
Abstract
Ecological and physiological studies focused on dietary preferences, lipid biochemistry and energetics within the three Antarctic chaetognaths Eukrohnia hamata, E. bathypelagica and E. bathyantarctica from meso- and bathypelagic depths. Eukrohnia hamata and E. bathypelagica respired 0.15 μL O2 mg dry mass (DM)−1 h−1, which translates to an average metabolic loss of only <1.1% of body carbon per day. Lipid storage was not substantial in E. bathypelagica (mean 11.5 ± 6.5% DM) and E. bathyantarctica (mean 15.4 ± 4.1% DM) during summer and winter, suggesting year-round feeding of these predators mainly on copepods. In E. bathypelagica, total fatty acids were dominated by the fatty acids 16:0, 20:5(n-3) and 22:6(n-3) and in E. bathyantarctica also by 18:1(n-9), a fatty acid usually found in storage lipids. Only the latter species was characterized by significant amounts of wax esters, consisting largely of the common fatty alcohols 16:0, 20:1(n-9) and the unusual fatty alcohol isomer 22:1(n-9).
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Chaetognatha are a small phylum that consists of about 150 species (Kapp 2004). They are known as important predators in all oceans, including the Southern Ocean (Pakhomov et al. 1999) and may contribute substantially to total zooplankton abundance and biomass (Hosie and Cochran 1994; Pakhomov et al. 1999, 2000). As important predators of copepods (Øresland 1990, 1995) and as a significant food source for a wide variety of larger organisms, they hold a central position in planktonic food webs (Feigenbaum 1991).
Intensive studies especially on the taxonomy, abundance and distribution of Antarctic chaetognaths have been carried out in the past 100 years (e.g. Ritter-Záhony 1911; David 1958; Alvariño et al. 1983a, b; Hagen 1985; Kruse et al. 2009). Other than their diet composition, there was limited information on the feeding behavior of chaetognaths due to the difficulty to keep these delicate animals alive in the laboratory (Feigenbaum and Maris 1984; Feigenbaum 1991). One of the first attempts to systematically analyze the gut content of chaetognaths was made by Wimpenny (1936) on Sagitta setosa and S. elegans. A first report of controlled laboratory feeding was given by Reeve (1964) on Sagitta hispida. Subsequent studies also primarily focused on the species-rich genus Sagitta. In contrast, our knowledge on the feeding ecology of the genus Eukrohnia is rudimentary, except for the abundant E. hamata (e.g. Øresland 1990; Froneman et al. 1998). To date, there are no studies available on the feeding of Antarctic meso- and bathypelagic chaetognaths.
Gut contents and fatty acid trophic markers were analyzed to investigate the feeding preferences of Eukrohnia hamata and the two deeper-living species E. bathypelagica and E. bathyantarctica. The trophic marker concept is based on the largely unmodified incorporation of dietary fatty acids into the body lipid of zooplankton (e.g. Lee et al. 1971b). In our study, this observation was used to estimate the chaetognath dietary composition. It is the first application to the deeper-living Eukrohnia species. As information on the metabolism and energy budget in this genus is limited (Båmstedt 1979; Thuesen and Childress 1993), we studied lipids and respiration of the three chaetognath species. Our aim was to elucidate the feeding habits and lipid dynamics of these meso- and bathypelagic predators.
Materials and methods
Sampling
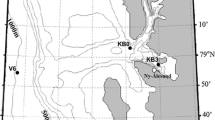
Chaetognaths were sampled with RV “Polarstern” in the Weddell Sea during the Antarctic winter 2006 (17 June–21 August 2006, ANT 23-6) and the Antarctic summer 2007/2008 (28 November 2007–04 February 2008, ANT 24-2). These two expeditions were carried out within the scope of the German krill project LAKRIS (Meyer 2005). The study area was located between 60°–70°S and 3°W–3°E (Fig. 1). During winter, samples were taken at 28 stations (with eight repetitive stations at ~66°S 0°E) with a multinet (MN: five nets, 100 μm mesh size, 0.25 m2 mouth area) and at three stations with a rectangular midwater trawl (RMT 8: 4.5 mm mesh size, 8 m2 mouth area; RMT 1: 320 μm mesh size, 1 m2 mouth area). In summer, the MN was deployed at 15 stations (two at 52°S 0°E, not shown on Fig. 1) and a multiple RMT, consisting of three pairs of nets, at four stations (two analyzed in this study). The MN sampled the following standard depth intervals during both seasons: 2,000–1,500–1,000–750–500–0 m (except for two stations to 3,000 m, one to 1,250 m and one to 1,500 m; see Kruse et al. 2009). For this study, only chaetognaths from below 500 m depth were analyzed. The winter RMT hauls included the depth range from the surface to approximately 3,000 m and back to the surface, whereas the multiple RMT was deployed to an opening depth at 1,900 and 2,500 m, respectively (at 64°30′S 3°E: 1,900–1,500–750–500 m; at 63°S 0°E: 2,500–2,000–1,000–500 m; Fig. 1). The RMT 8 cod-end bucket carried a volume of approximately 26 l of water, which ensured that the animals were suspended. This method yielded high survival rates and healthy specimens.

Map of stations along three transects during the expeditions ANT 23-6 (winter) and ANT 24-2 (summer) in the Weddell Sea, Southern Ocean. MN multinet, RMT rectangular midwater trawl, black winter stations, white summer stations
Prior to the experiments and analyses, all chaetognaths were identified using the relevant literature (Alvariño 1969; Casanova 1999). They were assigned to maturity stages and measured (head to tail, excluding tail fin) to the nearest 0.5 mm under a stereomicroscope (Olympus SZX12) either on board or later in the home laboratory. Eukrohnia bathypelagica and E. bathyantarctica destined for lipid analysis were classified into maturity stages according to Alvariño (1967, 1969). Specimens carrying brood sacs (only empty brood sacs in the investigated specimens) were assigned to an additional fifth stage.
Chaetognaths of the species E. hamata, E. bathypelagica and E. bathyantarctica were proven to be in good condition in the MN samples. They were either quickly rinsed with Milli-Q water and stored in glass vials at −80°C for biochemical analysis or maintained alive in respiration experiments. A few specimens of E. bathypelagica and E. bathyantarctica were also taken from the RMT for the lipid and fatty acid analyses. The remaining MN and RMT samples were either frozen or preserved in formaldehyde (4% final concentration, buffered with hexamine) for later investigations.
Gut content analysis
Gut contents of formaldehyde-preserved and frozen chaetognath specimens from MN and RMT samples were analyzed. Two hundred specimens of each species (Eukrohnia hamata, E. bathypelagica, E. bathyantarctica) were investigated from both summer and winter season. Each chaetognath was transferred to a few drops of glycerine on a microscope slide. The presence of lipid droplets in the gut was recorded. The gut fullness was estimated prior to dissection as percentage of total gut length filled with content and divided into four categories (<10, 10–25, 25–50, 50–100%). When ripe ovaries filled the body cavity of E. bathypelagica, gut fullness was determined after dissection of the specimen. Jellyfish remains found in the guts were not included for the determination of gut fullness. The gut was dissected with fine needles (0.15 mm) under a stereomicroscope and further analyzed under an inverted microscope (Axiovert 40C; up to 400× magnification). The percentage of Eukrohnia specimens containing food (food containing ratio = FCR) and the number of prey per chaetognath (NPC) were recorded for each species. The FCR is given as total FCR when the gut contained food, even if it was an unidentifiable mass. The FCR for identifiable prey was additionally calculated. Prey items found in the foregut were omitted from the analysis to avoid bias from possible cod-end feeding.
Analyses of dry mass, elemental and biochemical composition
For dry mass (DM), carbon (C) and nitrogen (N) analyses the chaetognaths of the three species Eukrohnia hamata (summer: n = 72, winter: n = 179), E. bathypelagica (summer: n = 40, winter: n = 43) and E. bathyantarctica (summer: n = 42, winter: n = 35) from MN and RMT samples were freeze-dried for 24 h and weighed on a Sartorius microbalance (Sartorius Micro and Sartorius Supermicro 4504 MP8). Each specimen was ground and then analyzed (entire animal or subsample) on a Euro Elemental Analyzer for C and N compositions with acetanilide as standard.
The lipid and fatty acid analyses focused on the two meso- and bathypelagic species E. bathypelagica and E. bathyantarctica. Eukrohnia bathypelagica individuals originated from 500 to 2,000 m depth, whereas E. bathyantarctica individuals were mostly taken from 1,500 to 2,000 m depth. To ensure sufficient biomass for lipid analysis, some individuals were pooled to obtain more than 1 mg DM per sample. Total lipid was then extracted from single lyophilized individuals or pooled samples with dichloromethane/methanol (2:1 by volume), and the lipid content was determined gravimetrically according to Folch et al. (1957) as modified by Hagen (2000).
Aliquots of the extracted samples were taken for characterization and quantification of the fatty acid and alcohol fractions by gas–liquid chromatographic analysis. Lipids were hydrolyzed, and the fatty acids were converted to their methyl ester derivatives in methanol containing 3% concentrated sulfuric acid at 80°C for 4 h (Kattner and Fricke 1986). The resulting fatty acid methyl esters and free fatty alcohols were separated and quantified using a Hewlett-Packard gas chromatograph (HP 6890A), equipped with a DB-FFAP column (30 m length, 0.25 mm inner diameter). Peaks were identified by comparing retention times with those from fish oil (Marinol) and copepod lipid lab standards of known composition, verified with commercially available standard mixtures (e.g. Matreya). If necessary, additional confirmation was carried out by gas chromatography–mass spectrometry. The proportions of wax esters relative to total lipid were estimated for E. bathyantarctica on the basis of the fatty alcohol to fatty acid contents.
The trophic ratio [18:1(n-9) + 20:1(n-9) + 22:1(n-9)]/[16:1(n-7) + 18:1(n-7) + 18:4(n-3)] was calculated as an indicator of carnivory, where the first three fatty acids are markers for carnivory and calanoid copepod ingestion, respectively, and the three latter fatty acids for phytoplankton ingestion (diatoms and flagellates, respectively; e.g. Falk-Petersen et al. 1990; Dalsgaard et al. 2003). Fatty acids that occurred with a portion of ≥1% of total fatty acids in at least one case during both seasons are presented in the results. The remaining minor fatty acids were combined to a single group.
Respiration rate measurements
Specimens from the 500 to 1,000 m depth interval (multinet samples only) were chosen for the respiration experiments with Eukrohnia hamata and E. bathypelagica to ensure good physiological conditions of the chaetognaths and to minimize decompression damage. Additionally, individuals from the 1,000 to 1,500 m depth range were included in eight analyses on E. hamata in winter and in four analyses on E. bathypelagica in summer. Eukrohnia bathyantarctica was not included in the respiration experiments because it was difficult to collect enough specimens with similar properties in good condition.
Before the start of the experiment, the chaetognaths were acclimated for up to a day at constant temperature (0°C) in the dark. For respiration rate measurements, specimens were incubated in sealed glass bottles of about 60 or 500 mL volume filled with filtered (0.7 μm pore size) and oxygen-saturated seawater (saturation above approximately 92%) for 19–26 h at the acclimation temperature and light conditions. During the winter expedition, two specimens were incubated in a 60-mL bottle. In summer, additional experiments were conducted with four specimens per 500-mL bottle to evaluate the influence of bottle volume on respiration. Individuals similar in size and maturity (primarily stage II) were chosen from the same depth range for each measurement. Each respiration experiment consisted of three to eight bottles with chaetognaths and two to four controls without chaetognaths. During winter and summer, 52 and 51 measurements (including 34 in 500-mL bottles) were done with E. hamata individuals. Five and nine measurements (including eight in 500-mL bottles) were performed with E. bathypelagica individuals in winter and summer. Measurements of dissolved oxygen were performed by the Winkler titration method (Grasshoff 1983) on the entire sample bottle or on subsamples, which were carefully siphoned out of the 500-mL bottles. The decrease in oxygen concentration in the bottles with chaetognaths compared to the controls was always less than 10%. All data were normalized per unit dry mass. Specimens that remained in the 500-mL experimental bottles and specimens separately frozen from the net samples were taken for dry mass and carbon content determination.
To estimate the metabolic loss in terms of the absolute and relative amount of carbon, respired by E. hamata and E. bathypelagica per day, the measured oxygen consumption rates in μL O2 individual−1 h−1 were converted to carbon units using a respiratory quotient (RQ) value of 0.97 in the following equation (Ikeda et al. 2000):
where 12/22.4 is the carbon weight (12 g) in 1 mol of CO2 (22.4 L). For E. bathyantarctica, we applied a comparable calculation based on the general DM and carbon measurements using the average respiration rate of the two other Eukrohnia species.
Statistical analyses
Differences in carbon (C; μg mg DM−1), nitrogen (N; μg mg DM−1) and lipid content (mg lipid mg DM−1) were analyzed by a full factorial two-way ANOVA for each parameter (C/N/lipid versus species & season & species × season) with a subsequent post hoc Tukey HSD test on differences between means (α = 0.05, Sokal and Rohlf 1981). We applied a full factorial two-way ANOVA for each species (lipid content versus maturity stage & season & maturity stage × season) to test for stage-specific differences in the lipid content of Eukrohnia bathypelagica and E. bathyantarctica. This analysis was done separately because the same maturity stages were not present for both species during both seasons. Stage I and III specimens were excluded from the analyses for E. bathypelagica, and stages IV and V were excluded for E. bathyantarctica analyses. Individuals of these stages were not in sufficiently good condition during winter.
A full factorial three-way ANOVA was used to examine the influence of depth on respiration (μL O2 mg DM−1 h−1; respiration versus species & season & depth & species × season & species × depth & season × depth & species × season × depth). The 1,000–1,500 m depth range was excluded from the analysis, since no measurements were conducted with E. hamata from this depth range in summer and with E. bathypelagica in winter. A t-test was applied to analyze the dependence of bottle volume on respiration of E. hamata in summer. Prior to these parametric tests, the data were Box-Cox transformed (Sokal and Rohlf 1981) when necessary to achieve normality and homogeneity of variances.
We performed a principal component analysis (PCA) on the fatty acid compositions (percentages) to analyze differences between species, maturity stages, depths and seasons. Fourteen common fatty acids found in E. bathypelagica and E. bathyantarctica were included in this analysis. All statistical analyses were performed with the software Statistica 8 (StatSoft).
Results
Gut content
Less than half of all Eukrohnia hamata specimens analyzed had food in their guts (39% during summer and 42% during winter; total FCR, Table 1). In E. bathypelagica, 48% contained food during winter and 31% in summer. In both species, the degree of gut fullness was rather low, usually less than 25%. Gut contents were mostly located in the middle and posterior part of the gut. In contrast to the former two species, the gut of E. bathyantarctica is characterized by an orange color. Due to this pigmentation, the gut content was more difficult to identify in this species and the gut fullness could not be recorded precisely. Even after careful dissection, no identifiable particles were detected in E. bathyantarctica.
In E. hamata and E. bathypelagica we found identifiable prey items, although most prey were in advanced stages of decomposition. Copepods, protozoans (Acantharia, Radiolaria, tintinnids), jellyfish remains (particularly nematocysts) and diatoms were the main sources of food identified in both species (Table 1). Only one complete copepod and few mandibles (particularly coxal gnathobases) of copepods were found in comparatively good condition. These copepod remains could not be related to the respective species. Excluding diatoms, Acantharia and jellyfish remains, on average ≤0.11 prey items were observed per specimen (NPC, Table 1).
Regardless of the season, oil droplets were found in the guts of most Eukrohnia hamata specimens (>82%). In E. bathypelagica, 34% of all guts contained oil droplets in summer and 57% in winter (Table 1). Due to the intense gut pigmentation and the oil globules in the intestinal tissue, oil droplets in the guts of E. bathyantarctica were difficult to identify even after dissection.
Carbon, nitrogen and total lipid content
The carbon and nitrogen contents of the meso- and bathypelagic chaetognaths depended significantly on season (ANOVA, for C: F = 117.206, P < 0.001; for N: F = 72.595, P < 0.001) and species (ANOVA, for C: F = 40.442, P < 0.001; for N: F = 23.825, P < 0.001). Carbon, nitrogen and C:N ratios were higher in winter than in summer (Table 2). In Eukrohnia bathypelagica, we observed the lowest carbon (winter: 31.3% DM, summer: 24.6% DM) and nitrogen contents (winter: 6.9% DM, summer: 5.7% DM) of all three species. The C and N contents of E. hamata and E. bathyantarctica were similar but generally higher than those of E. bathypelagica. The same was true for the C:N ratio of E. bathypelagica (winter: 4.5, summer: 4.3), which was significantly lower than for the two other species (Tukey HSD, P < 0.001).
The total lipid contents of the two deeper-living species E. bathypelagica and E. bathyantarctica were significantly different from each other (ANOVA, F = 9.026, P < 0.01; Tables 3, 4). Eukrohnia bathyantarctica had an average lipid content of 15.4% DM (SD ± 4.1) and showed no significant variability related to maturity stage (ANOVA, F = 1.393, P > 0.05) or season (ANOVA, F = 0.001, P > 0.05; Table 4). In contrast, the lipid contents of E. bathypelagica ranged between 1.4 and 25.4% DM and differed distinctly among maturity stages (ANOVA, F = 5.499, P < 0.05; Table 3). Stage IV (winter: 12.7% DM, summer: 20.0% DM) contained relatively more lipids than stages II and V. Moreover, a higher lipid level in E. bathypelagica was measured in summer than in winter (winter: 8.4% DM, summer: 13.9% DM; ANOVA, P < 0.05; all maturity stages included).
Fatty acid and fatty alcohol compositions of E. bathypelagica and E. bathyantarctica
Total lipids of Eukrohnia bathypelagica consisted mainly of fatty acids (mean 95.9% of total fatty acids and alcohols) with only minor amounts of fatty alcohols (mean 4.1% of total fatty acids and alcohols). Eukrohnia bathyantarctica had higher amounts of fatty alcohols (Table 4; mean 22.8%) compared to E. bathypelagica. The fatty alcohols in E. bathyantarctica indicated the presence of wax esters with a contribution of up to 57.3% of total lipids.
The five fatty acids 16:0, 16:1(n-7), 18:1(n-9), 20:5(n-3) and 22:6(n-3) generally dominated within the two chaetognath species E. bathypelagica and E. bathyantarctica and comprised 60.3 and 69.0% of total fatty acids (TFA), respectively. However, there were distinct differences between the fatty acid patterns of these two species, which are reflected by the clustering in the principal component analysis (PCA, Fig. 2). The PCA based on the fatty acid composition extracted three principal components with eigenvalues >1% of variance. Results were presented for the major two components, together accounting for 73.9% of variance (Fig. 2).

Principal component analysis (PCA) of Eukrohnia bathypelagica and E. bathyantarctica according to their fatty acid compositions. Two-dimensional PCA score (a) and loading (b) plot of the first two principal components (PC1 and PC2) based on the fatty acid profiles of both species (14 common and relevant fatty acids). Eukrohnia bathypelagica—dots (empty dots stage IV individuals in the 1,500–2,000 m depth range), E. bathyantarctica—triangles
The fatty acid composition of E. bathypelagica was characterized by higher amounts of 16:0, 20:5(n-3) and 22:6(n-3). These fatty acids contributed mean values of 9.7, 11.0 and 22.1%, respectively, to TFA (Table 3). Season, depth and maturity stage had a marginal influence on the fatty acid compositions of E. bathypelagica. The PCA revealed a grouping of stage IV E. bathypelagica individuals originating from 1,500 to 2,000 m depth that contained lower mean amounts of 22:6(n-3) (winter: 15.1%, summer: 13.3%) compared to the other maturity stages (Fig. 2). Hence, particularly stages I, II and V from 500 to 1500 m depth were similar in their fatty acid composition with the exception of three samples (Fig. 2). Generally, moderate amounts of the monounsaturated fatty acids 16:1(n-7) (mean 7.5%) and 18:1(n-9) (mean 10.0%) were measured in E. bathypelagica (Table 3). The fatty acids 20:1(n-9) and 22:1(n-11) represented up to 14.3% (mean 4.9%) and 20.7% TFA (mean 4.3%) in this species. The remaining fatty acids each accounted ≤4% TFA on average.
Eukrohnia bathyantarctica contained high levels of 18:1(n-9), with a maximum of 39.1% of TFA (Table 4; mean 32.7%). The fatty acids 16:0, 20:5(n-3) and 22:6(n-3) were considerably less abundant in E. bathyantarctica compared to E. bathypelagica. On average 22:6(n-3) accounted for 11.5% TFA (Table 4), which is about half of its contribution to TFA in E. bathypelagica. The fatty acids 16:0 and 20:5(n-3) contributed only 6.5 and 5.9% to TFA, respectively. 16:1(n-7) and 18:1(n-7) exhibited higher percentages of 12.3 and 6.4% in this species, whereas similarly low levels of 20:1(n-9) were measured. PCA revealed a species cluster with no apparent seasonal, depth or maturity stage-related pattern identifiable (Fig. 2).
The fatty acid ratio [18:1(n-9) + 20:1(n-9) + 22:1(n-9)]/[16:1(n-7) + 18:1(n-7) + 18:4(n-3)], used as an indicator of carnivory, was similar in E. bathypelagica and E. bathyantarctica with mean values of 1.9 and 2.0, respectively (Tables 3, 4). The fatty alcohol content was highly variable in E. bathypelagica and ranged between 0 and 26.6% of TFA and total fatty alcohols (TFAlc; mean 4.1%, not shown in the table). Eukrohnia bathyantarctica contained a higher portion of fatty alcohols (mean 22.8% of TFA and TFAlc) compared to E. bathypelagica. These fatty alcohols mainly consisted of 16:0 (mean 25.2% TFAlc), 18:1(n-9) (13.3%), 20:1(n-9) (17.7%) and 22:1(n-9) (18.5%; Table 4).
Oxygen consumption
The respiration experiments showed that the bottle volume (1 ind. per approximately 140 or 30 mL) had no significant influence on oxygen consumption of Eukrohnia hamata (t-test, t = 1.34, P > 0.05; 0.16 ± 0.09 μL O2 mg DM−1 h−1 for 140 mL and 0.14 ± 0.09 μL O2 mg DM−1 h−1 for 30 mL). Mean respiration rates of 0.15 μL O2 mg DM−1 h−1 (SD ± 0.08) were measured in E. hamata and E. bathypelagica (Fig. 3). No significant differences were observed between species (ANOVA, F = 0.941, P > 0.05) and seasons (ANOVA, F = 0.637, P > 0.05). Mass-specific oxygen consumption did not differ between specimens originating from different depths (ANOVA, F = 2.201, P > 0.05; 500–750, 750–1,000 m).

Eukrohnia hamata and E. bathypelagica. Oxygen consumption during winter and summer at 0°C. n Number of measurements
The mean metabolic loss for all three Eukrohnia species during both seasons was very low, with <1.1% of body carbon respired per day (Table 2). The metabolic loss tended to be lower in winter (<0.5%) due to the higher body carbon contents but constant respiration rates.
Discussion
Feeding ecology
Our results on chaetognath feeding suggest a copepod-based diet, although protozoans and chaetognaths may also be found in the chaetognaths’ guts. The fatty acid analyses of Eukrohnia bathypelagica and E. bathyantarctica revealed species-specific results, which further indicate differences in the diet between these two species. The feeding ecology of the three investigated chaetognath species will be discussed in detail in the following section.
Copepods create a distinct flow field in the water (Bundy and Paffenhöfer 1996; Jiang and Osborn 2004), which is readily detected by chaetognaths via sensory hairs (Newbury 1972; Feigenbaum and Reeve 1977). Based on this fact and due to the dominance of copepods, chaetognaths feed largely on copepods (e.g. Øresland 1990; Froneman and Pakhomov 1998). The Antarctic E. hamata preys on the copepodite stages of Calanus spp., Euchaeta spp., Metridia gerlachei and Rhincalanus gigas as well as on smaller copepods like Microcalanus pygmaeus, Oithona spp. and Oncaea spp. (Hopkins 1985, 1987; Hopkins and Torres 1989; Øresland 1990, 1995; Froneman et al. 1998). Apart from diatoms and jellyfish remains, copepods were the most frequent food items found in the guts of E. hamata and E. bathypelagica in our study. Due to the high degree of digestion, mainly parts of copepods were found. Even the few mandible remains and the one complete copepod in the chaetognaths’ guts could not be identified to species level. They could belong to deep-sea copepods because their characteristics do not seem to match species frequently encountered in the upper hundred meters of the Southern Ocean. Diatoms were assumed to be ingested inadvertently or via herbivorous prey (Feigenbaum 1991). Medusae were also assumed to be artifacts of collecting and preserving methods rather than being true prey items (Feigenbaum 1991 and references therein). These were found particularly in the guts of chaetognaths from RMT samples, which often also contained jellyfish. Some degree of cod-end feeding or better indiscriminately grasping within the cod-end cannot be excluded. Appendicularians, ostracods and other chaetognaths were documented in the chaetognath’s diet (Øresland 1990; Froneman et al. 1998; Froneman and Pakhomov 1998). Few chaetognath hooks or tintinnids were observed in E. hamata guts, although the latter prey is more common in young chaetognaths (Pearre 1981). The two acantharians found in the guts of E. hamata were in very good condition and probably accidentally ingested by the chaetognaths, either in the cod-end or eaten via their copepod prey. To our knowledge, no data exist on gut contents of the two deeper-living species E. bathypelagica and E. bathyantarctica. Due to several methodological constraints of the gut content analysis, which will be discussed below, these analyses provide only rough estimates of the feeding preferences.
Generally, the frequency of chaetognaths with gut content is low (e.g. David 1955; Hopkins 1985; Lancraft et al. 1991; Øresland 1995). In our investigation, the number of chaetognaths containing food in their guts ranged between 31 and 48% (FCR total) for E. hamata and E. bathypelagica, irrespective of the state of digestion. In several previous studies, only the identifiable contents were counted, or as in the case of Froneman and Pakhomov (1998), oil droplets were also considered as gut content, which we treated separately. In the Australian sector of the Southern Ocean, Johnson and Terazaki (2004) found 15.4–48.9% (station means) of E. hamata specimens with prey in their guts. Usually, more than 84% of the guts of E. hamata or Sagitta gazellae contain no food items (Øresland 1995; Froneman et al. 1998). Up to 4.0% of our chaetognaths’ guts contained identifiable material when we exclude diatoms, Acantharia and jellyfish remains from our data. However, the amount of unidentifiable food was high in our study. In summer, 0.02–0.06 (Froneman et al. 1998) and 0.21 prey items (0.10–0.26 during 24 h sampling, Øresland 1990) were found in E. hamata. For the winter season, 0.08 and 0.12 prey items per E. hamata were counted at two different stations in Gerlache Strait (Øresland 1995), which is in the same range as our data (0.01–0.11, diatoms, Acantharia and jellyfish remains excluded). The mean number of prey per chaetognath (NPC) seems to be slightly higher in Sagitta compared to Eukrohnia species (Øresland 1990: S. marri: 0.23, S. gazellae: 0.26, S. maxima: 0.20).
The gut content analysis has various drawbacks that are associated with the sampling procedure. In our investigation, chaetognaths were taken from great depths, which greatly increases the retrieval time. Two uncertainties, progressed digestion and cod-end feeding, may be associated with such long retrieval times. A digestion time of 9.3 h can be assumed for large copepods, 4.9 h for small copepods and unidentified prey (for Sagitta elegans at 6°C in Gullmarsfjorden, Sweden; Øresland 1987). In the Southern Ocean, Giesecke et al. (2010) estimated a digestion time between 9 and 15.8 h (mean 11.5 h, at 0 ± 1°C) for S. gazellae, also tending to increase with larger prey. Thus, lower temperatures in the Southern Ocean may result in longer digestion times, but yet this process has to be taken into consideration. Cod-end feeding in plankton net hauls is another problem when applying gut content analysis (Baier and Purcell 1997). Therefore, prey items found in the foreguts were excluded from our analyses. We can also not exclude regurgitation in chaetognaths. Regurgitation and defecation might occur as a stress reaction on capturing or on preservation. Baier and Purcell (1997) presumed that the prey loss in chaetognath guts during sampling was due to stress-induced gut evacuation rather than to continued digestion. This could explain the generally low NPC. Prey loss seems rather to occur at the beginning of the catch, whereas cod-end feeding may take place throughout the tow (Baier and Purcell 1997). We tried to reduce the stress during the catch by using large cod-ends, thus avoiding crowded samples. Nevertheless, the sampling method seems to have a strong effect on results of chaetognath feeding, and hence it is difficult to draw conclusions on seasonal and species-specific differences in diet composition and feeding activity only on the basis of gut content analyses.
To elucidate diet preferences in the two deeper-living species E. bathypelagica and E. bathyantarctica, we therefore conducted additional fatty acid trophic marker analyses. The latter species was of special interest, as no identifiable prey items were found in its guts. The fatty acid 18:1(n-9) distinguishes these two chaetognaths. In E. bathyantarctica, it was a dominant component (32.7%), whereas E. bathypelagica contained only moderate amounts of this fatty acid (10.0%). As a trophic marker for carnivory (Falk-Petersen et al. 1990), 18:1(n-9) indicates a high level of carnivorous feeding in E. bathyantarctica. Omnivorous and carnivorous zooplankton species are known to have wax esters with high amounts of the 18:1(n-9) fatty acid (Lee et al. 2006). However, the ratio [18:1(n-9) + 20:1(n-9) + 22:1(n-9)]/[16:1(n-7) + 18:1(n-7) + 18:4(n-3)] was similar in both species. The higher this value is, i.e. the larger the amount of the three biomarkers for carnivorous feeding are relative to the three phytoplankton biomarkers, the more pronounced the degree of carnivory will be. The difference in fatty acid compositions between E. bathypelagica and E. bathyantarctica seems to reflect different prey including copepods. The high amounts of the short-chain fatty alcohols 14:0 and 16:0 again support the high degree of carnivory (Lee et al. 2006) in E. bathyantarctica. These two fatty alcohols are also major components in the fatty alcohol compositions of copepods like Rhincalanus gigas (Kattner et al. 1994) and Paraeuchaeta antarctica (Hagen et al. 1995). The fatty acids 20:1(n-9) and 22:1(n-11) and the respective alcohols are biomarkers for calanid copepods such as Calanoides acutus (Graeve et al. 1994; Kattner and Hagen 1995). The mean amounts of these long-chain monounsaturated fatty acids were low in both chaetognath species, but the relative portion of the fatty alcohol 20:1(n-9) was high in E. bathyantarctica. This suggests that both chaetognath species feed on calanid copepods, but to a different extent. However, Calanus propinquus, which produces large amounts of the unusual 22:1(n-9) fatty acid (Hagen et al. 1993), does not seem to be a preferred prey, as indicated by the low level of this fatty acid in both species. The fatty alcohol 22:1(n-9), however, made up a considerable percentage of the TFAlc composition in E. bathyantarctica. This is surprising, because this fatty alcohol is found only in trace amounts in Antarctic copepods and may have been synthesized de novo by E. bathyantarctica. We cannot prove, however, whether E. bathyantarctica is able to synthesize wax esters. The moderate mean percentages of 16:1(n-7) in E. bathypelagica and E. bathyantarctica, 7.5 and 12.3%, respectively, could suggest the ingestion of diatoms (Dalsgaard et al. 2003). However, this marker fatty acid was probably ingested via herbivorous copepods, which fed on diatoms in epipelagic layers, or via carnivorous copepods. Paraeuchaeta antarctica could be a potential prey species. This carnivorous copepod is known to accumulate considerable amounts of the monounsaturated fatty acid 16:1(n-7) (Hagen et al. 1995).
The two long-chained polyunsaturated fatty acids 20:5(n-3) and 22:6(n-3) that represented a significant percentage of the TFA in E. bathypelagica were most likely obtained from their prey. A biosynthesis of these fatty acids seems to be improbable. The higher relative portion of these fatty acids compared to E. bathyantarctica may be attributed to the lower lipid content and thus less storage lipids of E. bathypelagica.
Energetics
The C:N ratios of the three chaetognath species were above 4 on average, which is comparable to the ratios and C and N values previously reported for chaetognaths (Omori 1969: 4.2; Schneider 1990: ~4.4). Terazaki (1993) found a C:N ratio of 4.7 and 3.5 for Sagitta elegans in the Japan Sea and in the Pacific, respectively. He assumed that this high ratio in the Japan Sea is caused by body lipids, as oil droplets were observed in the intestinal tissue as in meso- and bathypelagic species (like in Eukrohnia bathyantarctica in our study). It was observed that E. hamata and E. bathypelagica had oil droplets in their guts, independent of maturity stage and season. These oil droplets were already reported by several authors for E. hamata (Sameoto 1987; Øresland 1990; Froneman et al. 1998; Froneman and Pakhomov 1998). Their function is still unknown, although they are assumed to act as energy deposit (Kapp 1991), as buoyancy aid or both (Øresland 1990). The role of these lipid droplets with regard to seasonal and breeding migrations and reproduction is not understood.
The dominance of the fatty acids 16:0, 20:5(n-3) and 22:6(n-3) in E. bathypelagica, which are typical components of biomembranes, indicates a limited dependence on lipid reserves (Graeve et al. 1997). However, lipids seem to gain in importance with increasing sexual maturity of this species. The higher lipid values in summer primarily reflect the higher number of mature specimens investigated during this season, as ripe individuals with large ovaries filling the body cavity (maturity stage IV) had maximum lipid contents. The stage V individuals carried only empty brood sacs, from which their offspring had been released. Lower lipid levels in these specimens are therefore not surprising. The ovaries in this species appear to accumulate fatty tissue (Alvariño 1983), as most brooding species do. Lipid droplets are often observed in zooplankton ovaries, which may partially be transferred to developing oocytes (Lee et al. 2006). In chaetognaths, this is probably an important strategy to support the development of the offspring from oocytes to young chaetognaths that leave their parental brood sacs. The increase in chaetognath density associated with gonad maturation may be a reason for the movement of adult individuals to deeper layers (Alvariño 1964). This is in agreement with findings that mature individuals usually occur deeper in the water column (e.g. David 1955; Hagen 1985; Kruse 2009). A higher lipid content in adult specimens would partially compensate the increased body density and also act as buoyancy aid reducing the sinking speed of adults to deeper waters.
Eukrohnia bathyantarctica showed no stage-specific differences in its lipid content. However, the lipid level was usually higher (mean 15.4% DM) than in E. bathypelagica (mean 11.5% DM). The lack of differences between maturity stages may partially be explained by their less voluminous ovaries and their smaller number of ova, as compared to E. bathypelagica (Kruse 2009). In addition, the orange gut of E. bathyantarctica probably contains a carotenoid pigment with many small oil droplets similar to the gut of E. fowleri (Terazaki 1991). This might be responsible for the relatively higher lipid content in all maturity stages of E. bathyantarctica. It remains unsolved, whether the carotenoids are derived from the prey, e.g. lipid-rich copepods, or whether the chaetognaths synthesize these pigments themselves. Based on their investigations on Eukrohnia fowleri and Sagitta macrocephala, Terazaki et al. (1977) assume that the carotenoids of the intestinal tissue are produced by the chaetognaths because these carotenoids have a unique character differing from that of phytoplankton and copepods.
Wax esters were a major lipid component in E. bathyantarctica (45.5% TL). These are efficient long-term energy reserves and also serve as buoyancy aid due to their very low density (Hagen 2000). A high wax ester content of 34 and 71% TL was found in unidentified deep-living subtropical chaetognaths with an orange colored gut (at 31°N 119°W, probably of the genus Eukrohnia, Lee et al. 1971a). In contrast, a deep-water chaetognath with orange pigment from the central South Pacific contained distinctly lower wax ester content (26% TL, Lee and Hirota 1973). The intestinal oil globules observed in E. bathyantarctica might contain wax esters that are missing in species like E. bathypelagica. We conclude that the truly bathypelagic species E. bathyantarctica apparently contains more wax esters, relies more on long-term energy reserves, and/or utilizes wax esters as buoyancy aids. These characteristics are in contrast to species that also inhabit upper mesopelagic layers like E. bathypelagica.
Eukrohnia hamata and E. bathypelagica respired 0.15 μL O2 mg DM−1 h−1 on average. They were usually inactive during the experiments. As chaetognaths have periods of activity and of inactivity in their natural environment (Thuesen and Childress 1993), our data may be a good estimate of inactive periods, i.e. basic metabolism. Seasonal differences are not expected due to an assumed year-round activity and feeding at meso- and bathypelagic depths. Båmstedt (1979) measured respiration rates between 0.10 and 0.21 μL O2 mg DM−1 h−1 for E. hamata in Kosterfjorden, western Sweden (“standard animals” of 4 mg DM, at 5–6°C, 0–200 m depth) with highest rates in spring (May) and lowest rates in winter (February). However, we found no seasonal differences. Båmstedt (1979) additionally reported a mean respiration rate of 0.11 and 0.86 μL O2 h−1 per individual E. bathypelagica and E. hamata, respectively. Our experiments at 0°C revealed consumption rates of 0.36 and 0.44 μL O2 ind.−1 h−1. The difference between the two chaetognath species was more pronounced in Sweden. However, respiration rates were still in the same range. Oxygen consumption rates between approximately 0.29 and 0.48 μL O2 ind.−1 h−1 (converted from a mean value of 0.805 μmol O2 g−1 h−1 and a wet mass range of 0.0161–0.0268 g) measured for E. hamata at 5°C (Thuesen and Childress 1993) support our data. Differences between the data sets may be attributable to the smaller number of measurements by Thuesen and Childress (1993) compared to our study. Additionally, differences in temperature and chaetognath body mass could contribute to the different results (Kruse et al. submitted). Previous investigations showed that hydrostatic pressure seems to have little effect on the metabolic activity of chaetognaths (Childress and Thuesen 1993). These authors observed no significant variation in metabolic rates at 0.101 MPa (surface) and at 10.1 MPa (~1,000 m). During our study, oxygen consumption rates were measured at normal surface pressure on chaetognaths mostly originating from 500 to 1,000 m depth, with no significant differences between the sampled strata.
As our respiration rates may be underestimations, considering higher animal activities in situ, the daily body carbon respired is probably underestimated as well. Our calculated data suggest between 0.45 and 1.09% of body carbon respired per day, which is low compared to other Antarctic taxa. Other Antarctic zooplankton species, e.g. amphipods, copepods and euphausiids, usually have higher daily metabolic losses of carbon, varying from 0.44 to 2.75% (at −0.8 to −1.4°C, Ikeda and Mitchell 1982). However, our mean values for the three Eukrohnia species are similar to data presented for the Antarctic species Sagitta gazellae with an average metabolic loss of 0.5% (at −0.9°C, Ikeda and Kirkwood 1989). Hence, Antarctic chaetognaths may differ from other zooplankton species in this respect.
Conclusions
The chaetognaths Eukrohnia hamata and E. bathypelagica from meso- and bathypelagic depths had generally low oxygen consumption rates and hence low body carbon respiration, which may indicate low food requirements in turn. Eukrohnia bathypelagica and E. bathyantarctica exhibit different modes of feeding and lipid storage. Eukrohnia bathypelagica was characterized by a lower but stage-dependent lipid level, whereas E. bathyantarctica contained a higher relative amount of total lipids and significant amounts of wax esters. The lipid reserves were not substantial with little seasonal differences during summer and winter in both species indicating the presence of year-round feeding. Based on our analyses of gut contents and trophic marker fatty acids, we assume that all three Eukrohnia species, including E. hamata, rely largely on copepods as prey (e.g. calanid copepods and Paraeuchaeta antarctica). Consequently, chaetognaths may exert a continuous predation impact on the copepod community. Feeding conditions for chaetognaths might be better in winter, when several copepod species overwinter in deeper layers (Schnack-Schiel and Hagen 1994; Hagen 1999; Auel and Hagen 2005). Similar fatty acid compositions during summer and winter point to a homogenous diet composition in meso- and bathypelagic waters, although the zooplankton community is known to vary due to seasonal vertical migration of e.g. copepods. The role of lipids in chaetognaths, also in terms of oil droplets, requires further clarification.
References
Alvariño A (1964) Bathymetric distribution of chaetognaths. Pac Sci 18:64–82
Alvariño A (1967) The Chaetognatha of the NAGA expedition (1959–1961) in the South China Sea and the Gulf of Thailand. Part 1. Systematics. NAGA Rep 4:1–197
Alvariño A (1969) Los Quetognatos del Atlántico. Distribución y notas esenciales de sistemática. Trab Inst Esp Oceanogr 37:1–290
Alvariño A (1983) Chaetognatha. In: Adiyodi KG, Adiyodi RG (eds) Reproductive biology of invertebrates. Volume I: oogenesis, oviposition and oosorption. Wiley, Chichester, pp 585–610
Alvariño A, Hosmer SC, Ford RF (1983a) Antarctic Chaetognatha: United States Antarctic Research Program Eltanin Cruises 8-28, Part 1. Biology of the Antarctic Seas XI. Antarct Res Ser 34:129–338
Alvariño A, Verfaillie DF, Ford RF (1983b) Antarctic Chaetognatha: United States Antarctic Research Program Eltanin Cruises 10-23, 25 and 27, Part 2. Biology of the Antarctic Seas XIV. Antarct Res Ser 39:69–204
Auel H, Hagen W (2005) Body mass and lipid dynamics of Arctic and Antarctic deep-sea copepods (Calanoidea, Paraeuchaeta): ontogenetic and seasonal trends. Deep Sea Res I 52:1272–1283
Baier CT, Purcell JE (1997) Effects of sampling and preservation on apparent feeding by chaetognaths. Mar Ecol Prog Ser 146:37–42
Båmstedt U (1979) Seasonal variation in the respiratory rate and ETS activity of deep-water zooplankton from the Swedish West coast. In: Naylor E, Hartnoll RG (eds) Cyclic phenomena in marine plants and animals. Proceedings 13th EMBS, Pergamon Press, Oxford, pp 267–274
Bundy MH, Paffenhöfer G-A (1996) Analysis of flow fields associated with freely swimming calanoid copepods. Mar Ecol Prog Ser 133:99–113
Casanova J-P (1999) Chaetognatha. In: Boltovskoy D (ed) South Atlantic zooplankton. Backhuys Publishers, Leiden, pp 1353–1374
Childress JJ, Thuesen EV (1993) Effects of hydrostatic pressure on metabolic rates of six species of deep-sea gelatinous zooplankton. Limnol Oceanogr 38:665–670
Dalsgaard J, St John M, Kattner G, Müller-Navarra D, Hagen W (2003) Fatty acid trophic markers in the pelagic marine environment: a review. Adv Mar Biol 46:225–340
David PM (1955) The distribution of Sagitta gazellae Ritter-Záhony. Discov Rep 27:235–278
David PM (1958) The distribution of the Chaetognatha of the Southern Ocean. Discov Rep 29:199–228
Falk-Petersen S, Hopkins CCE, Sargent JR (1990) Trophic relationships in the pelagic, Arctic food web. In: Barnes M, Gibson RN (eds) Trophic relationships in the marine environment. Proceedings of the 24th European Marine Symposium, Aberdeen University Press, Aberdeen, pp 315–333
Feigenbaum D (1991) Food and feeding behaviour. In: Bone Q, Kapp H, Pierrot-Bults AC (eds) The biology of chaetognaths. Oxford University Press, Oxford, pp 45–54
Feigenbaum DL, Maris RC (1984) Feeding in the Chaetognatha. Oceanogr Mar Biol Ann Rev 22:343–392
Feigenbaum D, Reeve MR (1977) Prey detection in the Chaetognatha: response to a vibrating probe and experimental determination of attack distance in large aquaria. Limnol Oceanogr 22:1052–1058
Folch J, Lees M, Sloane-Stanley GH (1957) A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem 226:497–509
Froneman PW, Pakhomov EA (1998) Trophic importance of the chaetognaths Eukrohnia hamata and Sagitta gazellae in the pelagic system of the Prince Edward Islands (Southern Ocean). Polar Biol 19:242–249
Froneman PW, Pakhomov EA, Perissinotto R, Meaton V (1998) Feeding and predation impact of two chaetognath species, Eukrohnia hamata and Sagitta gazellae, in the vicinity of Marion Island (Southern Ocean). Mar Biol 131:95–101
Giesecke R, González HE, Bathmann U (2010) The role of the chaetognath Sagitta gazellae in the vertical carbon flux of the Southern Ocean. Polar Biol 33:293–304
Graeve M, Hagen W, Kattner G (1994) Herbivorous or omnivorous? On the significance of lipid compositions as trophic markers in Antarctic copepods. Deep Sea Res I 41:915–924
Graeve M, Kattner G, Piepenburg D (1997) Lipids in Arctic benthos: does the fatty acid and alcohol composition reflect feeding and trophic interactions? Polar Biol 18:53–61
Grasshoff K (1983) Determination of oxygen. In: Grasshoff K, Ehrhardt M, Kremling K (eds) Methods of seawater analysis. Second, revised and extended edition. Verlag Chemie, Weinheim, pp 61–72
Hagen W (1985) On distribution and population structure of Antarctic Chaetognatha. Meeresforsch 30:280–291
Hagen W (1999) Reproductive strategies and energetic adaptations of polar zooplankton. Invert Rep Dev 36:25–34
Hagen W (2000) Lipids. In: Harris RP, Wiebe PH, Lenz J, Skjoldal HR, Huntley M (eds) ICES zooplankton methodological manual. Academic Press, San Diego, pp 113–119
Hagen W, Kattner G, Graeve M (1993) Calanoides acutus and Calanus propinquus, Antarctic copepods with different lipid storage modes via wax esters or triacylglycerols. Mar Ecol Prog Ser 97:135–142
Hagen W, Kattner G, Graeve M (1995) On the lipid biochemistry of polar copepods: compositional differences in the Antarctic calanoids Euchaeta antarctica and Euchirella rostromagna. Mar Biol 123:451–457
Hopkins TL (1985) Food web of an Antarctic midwater ecosystem. Mar Biol 89:197–212
Hopkins TL (1987) Midwater food web in McMurdo Sound, Ross Sea, Antarctica. Mar Biol 96:93–106
Hopkins TL, Torres JJ (1989) Midwater food web in the vicinity of a marginal ice zone in the western Weddell Sea. Deep Sea Res 36:543–560
Hosie GW, Cochran TG (1994) Mesoscale distribution patterns of macrozooplankton communities in Prydz Bay, Antarctica—January to February 1991. Mar Ecol Prog Ser 106:21–39
Ikeda T, Kirkwood R (1989) Metabolism and elemental composition of a giant chaetognath Sagitta gazellae from the Southern Ocean. Mar Biol 100:261–267
Ikeda T, Mitchell AW (1982) Oxygen uptake, ammonia excretion and phosphate excretion by krill and other Antarctic zooplankton in relation to their body size and chemical composition. Mar Biol 71:283–298
Ikeda T, Torres JJ, Hernández-León S, Geiger SP (2000) Metabolism. In: Harris RP, Wiebe PH, Lenz J, Skjoldal HR, Huntley M (eds) ICES zooplankton methodological manual. Academic Press, San Diego, pp 455–532
Jiang H, Osborn TR (2004) Hydrodynamics of copepods: a review. Surv Geophys 25:339–370
Johnson TB, Terazaki M (2004) Chaetognath ecology in relation to hydrographic conditions in the Australian sector of the Antarctic Ocean. Polar Biosci 17:1–15
Kapp H (1991) Some aspects of buoyancy adaptations of chaetognaths. Helgoländer Meeresunters 45:263–267
Kapp H (2004) Chaetognatha oder Pfeilwürmer—Leben und Entwicklung im Pelagial. Natur und Museum—Bericht der Senckenbergischen Naturforschenden Gesellschaft 134:398–406
Kattner G, Fricke HSG (1986) Simple gas-liquid chromatographic method for the simultaneous determination of fatty acids and alcohols in wax esters of marine organisms. J Chromatogr 361:263–268
Kattner G, Hagen W (1995) Polar herbivorous copepods—different pathways in lipid biosynthesis. ICES J Mar Sci 52:329–335
Kattner G, Graeve M, Hagen W (1994) Ontogenetic and seasonal changes in lipid and fatty acid/alcohol compositions of the dominant Antarctic copepods Calanus propinquus, Calanoides acutus and Rhincalanus gigas. Mar Biol 118:637–644
Kruse S (2009) Population structure and reproduction of Eukrohnia bathyantarctica and Eukrohnia bathypelagica in the Lazarev Sea, Southern Ocean. Polar Biol 32:1377–1387. doi:10.1007/s00300-009-0633-2
Kruse S, Bathmann U, Brey T (2009) Meso- and bathypelagic distribution and abundance of chaetognaths in the Atlantic sector of the Southern Ocean. Polar Biol 32:1359–1376. doi:10.1007/s00300-009-0632-3
Kruse S, Brey T, Bathmann U (submitted) The role of midwater chaetognaths in Southern Ocean pelagic energy flow. Mar Ecol Prog Ser
Lancraft TM, Hopkins TL, Torres JJ, Donnelly J (1991) Oceanic micronektonic/macrozooplanktonic community structure and feeding in ice covered Antarctic waters during the winter (AMERIEZ 1988). Polar Biol 11:157–167
Lee RF, Hirota J (1973) Wax esters in tropical zooplankton and nekton and the geographical distribution of wax esters in marine copepods. Limnol Oceanogr 18:227–239
Lee RF, Hirota J, Barnett AM (1971a) Distribution and importance of wax esters in marine copepods and other zooplankton. Deep Sea Res 18:1147–1165
Lee RF, Nevenzel JC, Paffenhöfer G-A (1971b) Importance of wax esters and other lipids in the marine food chain: phytoplankton and copepods. Mar Biol 9:99–108
Lee RF, Hagen W, Kattner G (2006) Lipid storage in marine zooplankton. Mar Ecol Prog Ser 307:273–306
Meyer B (2005) German contribution to SO-GLOBEC: Lazarev Sea Krill Study (LAKRIS). GLOBEC Int Newsl 11(1):46–47
Newbury TK (1972) Vibration perception by chaetognaths. Nature 236:459–460
Omori M (1969) Weight and chemical composition of some important oceanic zooplankton in the North Pacific Ocean. Mar Biol 3:4–10
Øresland V (1987) Feeding of the chaetognaths Sagitta elegans and S. setosa at different seasons in Gullmarsfjorden, Sweden. Mar Ecol Prog Ser 39:69–79
Øresland V (1990) Feeding and predation impact of the chaetognath Eukrohnia hamata in Gerlache Strait, Antarctic Peninsula. Mar Ecol Prog Ser 63:201–209
Øresland V (1995) Winter population structure and feeding of the chaetognath Eukrohnia hamata and the copepod Euchaeta antarctica in Gerlache Strait, Antarctic Peninsula. Mar Ecol Prog Ser 119:77–86
Pakhomov EA, Perissinotto R, Froneman PW (1999) Predation impact of carnivorous macrozooplankton and micronekton in the Atlantic sector of the Southern Ocean. J Mar Syst 19:47–64
Pakhomov EA, Perissinotto R, McQuaid CD, Froneman PW (2000) Zooplankton structure and grazing in the Atlantic sector of the Southern Ocean in late austral summer 1993. Part 1. Ecological zonation. Deep Sea Res I 47:1663–1686
Pearre S Jr (1981) Feeding by Chaetognatha: energy balance and importance of various components of the diet of Sagitta elegans. Mar Ecol Prog Ser 5:45–54
Reeve MR (1964) Feeding of zooplankton, with special reference to some experiments with Sagitta. Nature 201:211–213
Sameoto DD (1987) Vertical distribution and ecological significance of chaetognaths in the Arctic environment of Baffin Bay. Polar Biol 7:317–328
Schnack-Schiel SB, Hagen W (1994) Life cycle strategies and seasonal variations in distribution and population structure of four dominant calanoid copepod species in the eastern Weddell Sea, Antarctica. J Plankton Res 16:1543–1566
Schneider G (1990) Phosphorus content of marine zooplankton dry material and some consequences; a short review. Plankton Newsl 12:41–44
Sokal RR, Rohlf FJ (1981) Biometry. The principles and practice of statistics in biological research, 2nd edn. WH Freeman and company, San Francisco
Terazaki M (1991) Deep sea chaetognaths. In: Bone Q, Kapp H, Pierrot-Bults AC (eds) The biology of chaetognaths. Oxford University Press, Oxford, pp 117–121
Terazaki M (1993) Deep-sea adaptation of the epipelagic chaetognath Sagitta elegans in the Japan Sea. Mar Ecol Prog Ser 98:79–88
Terazaki M, Marumo R, Fujita Y (1977) Pigments of meso- and bathypelagic chaetognaths. Mar Biol 41:119–125
Thuesen EV, Childress JJ (1993) Enzymatic activities and metabolic rates of pelagic chaetognaths: lack of depth-related declines. Limnol Oceanogr 38:935–948
von Ritter-Záhony R (1911) Revision der Chätognathen. Dt Südpol-Exped 13:1–71
Wimpenny RS (1936) The distribution, breeding and feeding of some important plankton organisms of the south-west North Sea in 1934. I. Calanus finmarchicus (Gunn), Sagitta setosa (J. Müller), and Sagitta elegans (Verrill). Fish Invest Lond Ser 2 15:1–53
Acknowledgments
We are grateful to the captains and the crew of RV “Polarstern” for their professional support during ANT 23-6 and ANT 24-2. We also thank Petra Wencke for analyzing the lipids and fatty acids during this study and Martin Graeve for fatty acid confirmation by GC–MS. Helpful and constructive comments on the manuscript by Gerhard Kattner, Martin Graeve and three anonymous reviewers are highly appreciated. This publication is part of the doctoral thesis of Svenja Kruse.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. A. Poulet.
Rights and permissions
About this article
Cite this article
Kruse, S., Hagen, W. & Bathmann, U. Feeding ecology and energetics of the Antarctic chaetognaths Eukrohnia hamata, E. bathypelagica and E. bathyantarctica . Mar Biol 157, 2289–2302 (2010). https://doi.org/10.1007/s00227-010-1496-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-010-1496-3