
Abstract
Accurate fecundity estimates are central to population modelling of reproductive status and egg production in a fishery. Southern rock lobster (Jasus edwardsii) fisheries are managed with the consideration of performance measures based on the level of current egg production relative to the unfished stock. Egg production is a function of the size at the onset of maturity, the size structure of the stock, and the fecundity at length of lobsters. Fecundity at length of the Tasmania stock of J. edwardsii has not been reported previously and was estimated from seven sites around Tasmania. Data were analysed by classical linear regression and also generalised linear models (GLM) with gamma and Poisson distributions. GLM with a gamma distribution produced the best fit and did not require a correction factor to generate a model on the linear scale. Individual fecundity ranged from 43,918 to 660,156 and could be predicted from carapace length according to the equation F = −1.707 + 2.969 (log length). The coefficient of length approximated three, implying the fecundity at length relationship is cuboidal. Despite the spatial heterogeneity in reproductive parameters in this fishery, the inclusion of spatial information and egg development stage into the analysis only explained an additional 1% of the variance in fecundity. Thus, it appears valid to apply the relationship across the fishery without spatial separation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Southern rock lobster, Jasus edwardsii, forms one of the major single species fisheries in Australia with the revenue of approximately $220AUD million and a market capitalisation of quota units of over $740AUD million in 2008/2009. The Tasmanian stock examined in this paper come from one of the larger management zones for the species with a total allowable commercial catch (TACC) of 1,470 t year−1. A length-based population model is employed for stock assessments (Punt and Kennedy 1997) and the level of egg production relative to the unfished state is one of the performance measures used for management. Performance of the stock against this measure is spatially heterogeneous. In some areas, egg production is close to virgin levels while in other it is highly depleted (Ford 2001). There is currently no spatial resolution in management rules, although a move to a spatially explicit management regime is currently under consideration, motivated by the need to better manage reproductive output. Management of the reproductive output of this important temperate reef species is thus reliant, in part, on fecundity at size information.
Examples of the relationship between egg production and length are a fundamental need for fisheries management and are consequently prevalent in the fisheries literature. The relationship is usually modelled by classical linear regression (CLR), although the relationship between fecundity and length is often nonlinear (e.g. Annala and Bycroft 1987; MacDiarmid 1989), and the assumption of homogeneity of variance is usually violated as variance tends to increase with the increasing length. The usual solution has been to apply log transformations to both the response (fecundity) and the predictor (length) so that the transformed data satisfy the requirements of a CLR. However, to be useful to stock assessments, a predictive relationship between fecundity and length is required in the original scale of measurement, which requires a back-transformation of estimates obtained from the linear regression model. A major problem with the use of transformed data and linear models can be interpretative difficulties with the reference to the original scale of measurement. Generalised linear models (GLM) provide a means whereby the model may be adjusted to suit the data rather than vice versa. Hence, estimates are provided on the original scale of measurement that in turn removes interpretative difficulties associated with back transformations.
The aim of this study is to describe the predictive relationship between fecundity and size for the Tasmanian rock lobster fishery, and to assess whether this relationship varied according to the location, depth or developmental stage of eggs. A secondary objective was to examine the most appropriate model for describing this relationship.
Materials and methods
Sampling
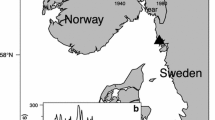
Samples of egg-bearing females were taken from the pots of commercial fishing vessels at seven sites around Tasmania during 1988 (Fig. 1). Additional samples were obtained during 1990 in order to increase the size range at two of these locations (Table 1). The timing of extrusion varies around Tasmania, occurring from late May to July in the northern regions (sites a, b, Fig. 1) and from April to May for females in southern regions (sites f, g, Fig. 1). In order to minimise the possible variation caused by egg loss during incubation, an attempt was made to collect samples at a similar period after egg extrusion in each area. With the exception of site f, this objective was achieved (Table 1).

Map of Tasmania, Australia, with an inset of Australia. a–g Mark the seven sites where sampling for egg-bearing females took place. Numbered areas around the state (1–8) are the eight zoned management units. Lines of latitude and longitude are marked
Samples were taken from the entire size range of berried females caught in each area, with the size measured as carapace length (CL, mm). Sampled lobsters were held live in the wells of vessels, so it was not possible to associate the depth of capture with the individual lobsters. Instead, individuals were classified into one of two colour classes (red or pale) because these colours have a close association with depth, red lobsters occurring predominantly above 40 m and pale lobsters most common in depths below 50 m (Chandrapavan et al. 2009).
Once landed, individuals were packed into separate plastic bags to minimise egg loss and then transported to the laboratory where the eggs from each individual were classified to one of five developmental stages (Table 2). Lobsters were then frozen. After 1 month, the lobsters were thawed, and the eggs were separated from ovigerous setae by soaking in 1 N KOH (potassium hydroxide) for 6–12 h (Bycroft 1986). Eggs were soaked until the funiculae, which attach the eggs to the setae were dissolved. Eggs were rinsed in seawater after they had separated from the setae, and the pleopods and setae were removed. The eggs were then preserved in 5% formalin with seawater and stored in the dark to prevent bleaching. Within a year of preservation, the total number of eggs on each individual was counted using an electronic egg counter (coulter counter, Bycroft 1986). Eggs from a total of 180 lobsters were counted, after excluding samples based on the missing information or because the egg separation techniques caused partial disintegration of eggs giving rise to unreliable counts (19 cases).
Analyses
Three broad types of analyses were used to examine the relationship between fecundity and size. In the first two analyses, GLMs were used with a log-link function and Poisson and gamma distributions. The third model was a CLR model, with normal distribution and assumed constant variance, in which fecundity was log transformed. All analyses used log-transformed values of carapace length. GLMs were used to assess the relationship between fecundity and size, and to check whether this relationship varied with site, egg developmental stage or depth (as inferred from colour category) using GLIM3.77 software (Royal Statistical Society, UK). In each case, distributions of the residuals were checked for normality and overdispersion of the variance by estimating the variance of the residuals and the mean fecundity in each of five length classes.
Associated with each distributional form, there is a link function which is both consistent with the nature of the response variable and has broad applicability (McPherson 1990). For a Poisson distribution, this is the log link. We plotted fecundity versus length using different scales of measurement to check the most appropriate link function, which was the log link. The reciprocal link function is the “standard” link for the Gamma distribution, although there was a linear relationship between log fecundity and log length (Fig. 2a) whereas plots of 1/fecundity versus length, log length, and the reciprocals of these were not linear. Furthermore, the use of a log link maintains a degree of compatibility with other studies involving linear models using a log transformation of fecundity and other models used in this paper.

Relationship between fecundity (number of eggs) and carapace length of southern rock lobster, Jasus edwardsii. a ln transformed fecundity and length. b. Black dashed line is the Gamma model and grey solid line is the Normal model. The curve shown for the normal model has been adjusted with a correction factor
In a linear model, the distribution of the responses (y’s) for each value of x must follow a normal distribution. This restriction is greatly reduced for GLMs, in which the responses may have a distributional form which comes from any of the exponential family of distributions (including normal, binomial, multinomial, Poisson, and gamma), but the error term (e) of a GLM is in the original scale of measurement.
Results
GLM with Poisson distribution
There was a linear relationship when log fecundity was plotted versus log length (Fig. 2a), suggesting the log-link function was reasonable provided that analyses were conducted using log-transformed values of length. The variance was much larger than the mean and the variance appears to increase exponentially with increases in the mean. Consequently, the Poisson distribution was unacceptable for predicting the fecundity at length relationship, so no further Poisson models were examined.
GLM with gamma distribution
The normality plot of the residuals was close to linear indicating that the assumption of a gamma distribution was reasonable. Carapace length explained most (90.94%) of the variation in fecundity. There was no evidence of a depth effect (as inferred by colour) on the relationship between fecundity and length (Table 3). Fecundity differed between sites (P < 0.01) and after this effect was removed, differences could be detected between the egg development stages (P < 0.05, Table 3). The inclusion of site and egg development stage into the analysis explained only an additional 1% of the variance in fecundity. There was no evidence that site or egg development stage interacted to influence fecundity estimates (Table 3). This suggests that the “best” model for the relationship between fecundity and length is one with a common slope, but different intercepts for both sites and egg development stages. There was not a clear relationship between fecundity and egg development stage. Fecundity counts were lowest for stage 1 eggs, although there was not a continuous trend in the size of counts from stage 1 through to stage 5.
Log-linear CLR
On the transformed scale, the relationship between fecundity and length was linear with constant variance (Fig. 2a). When the residuals were examined, there was no distinct trend in the variance–mean relationship and the absolute differences in variances were small (never exceeding a magnitude of two), indicating that the assumption of constant variance on the transformed scale was reasonable. The best model was one with a common slope for the length and with different intercepts for both site and egg development stage (Table 4; Fig. 2b). Parameter estimates were very similar to those from the gamma distribution with log link (Table 3; Fig. 2b), as were results comparing fecundity between sites and egg stages (Table 5). There was no evidence of a depth effect (as inferred by colour) on the relationship between fecundity and length (Table 5). Fecundity differed between sites and egg development stages (P < 0.05, Table 5). There was no evidence that site or egg development stage interacted to influence fecundity estimates (Table 5). Following the formula provided by Sprugel (1983), the correction factor for the best model was 1.0003622, and the correction factor for the length only model was 1.0004584.
Discussion
Fecundity at length relationship
Length was a very good predictor of fecundity in the Tasmanian southern rock lobster stock, explaining 91% of the variation in the data. The relationship between length and fecundity followed a cuboidal trajectory (i.e. coefficient approximating 3), following basic body-size scaling laws where volume (which restricts the number of eggs which can be retained) increases as a cube of length. A similar relationship was described in the same species in two States in Australia (VIC and SA) and New Zealand (MacDiarmid 1989; Hobday and Ryan 1998; Linnane et al. 2008), and in confamilials Panulirus regius (Freitas et al. 2007), P. marginatus (DeMartini et al. 2003), P. argus (Bertelsen and Matthews 2001), P. pencillatus (Junio 1987) and Palinurus elephas (Galhardo et al. 2006). In contrast, a linear relationship between length and fecundity has been reported in some other Palinurids, e.g. Palinurus elephas (Goni et al. 2003), P. gilchristi (Groeneveld 2005) Panulirus longipes cygnus (Morgan 1972). The shape of the fecundity–length relationship is generally influenced by sampling the full size range or reproductive females.
There were significant differences in the elevation of the fecundity at length relationship between sites, although these were small in magnitude and no pattern was apparent between the location of sites and the parameter estimates for those sites. Similar unexplained site differences occurred in the same species in New Zealand (Annala and Bycroft 1987), although there were differences in slopes between sites that were not apparent in the present study. The two most northerly sites in Tasmania had estimates that were more similar to the two most southerly sites than to the more central sites. This spatial pattern contrasts with other population characteristics such as size at first maturity, which has a clear trend with latitude (Gardner et al. 2006). Depth, as inferred here by lobster shell colour, did not influence the fecundity at length relationship.
Our strategy to account for variation caused by egg loss during incubation did not include a temporal sampling as did the study by Annala and Bycroft (1987). Instead, we graded egg samples by the developmental stage and incorporated this into the overall analysis. After site effects were removed, there was a significant effect of egg development stage on the fecundity at length relationship, although egg number increased, rather than decreased as would be consistent with a loss through incubation. We can, therefore, reject the null hypothesis that egg loss increased during incubation. One of a number of possible explanations is that females that went into berry earlier in the season produced more eggs. Individual females would need to be tracked throughout the egg-bearing season to accurately assess the rate and direction of a change on egg number.
Choosing the best fit
The model including site effects added little in the way of accuracy to the model predicting fecundity from length alone. This implies that fecundity at length parameters can be estimated from pooled data, thus reducing the risk of extrapolation beyond the size range of lobsters from smaller sample sizes for each site.
Of the three broad types of analysis conducted, only GLM with gamma distribution and CLR with log normal distribution provided a reasonable fit to the data. Data in the form of counts (such as number of eggs) often follow a Poisson distribution in which, the variance equals the mean. The model with a Poisson distribution was not satisfactory because of a non-linear variance/mean relationship. Furthermore, a severe case of overdispersion (variance greater than the mean) was evident for the Poisson model. The Poisson distribution is a distribution of independent events, hence, there is an assumption that the generation of one event (e.g. the production of an egg) does not change the probability of another event (i.e. the production of another egg), although, production of individual eggs is unlikely to be an independent process. Oocytes develop simultaneously in the ovary, and after extrusion, eggs are attached to the abdomen as clusters of eggs, not independent units. Any loss of eggs is likely to involve a loss of numerous eggs rather than a single egg. The present study considered a number of explanatory variables including length, site, egg stage, and depth (inferred from colour), although, there may have been other important variables that were not considered. The unexplained results for site and egg stage estimates suggest that other factors may be involved.
Both gamma and log normal distributions provided a reasonable fit to the data. Both models resulted in the same conclusions, produced similar parameter estimates with similar standard errors and provided a good description of the relationship between fecundity and length. The GLM with a gamma distribution and log-link function has the advantage over the CLR model approach with log-transformed fecundity of working directly with fecundity in the original scale of measurement; thus, there was no need for a correction factor for bias.
Conclusion
Fecundity of J. edwardsii varied with site and egg development stage, but based on the present data, not enough to warrant their inclusion into population models used for fisheries management purposes. Consequently, a model predicting fecundity from length alone, according to F = −1.707 + 2.969 (log length), is the most cost-effective solution to assessing fecundity of rock lobster on a large scale. A gamma distribution was the best estimator of the relationship between fecundity and length of the southern rock lobster, J. edwardsii.
References
Annala JH, Bycroft BL (1987) Fecundity of the New Zealand red rock lobster, Jasus edwardsii. N Z J Mar Freshw Res 21:591–597
Bertelsen RD, Matthews TR (2001) Fecundity dynamics of female spiny lobster (Panulirus argus) in a south Florida fishery and Dry Tortugas National Park lobster sanctuary. Mar Freshw Res 52:1559–1565. doi:https://doi.org/10.1071/MF01214
Bycroft BL (1986) A technique for separating and counting rock lobster eggs. N Z J Mar Freshw Res 20:623–626
Chandrapavan A, Gardner C, Linnane A, Hobday D (2009) Colour variation in the southern rock lobster Jasus edwardsii and its economic impact on the commercial Industry. N Z J Mar Freshwater Res 43:537–545
DeMartini EE, DiNardo GT, Williams HA (2003) Temporal changes in population density, fecundity, and egg size of the Hawaiian spiny lobster (Panulirus marginatus) at Necker Bank, Northwestern Hawaiian Islands. Fish Bull (Wash DC) 101:22–31
Ford W (2001) Restructuring the Tasmanian rock-lobster fishery—the effect of two years of management under individual transferable quotas. Mar Freshw Res 52:1641–1648. doi:https://doi.org/10.1071/MF01109
Freitas R, Medina A, Correia S, Castro M (2007) Reproductive biology of spiny lobster Panulirus regius from the northwestern Cape Verde Islands. Afr J Mar Sci 29:201–208. doi:https://doi.org/10.2989/AJMS.2007.29.2.5.188
Galhardo AC, Serafim P, Castro M (2006) Aspects of the biology and fishery of the European spiny lobster (Palinurus elephas) from the southwest coast of Portugal. J Crustacean Biol 26(4):601–609. doi:https://doi.org/10.1651/S-2715.1
Gardner C, Frusher S, Barrett NS, Haddon M, Buxton C (2006) Spatial variation in size at onset of maturity of female southern rock lobster Jasus edwardsii around Tasmania, Australia. Sci Mar 70:423–430. doi:https://doi.org/10.3989/scimar.2006.70n3423
Goni R, Quetglas A, Renones O (2003) Size at maturity, fecundity and reproductive potential of a protected population of the spiny lobster Palinurus elephas (Fabricius, 1787) from the western Mediterranean. Mar Biol (Berl) 143:583–592. doi:https://doi.org/10.1007/s00227-003-1097-5
Groeneveld JC (2005) Fecundity of spiny lobster Palinurus gilchristi (Decapoda: Palinuridae) off South Africa. Afr J Mar Sci 27:231–238
Hobday DK, Ryan TJ (1998) Contrasting sizes at sexual maturity of southern rock lobsters (Jasus edwardsii) in the two Victorian fishing zones: implications for total egg production and management. Mar Freshw Res 48:1009–1014. doi:https://doi.org/10.1071/MF97143
Junio MAR (1987) Some aspects of the reproduction of Panulirus penicillatus. Bull Mar Sci 41:242–252
Linnane A, Penny S, Ward T (2008) Contrasting fecundity, size at maturity and reproductive potential of southern rock lobster Jasus edwardsii in two South Australian fishing regions. J Mar Biol Assoc UK 88:583–589. doi:https://doi.org/10.1017/S0025315408001021
MacDiarmid AB (1989) Size at onset of maturity and size-dependent reproductive output of female and male spiny lobsters Jasus edwardsii (Hutton)(Decapoda, palinuridae) in Northern New Zealand. J Exp Mar Biol Ecol 127:229–243. doi:https://doi.org/10.1016/0022-0981(89)90076-2
McPherson G (1990) Statistics in scientific investigation: its basis, application and interpretation. Springer, New York
Morgan GR (1972) Fecundity in Western rock lobster Panulirus longipes cygnus (George) (Crustacea–Decapoda–Palinuridae). Aust J Mar Freshw Res 23:133–141. doi:https://doi.org/10.1071/MF9720133
Punt AE, Kennedy RB (1997) Population modelling of Tasmanian rock lobster, Jasus edwardsii, resources. Mar Freshw Res 48:967–980. doi:https://doi.org/10.1071/MF97070
Silberbauer BI (1970) The biology of the South African rock lobster Jasus lalandii (H. Milne Edwards) 1. Development. Division of Sea Fisheries, 92, Cape Town
Sprugel DG (1983) Correcting for bias in Log-transformed allometric equations. Ecology 64:209–210. doi:https://doi.org/10.2307/1937343
Acknowledgments
We thank D. Tarbath, R. Pearn, P. Terry, G. Zucherans, M. Edmunds and C. Sanderson for field and laboratory assistance. Thanks also to G. McPherson for invaluable advice for data analyses.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by F. Bulleri.
Rights and permissions
About this article
Cite this article
Green, B.S., Gardner, C. & Kennedy, R.B. Generalised linear modelling of fecundity at length in southern rock lobsters, Jasus edwardsii . Mar Biol 156, 1941–1947 (2009). https://doi.org/10.1007/s00227-009-1226-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-009-1226-x