled to the legend, given as days of data
The summary statistics for the classification of daily behavior for the 96 bigeye during their times at liberty as characteristic, associative, or other, by age class, are given in Table 4. For age classes 1.25 through 3.75 years, there are positive correlations between the percentage of time fish exhibited characteristic behavior (Pearson, r 2 = 0.72, n = 11, P = 0.001) and age, and between the average duration of those events (Pearson, r 2 = 0.47, n = 11, P = 0.021) and age. There is a negative correlation in the percentage of time fish exhibited associative behavior (Pearson, r 2 = 0.83, n = 11, P < 0.0001) and age, and between the average duration of those events (Pearson, r 2 = 0.43, n = 11, P = 0.029) with age. There is also a negative correlation in the percentage of time fish exhibited other behavior (Pearson, r 2 = 0.44, n = 11, P = 0.025) and age, and between the average duration of those events (Pearson, r 2 = 0.49, n = 11, P = 0.017) with age.
Deep-diving behavior
An example of deep-diving behavior exhibited by a 77-cm fish for 1 week is shown in Fig. 9a, and for one day in Fig. 9b. During the week, the fish, which exhibiting characteristic and other behavior, undertook five deep dives, in excess of 500 m, with each occurring in the morning. The deep dive shown in Fig. 9b was to a maximum depth of 1,565 m, minimum of 3.1°C, and a duration of 88 min from the initial descent until it ascended to within the mixed layer. This dive profile includes the cooling of core temperature to about 16.9°C, and the inability for heart function at low ambient temperatures (Brill et al. 2005) was potentially the physiological limitation that caused the fish to ascend back into the warm upper mixed layer.

Depth and temperature records for an estimated 77-cm bigeye tuna (tag no. 1103), exhibiting deep-diving behavior during the week (5/31 to 6/6/2005) (a), and during a day (6/3/2005) (b)
For 92 fish, there were 1,618 deep-diving events, which occurred primarily during daylight hours (Fig. 10a), with a duration of from 3 to 392 min and an average of 54 min (Fig. 10b), and maximum depths ranged from 500 to 1,902 m with an average of 853 m (Fig. 10c). Deep-diving events are not extremely common for bigeye observed in this study, as the frequency of such events was an average of 3.5 dives per fish/month (range: 2.2 to 7.1 dives per fish/month). Deep-diving events occurred 60.1% of the time on days classified as characteristic, 5.6% of the time as associative, and 34.3% of the time as other.

Distributions of the time of day (a), duration (b), and maximum depth (c) for 1,618 deep-diving events exhibited by 92 fish
Vertical habitat
The vertical habitat utilized by the 96 bigeye while east of 110°W, when exhibiting non-associative (characteristic and other) behavior, is presented by length classes, as composite distributions, by night and day, along with thermal profiles, in Fig. 11. The average depths and percentage of time below the thermocline, for those data illustrated in Figs. 11a-c, by length classes, are given in Table 5. Analyses of these data indicate significant differences between length classes in the nighttime (ANOVA, F (2,7,128,885) = 15,714, P < 0.0001) and daytime (ANOVA, F (2,8,118,411) = 28,335, P < 0.0001) average depths. These differences are a result of the fish in the length class 100–159.2 cm showing deeper daytime average depths, and the fish in the length class 80–99.9 cm showing shallower nighttime average depths.

Vertical habitat utilization, by day (open bars) and night (solid bars), for bigeye 54–79.9 cm (a), 80–99.9 cm (b), and 100–159.2 cm (c), for all days classified as characteristic or other type behavior. The points along the lines are the average temperatures within depth intervals. n = total number of records
A comparison of vertical habitat distributions during daytime, for the same data sets for 4 bigeye recaptured by purse-seine vessels and 4 recaptured by longline vessels, described in the section above, when exhibiting non-associative (characteristic and other) behavior was conducted. The purse-seine caught fish spent 76.9% of their time below the thermocline (69 m) at an average depth of 177 m, and the longline-caught fish spent 79.0% of their time below the thermocline (69 m) at an average depth of 185 m. Analyses of those data indicate a significant difference (ANOVA, F (1,147,944) = 203.0, P < 0.0001) in the average depth distributions for the two groups of fish. The difference is the result of the longline-caught fish having a slightly deeper average daytime depth distribution.
For the 96 bigeye, when exhibiting associative behavior east of 110°W, the average nighttime and daytime depths by length classes are given in Table 5. Analyses of those data indicate significant differences in the average nighttime (ANOVA, F (2, 1,106,365) = 4,109, P < 0.0001) and daytime (ANOVA, F (2,1,236,128) = 3,513, P < 0.0001) depth distributions for the three length classes of fish. The differences are the result of the fish of 100–159.2 cm having, on average, a slightly deeper daytime depth distribution and the fish of 80–99.9 cm having, on average, a slightly shallower nighttime depth distribution, when associated with floating objects.
The average daytime depths, by 1-degree of longitude, for fish with tag no. 590051 (Table 1) between June 2007 and June 2009 (127–159 cm length), when exhibiting characteristic behavior are shown in Fig. 12. There is a significant positive correlation (Pearson, r 2 = 0.50, n = 81, P < 0.0001) between the average daytime depths by 1-degree of longitude, for days classified as characteristic behavior, and increasing westward longitudes. The average depth values were 202 m for 90° to 95°W, 220 m for 120° to 125°W, and 251 m for 150° to 155°W.

Average daytime depths by longitude, for fish with tag no. 590051 between June 2007 and June 2009 (127–159 cm), when exhibiting characteristic behavior
Discussion
These results represent a comprehensive analysis of the greatest amount of archival tag data published to date for any marine fish, which enabled us to quantitatively evaluate the vertical movements, behavior, and habitat utilization of bigeye in the equatorial EPO. The results indicate that the behavior, for all sizes and ages of fish observed in this study, is dominated by non-associative (characteristic and other) type behavior. Characteristic behavior by bigeye is not ubiquitous, although sometimes inferred, as other type behavior makes up a significant component of their behavioral repertoire. Associative behavior with floating objects is far less common than non-associative behavior, but observed for bigeye over the entire length (54–159 cm) and age (0.97–5.44 years) ranges in this study. It is a common misconception that associative behavior is exhibited only by juvenile bigeye, since their length at 50% maturity was estimated to be 135 cm in the eastern and central Pacific (Schaefer et al. 2005). The vertical habitat data in this study indicate that bigeye show some significant differences in their depth distributions, by length and age, when exhibiting non-associative and associative behavior by night and day, but the differences found are trivial.
There were problems with the longer-term performance of the archival tags utilized in this experiment, which limited the amount of useful data which could have been collected, based on the numbers of tag recoveries and times at liberty given in Table 1.
Vertical movements and behavior
The length- and age-specific estimates of the proportions of time and durations of events in which bigeye exhibited characteristic, associative and other type behaviors in this study are a significant improvement over previous estimates given by Schaefer and Fuller (2002). The current study is based on more data and expanded size and age distributions. The time-series data used for the classification of behavior types in this study was truncated, so as to exclude the bias in the initial time series in which the fish were associated with the tagging vessel, which was included in the previous study (Schaefer and Fuller 2002). In addition, in the current study, the estimates for proportions of time and durations of events in which bigeye exhibited each of the three behavior types is given by length and age classes, whereas in the previous study all sizes and ages were combined, providing only a single estimate for each behavior type. The high percentage of associative behavior reported for age class 1 fish in Table 4 is apparently an artifact of the relatively small amount of data available for that age class. It is thus uncertain whether there is a significant difference in the proportion of associative behavior by fish within age class 1 relative to the other age classes.
There was a very limited amount of useful data in this study for comparative analyses of the behavior of bigeye recaptured by purse-seine vessels fishing on floating objects, versus those recaptured by longline gear. However, from the results of the analyses conducted on those data for evaluations of differential behavior and/or habitat utilization, it appears that the differences are trivial and do not help to explain the recapture rates by the different gear types. We believe that bigeye recaptures by longline gear versus purse-seine gear is primarily a function of encounter rates by the bigeye when exhibiting daytime non-associative behavior, based on their geographical distribution relative to that of longline fishing effort. Further evidence of no differential behavior, for tagged bigeye recaptured by longline versus purse-seine gear, is provided by several bigeye recaptures during the same time period covered in this study, by purse-seine vessels fishing on floating objects in the equatorial eastern Pacific Ocean (EEPO), tagged and released at about 40–80 cm, on Korean longline fishing vessels operating in the eastern and central Pacific.
The frequency and duration in which bigeye exhibit different behavior types should be correlated with their encounters of prey and/or floating objects. Food-habit studies (King and Ikehara 1956; Blunt 1960) and fine-scale acoustic and sonar studies in the Pacific (Josse et al. 1998; Dagorn et al. 2000; Schaefer and Fuller 2005) indicate bigeye forage predominantly on DSL prey organisms, consisting primarily of cephalopods and mesopelagic fish. The diel vertical movement patterns for bigeye exhibiting characteristic behavior are indicative of active searching for prey items of the vertically migrating DSL (Longhurst 1976), as previously reported for bigeye in this area (Schaefer and Fuller 2002; Schaefer and Fuller 2005). The vertical movement patterns for bigeye when exhibiting other behavior are most likely reflective of the behavior of their prey. The short average durations of all behavior types are indicative of the transitional and highly adaptive foraging strategy of bigeye relative to the fine-scale spatiotemporal variability in the distribution and abundance of prey within their habitat.
High concentrations of forage organisms is obviously an important biotic factor in the fine-scale distribution and abundance of bigeye and appears to be one of the primary factors contributing to their observed regional fidelity to the highly productive area of the EEPO (Schaefer and Fuller 2009). The horizontal distribution of the DSL in the EEPO is not homogeneous, but patchy, and its distribution (horizontal and vertical), densities, variability, and other important characteristics have been inadequately investigated. DSL prey organisms appear to be a very important forage base for large pelagics, including tropical tunas (Schaefer and Fuller 2009) and marine mammals (Leatherwood and Evans 1979; Fiedler et al. 1998), and probably strongly influences their distributions, and yet the importance in the overall ecology of large pelagics has received little attention.
The distribution of floating objects in the EEPO is also not uniform, inferred from the distribution of catches of tunas associated with floating objects (Anonymous 2008). Drifting FADs are commonly deployed by purse-seine fishing vessels in preferred spatio-temporal areas, taking into consideration biological productivity, current patterns, and previous experience. The estimates of the durations of associative events by bigeye with floating objects are, on average, quite short about 2–3 days (Table 2), although events have been recorded to last for up to 24 days (Fig. 6b). The longevity of such events is probably related to the nighttime occurrence or absence of DSL prey organisms within mixed layer depths along the drift trajectories of those floating objects. Associative behavior by bigeye with moored weather buoys in Hawaiian waters for up to 34 days, based on their uniformly shallow distributions (<100 m), following release with implanted archival tags was reported by Musyl et al. (2003). It was also reported in that study that bigeye tagged with archival tags exhibited associative behavior with floating objects for up to 10 days, after exhibiting non-associative characteristic behavior, during their times at liberty.
Stomach content analysis of FAD-associated bigeye in the eastern Atlantic Ocean by Ménard et al. (2000) showed that 82.7% of the stomachs were empty, whereas only 25% of the stomachs of unassociated bigeye were empty. Although we agree with the conclusion in that study that FADs do not have a trophic function, it appears that an obvious bias in that study and most other tuna food-habit studies results from the dependency on collections of stomach samples from fish caught only during daylight hours. Based on our numerous field observations of the foraging of FAD-associated bigeye, skipjack, and yellowfin (Thunnus albacares) tunas on DSL prey organisms during the night in the EEPO (Schaefer and Fuller 2005), food-habit studies should be considered biased if they do not include stomach samples collected throughout the night, as well as during the day.
The most probable movement paths for the 96 fish utilized in this investigation (Fig. 1) show strong fidelity to a fairly restricted area of the EEPO (Schaefer and Fuller 2009). Even the movement path for the bigeye at liberty for 4.1 years (tag no. 590051, Table 1), which undertook two fairly extensive cyclical migrations to the central Pacific during its third and fourth years at liberty, shows strong fidelity to the area in which it was released. The movement paths (Fig. 1), along with the limited and short duration associative events with floating objects for all 96 bigeye in this study (Table 2), indicate that bigeye are not being transported extensive distances from the eastern to the western Pacific by westward-flowing currents, while associated with drifting FADs.
Although the estimates of the proportion of time which bigeye exhibit associative behavior with floating objects from the smallest to largest length classes is just 18.9–8.8% (Table 2), bigeye apparently have a greater affinity for associative behavior than do skipjack or yellowfin (Schaefer et al. 2009). Unfortunately, we know little about behavior patterns of skipjack tuna, including associative behavior with drifting FADs. However, considering the fact that skipjack are the primary catch of industrial purse-seine fisheries targeting tuna aggregations associated with floating objects throughout the Pacific, there is an urgent need to compile information on the behavior of skipjack similar to that which has been reported in this study for bigeye. The behavior of bigeye is strongly influenced by the presence of drifting FADs within their habitat. Even though associative behavior in this study was found to be limited in occurrence and duration, bigeye are highly vulnerable to capture by purse-seine vessels, as indicated by an average annual retained catch in the EPO of about 69,030 mt during 2000–2007 (Anonymous 2008).
Deep-diving behavior
Deep-diving behavior, dives in excess of 500 m, is not uncommon for bigeye as reported in this study and that of Evans et al. (2008), nor for other tuna species (Gunn and Block 2001; Schaefer et al. 2009). A plausible reason why they undertake these occasional and sometimes extremely deep dives is foraging for deep water squids (Roper and Young 1975; Roper et al. 1984) or mesopelagic fishes (Maynard et al. 1975). The profiles of these dives including the depths, ambient and internal temperatures, and dissolved oxygen (DO) concentrations are very informative regarding bigeye physiological tolerances and foraging strategies. The bigeye deep-dive profiles illustrate remarkable physiological thermoregulation (Holland and Sibert 1994; Malte et al. 2007), plus an understanding of the basis of how these fish can survive dives to depths in excess of 1,000 m, well below the oxygen minimum zone (about 300–500 m), where the DO concentrations are about 2 ml/L (Schaefer et al. 2009).
The deepest observed dive by a bigeye tuna (tag no. 2332, Table 1) in this study of 1,902 m, with total dive duration of 1.5 h, is the deepest dive recorded for any tuna species. That dive exceeds even the deepest dives recorded by tags of deep-diving large pelagics including swordfish, Xiphius gladius, at 1,185 m (Abascal et al. 2009), sperm whales, Physter macrocephalus, at 1,304 m (Aoki et al. 2007), and beaked whales, Ziphius cavirostris, at 1,888 m (Tyack et al. 2006). Deep dives in excess of 1,000 m have also been reported from archival tag data for bigeye tuna in the Coral Sea (Evans et al. 2008), and for other tunas including yellowfin in the Indian Ocean (Dagorn et al. 2006) and EPO (Schaefer et al. 2007; Schaefer et al. 2009), and Atlantic bluefin (Thunnus thynnus; Block et al. 2001).
Vertical habitat
The composite depth and temperature habitat utilization distributions for the three length classes (Fig. 11), while exhibiting non-associative (characteristic and other) behavior, indicate very little variability and the differences appear trivial. From the smallest (54–79.9 cm) to the largest (100–159.2 cm) length classes in this study, the vertical habitat utilized when exhibiting non-associative behaviors was 98.9–97.9% of the time above the thermocline depth (60 m) during the night, at the same average depth of 34 m, and 59.5–72% of their time below the thermocline during the day at an average depth of 163 to 183 m. For the same smallest to largest length classes, when exhibiting associative behavior, the average nighttime and daytime depths ranged from 25 to 21, and 33 to 37 m, respectively.
Depths of the DSL in the tropical EPO have been reported to be 300–400 m during the day and 0–100 m at night (Tont 1976; Fiedler et al. 1998). For bigeye in this study >100 cm, exhibiting non-associative behavior (Fig. 11c), the mode of maximum daytime depths is about 225–270 m at an ambient temperature about 13°C. Evans et al. (2008) report bigeye of comparable size with archival tags in the Coral Sea, exhibiting non-associative behavior, to have a deeper mode of maximum daytime depth at about 400–450 m, at similar ambient temperatures of about 12–14°C. It seems probable that the deeper daytime depths exhibited by bigeye when exhibiting characteristic behavior in the western Pacific are related to differences in the oceanography, including greater daytime DSL depths (Kuznetsov et al. 1982; Fiedler and Talley 2006). Bigeye vertical habitat is undoubtedly constrained by hypoxic conditions (Brill et al. 2005), perhaps anything less than 1 ml/L (Hanamoto 1987; Schaefer et al. 2009), the top of the oxygen minimum layer (OML; Fiedler and Talley 2006). It appears that during the daytime when the DSL depths are below those of the top of the OML in the EEPO (300 m; Fiedler and Talley 2006) or Coral Sea (500–600 m; Evans et al. 2008) bigeye are physiologically limited in their searching time at depths of their DSL prey, which have adapted to living in extremely low DO concentrations (Childress and Seibel 1998; Gilly et al. 2006). During the nighttime, however, when the DSL prey organisms ascend into the mixed layer depth (MLD) they are no longer distributed within a refugium, and bigeye foraging success escalates. Locating areas, while exhibiting characteristic behavior during the day, where DSL prey organisms are abundant, even though foraging success is most likely limited by physiological constraints, expectations to capitalize on those prey in that area at night may be an important facet of successful bigeye foraging strategy.
The archival tag data sets compiled in this study contain a wealth of information about the spatio-temporal dynamics of bigeye behavior and habitat in the EEPO. There are numerous applications, some currently realized and others not yet discovered, for which this information needs to be incorporated into stock assessments. Habitat-based stock assessment models have been developed for the integration of behavioral and environmental data, in order to standardize catch per unit of effort (CPUE) data, based on estimating fishing depths of longline gear in relation to the vertical habitat utilization of the target species, such as bigeye tuna, by time of day (Bigelow et al. 2002; Maunder et al. 2006). We suggest that information provided in this study on vertical behavior and habitat utilization, including associative behavior with floating objects, be given serious consideration for exploring vulnerability to detection and catchability by purse-seine and longline vessels so as to be integrated into stock assessment models for standardization of CPUE data.
Large-scale regional tuna tagging projects, utilizing archival tags, are needed throughout the Pacific to further improve our understanding of bigeye, as well as skipjack and yellowfin movements, behavior, and habitat utilization. Such information is useful for a better understanding of their ecology and improving stock assessments for these species. Greater knowledge of the physiological abilities and tolerances of these species derived through laboratory studies, and their habitat utilization derived through archival tags, are required to understand changes in their behavior and gear vulnerability relative to variable oceanographic conditions in order to develop appropriate procedures for standardization of CPUE. Elucidating the behavior of those species, when associated and non-associated with floating objects, could also permit the formulation of optimal purse-seine fishing techniques and/or strategies, including the reduction in catches of undesirable sizes of those tunas and also other non-target species.
References
Abascal FJ, Mejuto J, Quintans M, Ramos-Cartelle A (2009) Horizontal and vertical movements of swordfish in the southeast Pacific. ICES Jour Mar Sci 67:1–9
Aires-da-Siva A, Maunder MN (2009) Status of bigeye tuna in the eastern Pacific Ocean in 2007 and outlook for the future. Inter-Amer Trop Tuna Comm Stock Assess Rept 9:101–202
Anonymous (2008) Tunas and billfishes in the eastern Pacific Ocean in 2007. Fishery status report 6. Inter-Amer Trop Tuna Comm 140 pp
Aoki K, Amano M, Yoshioka M, Mori K, Tokuda D, Miyazaki N (2007) Diel diving behavior of sperm whales off Japan. Mar Ecol Prog Ser 349:277–287
Bigelow K, Hampton J, Miyabe N (2002) Application of a habitat-based model to estimate effective longline fishing effort and relative abundance of Pacific bigeye tuna (Thunnus obesus). Fish Oceanogr 11:143–155
Block BA (2005) Physiological ecology in the 21st century: advancements in biologging science. Integr Comp Biol 45:305–320
Block BA, Dewar H, Blackwell S, Williams T, Farwell CJ, Prince ED, Boustany A, Teo SLH, Seitz A, Fudge D, Walli A (2001) Electronic tags reveal migratory movements, depth preferences and thermal biology of Atlantic bluefin tuna. Science 293:1310–1314
Blunt CE Jr (1960) Observations on the food habits of longline caught bigeye and yellowfin tuna from the tropical eastern Pacific 1955–1956. Calif Fish Game 46:69–80
Brill R, Bigelow K, Musyl M, Fritsches K, Warrant E (2005) Bigeye tuna (Thunnus obesus) behavior and physiology and their relevance to stock assessments and fishery biology. ICCAT Col Vol Sci Pap 57:142–161
Childress JJ, Seibel BA (1998) Life at stable low oxygen levels: adaptations of animals to oceanic oxygen minimum layers. J Exp Biol 201:1223–1232
Dagorn L, Bach P, Josse E (2000) Movement patterns of large bigeye tuna (Thunnus obesus) in the open ocean, determined using ultrasonic telemetry. Mar Biol 136:361–371
Dagorn L, Holland KN, Hallier JP, Taquet M, Moreno G, Sancho G, Itano DG, Aumeeruddy R, Girard C, Million J, Fonteneau A (2006) Deep diving behavior observed in yellowfin tuna (Thunnus albacares). Aquat Living Resour 19:85–88
Evans K, Langley A, Clear NP, Williams P, Patterson T, Sibert J, Hampton J, Gunn JS (2008) Behaviour and habitat preferences of bigeye tuna (Thunnus obesus) and their influence on longline fishery catches in the western Coral Sea. Can J Fish Aquat Sci 65:2427–2443
Fiedler P, Talley L (2006) Hydrography of the eastern tropical Pacific: a review. Prog Ocean 69:143–180
Fiedler P, Barlow J, Gerrodette J (1998) Dolphin prey abundance determined from acoustic backscatter data in eastern Pacific surveys. Fish Bull 96:237–247
Gilly WF, Markaida U, Baxter CH, Block BA, Boustany A, Zeidberg L, Reisenbichler K, Robison B, Bazzino G, Salinas C (2006) Vertical and horizontal migrations by the jumbo squid Dosidicus gigas revealed by electronic tagging. Mar Ecol Prog Ser 324:1–17
Gunn JS, Block BA (2001) Advances in acoustic, archival, and satellite tagging of tunas. In: Block BA, Stevens ED (eds) Tunas: physiology, ecology, and evolution. Academic Press, San Diego, pp 167–224
Hanamoto E (1987) Effect of oceanographic environment on bigeye tuna distribution. Bull Jap Soc Fish Oceanogr 51:203–216
Holland KN, Sibert JR (1994) Physiological thermoregulation in bigeye tuna, Thunnus obesus. Environ Biol Fish 40:319–327
Holland KN, Brill RW, Chang RKC (1990) Horizontal and vertical movements of yellowfin and bigeye tuna associated with fish aggregating devices. Fish Bull 88:493–507
Josse E, Bach P, Dagorn L (1998) Simultaneous observations of tuna movements and their prey by sonic tracking and acoustic surveys. Hydrobiologia 371(372):61–69
King J, Ikehara I (1956) Comparative study of food of bigeye and yellowfin tuna in the central Pacific. US Fish Wildl Serv Fish Bull 57:61–85
Kuznetsov I, Stefanov S, Savagov V (1982) A migrating sound scattering layer in the equatorial Pacific Ocean. Oceanology 22:702–703
Lam CH, Nielsen A, Sibert J (2008) Improving light and temperature based geolocation by unscented Kalman filtering. Fish Res 91:15–25
Le Gall JY, Cayre P, Taquet M (eds) (2000) Mechanisms and effects of the aggregation of tuna by FADs. Aqua Liv Res 13:181–262
Leatherwood S, Evans WE (1979) Some recent uses and potentials of radiotelemetry in field studies of cetaceans. In: Winn HE, Olla BL (eds) Behavior of Marine Animals, current perspectives in research, vol 3. Plenum Press, New York-London, pp 1–31 Cetaceans
Longhurst AR (1976) Vertical migration. In: Cushing DH, Walsh JJ (eds) The ecology of the seas. Blackwell, London, pp 116–137
Malte H, Larsen C, Musyl M, Brill R (2007) Differential heating and cooling rates in bigeye tuna (Thunnus obesus Lowe): a model of non-steady state heat exchange. Jour Exp Biol 210:2618–2626
Matsumoto T, Okamoto H, Toyonaga M (2006) Behavioral study of small bigeye, yellowfin and skipjack tunas associated with drifting FADs using ultrasonic coded transmitter in the Central Pacific Ocean. WCPFC-SC2-2006/FT IP-7
Maunder M (2005) Report from the workshop on developing indices of abundance from purse-seine catch and effort data. Inter-Amer Trop Tuna Com Scientific meeting, November 3–5, 2004, La Jolla, CA (http://www.iattc.org/PDFFiles2/Report_PS_CPUE_meeting_Nov04ENG.pdf) (compiler)
Maunder M, Hinton M, Bigelow K, Langley A (2006) Developing indices of abundance using habitat data in a statistical framework. Bull Mar Sci 79:545–559
Maynard SD, Riggs FV, Walters JF (1975) Mesopelagic micronekton in Hawaiian waters: faunal composition, standing stock, and diel vertical migration. Fish Bull 73:726–736
Ménard F, Stequert B, Rubin A, Herrera M, Marchal E (2000) Food consumption of tuna in the equatorial Atlantic Ocean: FAD-associated versus unassociated schools. Aquat Liv Res 13:233–240
Musyl MK, Brill RW, Boggs CH, Curran DS, Kazama TK, Seki MP (2003) Vertical movements of bigeye tuna (Thunnus obesus) associated with islands, buoys, and seamounts near the main Hawaiian Islands from archival tagging data. Fish Oceanogr 12:152–169
Roper CFE, Young RE (1975) Vertical distribution of pelagic cephalopods. Smithsonian Contrib Zool 209:51
Roper CFE, Sweeney MJ, Nauen CE (1984) FAO species catalogue, vol. 3, Cephalopods of the world. An annotated and illustrated catalogue of species of interest to fisheries. FAO Fisheries Synopsis 3(125), p 277
Schaefer KM, Fuller DW (2002) Movements, behavior, and habitat selection of bigeye tuna (Thunnus obesus) in the eastern equatorial Pacific, ascertained through archival tags. Fish Bull 100:765–788
Schaefer KM, Fuller DW (2005) Behavior of bigeye (Thunnus obesus) and skipjack (Katsuwonus pelamis) tunas within aggregations associated with floating objects in the equatorial eastern Pacific. Mar Biol 146:781–792
Schaefer KM, Fuller DW (2006) Estimates of age and growth of bigeye tuna (Thunnus obesus) in the eastern Pacific Ocean, based on otolith increments and tagging data. Inter-Amer Trop Tuna Comm Bull 23:32–76
Schaefer KM, Fuller DW (2009) Horizontal movements of bigeye tuna (Thunnus obesus) in the eastern Pacific Ocean, as determined from conventional and archival tagging experiments initiated during 2000–2005. Inter-Amer Trop Tuna Comm Bull 24:191–247
Schaefer KM, Fuller DW, Miyabe N (2005) Reproductive biology of bigeye tuna (Thunnus obesus) in the eastern and central Pacific Ocean. Inter-Amer Trop Tuna Comm Bull 23:1–31
Schaefer KM, Fuller DW, Block BA (2007) Movements, behavior, and habitat utilization of yellowfin tuna (Thunnus albacares) in the northeastern Pacific Ocean, ascertained through archival tag data. Mar Biol 152:503–525
Schaefer KM, Fuller DW, Block BA (2009) Vertical movements and habitat utilization of skipjack (Katsuwonus pelamis), yellowfin (Thunnus albacares), and bigeye (Thunnus obesus) tunas in the equatorial eastern Pacific Ocean, as ascertained through archival tag data. In: Nielsen JL, Arrizabalaga H, Fragoso N, Hobday A, Lutcavage M, Sibert J (eds) Reviews: methods and technologies in fish biology and fisheries, vol 9, tagging and tracking of marine animals with electronic devices. Springer, Berlin, pp 121–144
Scott MD, Bayliff WH, Lennert-Cody CE, Schaefer KM (1999) Proceedings of the international workshop on the ecology and fisheries for tunas associated with floating objects. Inter Amer Trop Tuna Comm Spec Rep 11:480 (compilers)
Tont SA (1976) Deep scattering layers: patterns in the Pacific. Rep Calif Coop Ocean Fish Invest 18:112–117
Tyack PL, Johnson M, Soto NA, Sturlese A, Madsen PT (2006) Extreme diving of beaked whales. Jour Exp Biol 209:4238–4253
Zar JH (1974) Biostatistical analysis. Prentice Hall, New Jersey
Acknowledgments
This research was made possible through financial contributions by the Japan Fisheries Agency and the Taiwan Fisheries Agency. We are grateful for invaluable insights on the dynamics of FAD fishing in the EPO provided by Captains A. Parker and D. Stephenson. We are thankful to captain B. Blocker and his crew aboard the MV Her Grace for their relentless efforts in fishing and tagging operations. We are indebted to vessel owners, captains, fishermen, stevedores, and industry representatives for returning recovered tags. Thanks also to IATTC field office personnel for their efforts in recovering tags and recapture information. We thank Nick Vogel for his assistance with the development of computer programs for the processing of archival tag data for classifying daily behaviors. We also thank Tim Lam for his assistance in deriving a most probable movement path for the 4.1 year bigeye data set. We are grateful to B. Bayliff, R. Deriso, and two anonymous reviewers for constructive comments on drafts of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Garthe.
Rights and permissions
About this article
Cite this article
Schaefer, K.M., Fuller, D.W. Vertical movements, behavior, and habitat of bigeye tuna (Thunnus obesus) in the equatorial eastern Pacific Ocean, ascertained from archival tag data. Mar Biol 157, 2625–2642 (2010). https://doi.org/10.1007/s00227-010-1524-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-010-1524-3